MicroRNAs as Biomarkers in Amyotrophic Lateral Sclerosis



Abstract
:1. Introduction
1.1. Epidemiology and Genetic Factors
1.2. Diagnosis and Treatment
2. Role of Biomarkers in ALS
3. miRNAs as Biomarkers
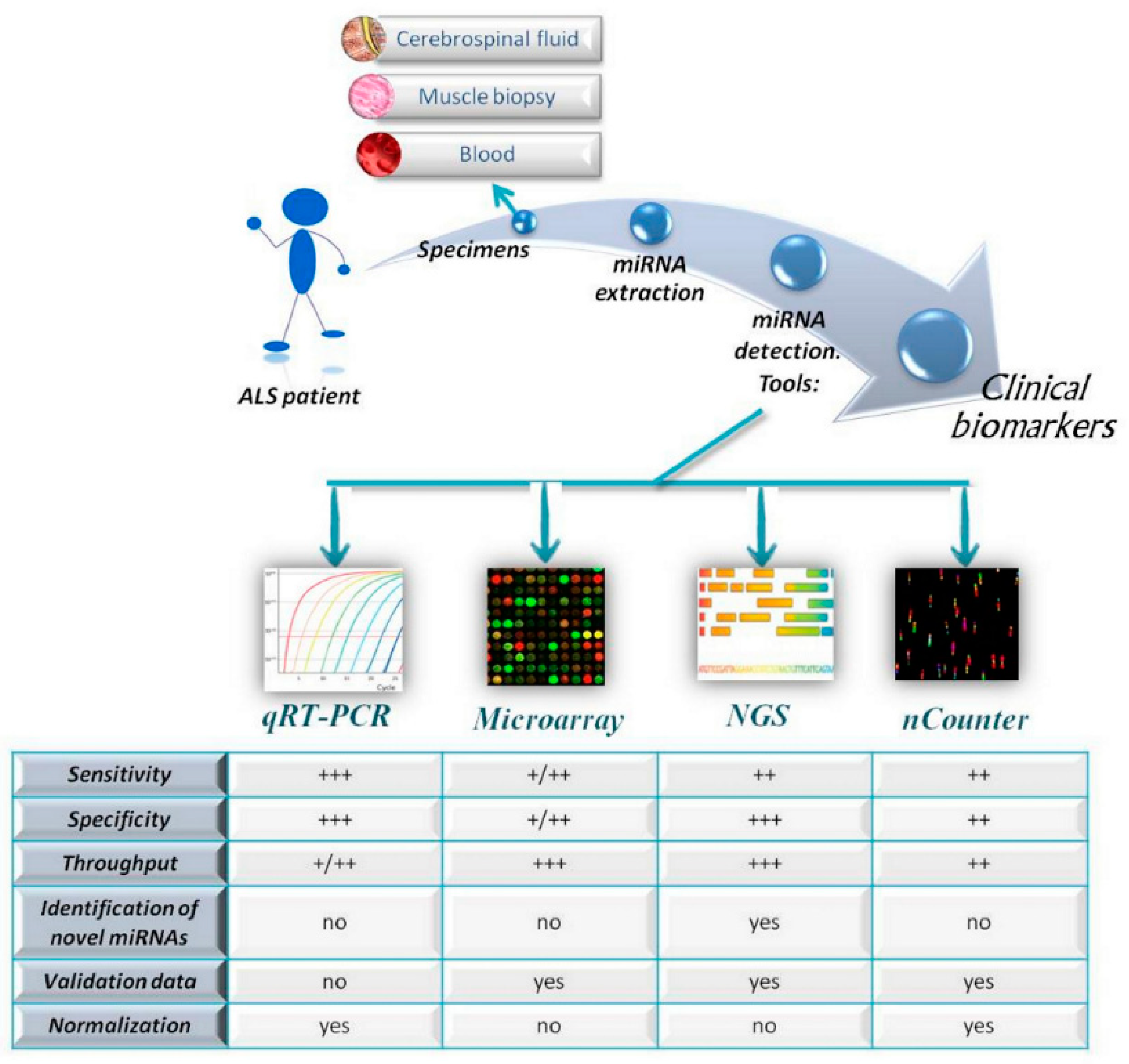
miRNA Detection
4. miRNAs as Biomarkers for ALS
4.1. miRNAs in Cerebrospinal Fluid
4.2. Circulating miRNAs
4.3. miRNAs in Muscle Biopsies
5. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Rowland, L.P.; Shneider, N.A. Amyotrophic lateral sclerosis. N. Engl. J. Med. 2001, 344, 1688–1700. [Google Scholar] [CrossRef] [PubMed]
- Rothstein, J.D. Current hypotheses for the underlying biology of amyotrophic lateral sclerosis. Ann. Neurol. 2009, 65, S3–S9. [Google Scholar] [CrossRef] [PubMed]
- Ilieva, H.; Polymenidou, M.; Cleveland, D.W. Non-cell autonomous toxicity in neurodegenerative disorders: ALS and beyond. J. Cell Biol. 2009, 187, 761–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijesekera, L.C.; Leigh, P.N. Amyotrophic lateral sclerosis. Orphanet J. Rare Dis. 2009, 4, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, R.H., Jr.; Al-Chalabi, A. Amyotrophic lateral sclerosis. N. Engl. J. Med. 2017, 377, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Chiò, A.; Logroscino, G.; Traynor, B.J.; Collins, J.; Simeone, J.C.; Goldstein, L.A.; White, L.A. Global epidemiology of amyotrophic lateral sclerosis: A systematic review of the published literature. Neuroepidemiology 2013, 41, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Logroscino, G.; Traynor, B.J.; Hardiman, O.; Chio’, A.; Couratier, P.; Mitchell, J.D.; Swingler, R.J.; Beghi, E. Descriptive epidemiology of amyotrophic lateral sclerosis: New evidence and unsolved issues. J. Neurol. Neurosurg. Psychiatry 2008, 79, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Manjaly, Z.R.; Scott, K.M.; Abhinav, K.; Wijesekera, L.; Ganesalingam, J.; Goldstein, L.H.; Janssen, A.; Dougherty, A.; Willey, E.; Stanton, B.R.; et al. The sex ratio in amyotrophic lateral sclerosis: A population based study. Amyotroph. Lateral Scler. 2010, 11, 439–442. [Google Scholar] [CrossRef] [PubMed]
- Sutedja, N.A.; Fischer, K.; Veldink, J.H.; van der Heijden, G.J.; Kromhout, H.; Heederik, D.; Huisman, M.H.; Wokke, J.J.; van den Berg, L.H. What we truly know about occupation as a risk factor for ALS: A critical and systematic review. Amyotroph. Lateral Scler. 2009, 10, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Andersen, P.M. Amyotrophic lateral sclerosis associated with mutations in the CuZn superoxide dismutase gene. Curr. Neurol. Neurosci. Rep. 2006, 6, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Ben Hamida, M.; Hentati, F.; Ben Hamida, C. Hereditary motor system diseases (chronic juvenile amyotrophic lateral sclerosis). Conditions combining a bilateral pyramidal syndrome with limb and bulbar amyotrophy. Brain 1990, 113, 347–363. [Google Scholar] [CrossRef] [PubMed]
- Andersen, P.M.; Al-Chalabi, A. Clinical genetics of amyotrophic lateral sclerosis: What do we really know? Nat. Rev. Neurol. 2011, 7, 603–615. [Google Scholar] [CrossRef] [PubMed]
- Bigio, E.H.; Weintraub, S.; Rademakers, R.; Baker, M.; Ahmadian, S.S.; Rademaker, A.; Weitner, B.B.; Mao, Q.; Lee, K.H.; Mishra, M.; Ganti, R.A.; Mesulam, M.M. Frontotemporal lobar degeneration with TDP-43 proteinopathy and chromosome 9p repeat expansion in C9ORF72: Clinicopathologic correlation. Neuropathology 2013, 33, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, F.; Riboldi, G.; Salani, S.; Nizzardo, M.; Simone, C.; Corti, S.; Hedlund, E. Cellular therapy to target neuroinflammation in amyotrophic lateral sclerosis. Cell. Mol. Life Sci. 2014, 71, 999–1015. [Google Scholar] [CrossRef] [PubMed]
- Volk, A.E.; Weishaupt, J.H.; Andersen, P.M.; Ludolph, A.C.; Kubisch, C. Current knowledge and recent insights into the genetic basis of amyotrophic lateral sclerosis. Med. Genet. 2018, 30, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, Y.; Mieda-Sato, A. TDP-43 promotes microRNA biogenesis as a component of the drosha and dicer complexes. Proc. Natl. Acad. Sci. USA 2012, 109, 3347–3352. [Google Scholar] [CrossRef] [PubMed]
- Lagier-Tourenne, C.; Polymenidou, M.; Cleveland, D.W. TDP-43 and FUS/TLS: Emerging roles in RNA processing and neurodegeneration. Hum. Mol. Genet. 2010, 19, R46–R64. [Google Scholar] [CrossRef] [PubMed]
- Brooks, B.R.; Miller, R.G.; Swash, M.; Munsat, T.L.; World Federation of Neurology Research Group on Motor Neuron Diseases. El escorial revisited: Revised criteria for the diagnosis of amyotrophic lateral sclerosis. Amyotroph. Lateral Scler. Other Motor. Neuron Disord. 2000, 1, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Zoing, M.C.; Burke, D.; Pamphlett, R.; Kiernan, M.C. Riluzole therapy for motor neurone disease: An early Australian experience (1996–2002). J. Clin. Neurosci. 2006, 13, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Biomarkers Definitions Working Group. Biomarkers and surrogate endpoints: Preferred definitions and conceptual framework. Clin. Pharmacol. Ther. 2001, 69, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Robelin, L.; Gonzalez De Aguilar, J.L. Blood biomarkers for amyotrophic lateral sclerosis: Myth or reality? Biomed. Res. Int. 2014, 2014, 10. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of microrna biogenesis, mechanisms of actions, and circulation. Front. Endocrinol. Lausanne 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, N.; Leidinger, P.; Becker, K.; Backes, C.; Fehlmann, T.; Pallasch, C.; Rheinheimer, S.; Meder, B.; Stähler, C.; Meese, A.; et al. Distribution of miRNA expression across human tissues. Nucleic Acids Res. 2016, 44, 3865–3877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mocellin, S.; Pasquali, S.; Pilati, P. Oncomirs: From tumor biology to molecularly targeted anticancer strategies. Mini Rev. Med. Chem. 2009, 9, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Aumiller, V.; Förstemann, K. Roles of microRNAs beyond development—metabolism and neural plasticity. Biochim. Biophys. Acta. 2008, 1779, 692–696. [Google Scholar] [CrossRef] [PubMed]
- Carissimi, C.; Fulci, V.; Macino, G. MicroRNAs: Novel regulators of immunity. Autoimmun. Rev. 2009, 8, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Bushati, N.; Cohen, S.M. MicroRNAs in neurodegeneration. Curr. Opin. Neurobiol. 2008, 18, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Rajgor, D. Macro roles for microRNAs in neurodegenerative diseases. Noncoding RNA Res. 2018, 3, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Rinchetti, P.; Rizzuti, M.; Faravelli, I.; Corti, S. MicroRNA Metabolism and dysregulation in amyotrophic lateral sclerosis. Mol. Neurobiol. 2018, 55, 2617–2630. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA expression profiles classify human cancers. Nature. 2005, 435, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The microRNA spectrum in 12 body fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortez, M.A.; Bueso-Ramos, C.; Ferdin, J.; Lopez-Berestein, G.; Sood, A.K.; Calin, G.A. MicroRNAs in body fluids-the mix of hormones and biomarkers. Nat. Rev. Clin. Oncol. 2011, 8, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Mo, M.H.; Chen, L.; Fu, Y.; Wang, W.; Fu, S.W. Cell-free circulating miRNA biomarkers in cancer. J. Cancer 2012, 3, 432–448. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.; Benito, E.; Fischer, A. MicroRNAs as biomarkers for CNS disease. Front. Mol. Neurosci. 2013, 6, 39. [Google Scholar] [CrossRef] [PubMed]
- Haramati, S.; Chapnik, E.; Sztainberg, Y.; Eilam, R.; Zwang, R.; Gershoni, N.; McGlinn, E.; Heiser, P.W.; Wills, A.M.; Wirguin, I.; et al. MiRNA malfunction causes spinal motor neuron disease. Proc. Natl. Acad. Sci. USA 2010, 107, 13111–13116. [Google Scholar] [CrossRef] [PubMed]
- Emde, A.; Eitan, C.; Liou, L.L.; Libby, R.T.; Rivkin, N.; Magen, I.; Reichenstein, I.; Oppenheim, H.; Eilam, R.; Silvestroni, A.; et al. Dysregulated miRNA biogenesis downstream of cellular stress and als-causing mutations: A new mechanism for ALS. EMBO J. 2015, 34, 2633–2651. [Google Scholar] [CrossRef] [PubMed]
- Strong, M.J. The evidence for altered RNA metabolism in amyotrophic lateral sclerosis (ALS). J. Neurol. Sci. 2010, 288, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Morlando, M.; Dini Modigliani, S.; Torrelli, G.; Rosa, A.; Di Carlo, V.; Caffarelli, E.; Bozzoni, I. FUS stimulates microRNA biogenesis by facilitating co-transcriptional drosha recruitment. EMBO J. 2012, 31, 4502–4510. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Ruan, K. MicroRNA detection by microarray. Anal. Bioanal. Chem. 2009, 394, 1117–1124. [Google Scholar] [CrossRef] [PubMed]
- Motameny, S.; Wolters, S.; Nürnberg, P.; Schumacher, B. Next generation sequencing of miRNAs–strategies, resources and methods. Genes 2010, 1, 70–84. [Google Scholar] [CrossRef] [PubMed]
- Friedländer, M.R.; Chen, W.; Adamidi, C.; Maaskola, J.; Einspanier, R.; Knespel, S.; Rajewsky, N. Discovering MicroRNAs from deep sequencing data using miRDeep. Nat. Biotechnol. 2008, 26, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Creighton, C.J.; Reid, J.G.; Gunaratne, P.H. Expression profiling of microRNAs by deep sequencing. Brief Bioinform. 2009, 10, 490–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mestdagh, P.; Feys, T.; Bernard, N.; Guenther, S.; Chen, C.; Speleman, F.; Vandesompele, J. High-throughput stem-loop RT-qPCR miRNA expression profiling using minute amounts of input RNA. Nucleic Acids Res. 2008, 36, e143. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Jiang, J.; Liu, Q.; Yang, L. A high-throughput method to monitor the expression of microRNA precursors. Nucleic Acids Res. 2004, 32, e43. [Google Scholar] [CrossRef] [PubMed]
- Murphy, J.; Bustin, S.A. Reliability of real-time reverse-transcription PCR in clinical diagnostics: Gold standard or substandard? Expert. Rev. Mol. Diagn. 2009, 9, 187–197. [Google Scholar] [CrossRef] [PubMed]
- De Planell-Saguer, M.; Rodicio, M.C. Detection methods for microRNAs in clinic practice. Clin. Biochem. 2013, 46, 869–878. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenbach, H.; da Silva, A.M.; Calin, G.; Pantel, K. Data normalization strategies for microRNA quantification. Clin. Chem. 2015, 61, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Peltier, H.J.; Latham, G.J. Normalization of microRNA expression levels in quantitative RT-PCR assays: Identification of suitable reference RNA targets in normal and cancerous human solid tissues. RNA 2008, 14, 844–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kroh, E.M.; Parkin, R.K.; Mitchell, P.S.; Tewari, M. Analysis of circulating microRNA biomarkers in plasma and serum using quantitative reverse transcription-PCR (qRT-PCR). Methods 2010, 50, 298–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Felice, B.; Guida, M.; Guida, M.; Coppola, C.; De Mieri, G.; Cotrufo, R. A miRNA signature in leukocytes from sporadic amyotrophic lateral sclerosis. Gene 2012, 508, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Cogswell, J.P.; Ward, J.; Taylor, I.A.; Waters, M.; Shi, Y.; Cannon, B.; Kelnar, K.; Kemppainen, J.; Brown, D.; Chen, C.; et al. Identification of miRNA changes in Alzheimer’s disease brain and CSF yields putative biomarkers and insights into disease pathways. J. Alzheimers Dis. 2008, 14, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Alexandrov, P.N.; Dua, P.; Hill, J.M.; Bhattacharjee, S.; Zhao, Y.; Lukiw, W.J. microRNA (miRNA) speciation in Alzheimer’s disease (AD) cerebrospinal fluid (CSF) and extracellular fluid (ECF). Int. J. Biochem. Mol. Biol. 2012, 3, 365–373. [Google Scholar] [PubMed]
- Butovsky, O.; Siddiqui, S.; Gabriely, G.; Lanser, A.J.; Dake, B.; Murugaiyan, G.; Doykan, C.E.; Wu, P.M.; Gali, R.R.; Iyer, L.K.; et al. Modulating inflammatory monocytes with a unique microRNA gene signature ameliorates murine ALS. J. Clin. Invest. 2012, 122, 3063–3087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freischmidt, A.; Müller, K.; Ludolph, A.C.; Weishaupt, J.H. Systemic dysregulation of TDP-43 binding microRNAs in amyotrophic lateral sclerosis. Acta Neuropathol. Commun. 2013, 1, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Felice, B.; Annunziata, A.; Fiorentino, G.; Borra, M.; Biffali, E.; Coppola, C.; Cotrufo, R.; Brettschneider, J.; Giordana, M.L.; Dalmay, T.; et al. MiR-338-3p is over-expressed in blood, CFS, serum and spinal cord from sporadic amyotrophic lateral sclerosis patients. Neurogenetics 2014, 15, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Benigni, M.; Ricci, C.; Jones, A.R.; Giannini, F.; Al-Chalabi, A.; Battistini, S. Identification of miRNAs as potential biomarkers in cerebrospinal fluid from amyotrophic lateral sclerosis patients. Neuromolecular Med. 2016, 18, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Waller, R.; Wyles, M.; Heath, P.R.; Kazoka, M.; Wollff, H.; Shaw, P.J.; Kirby, J. Small RNA sequencing of sporadic amyotrophic lateral sclerosis cerebrospinal fluid reveals differentially expressed miRNAs related to neural and glial activity. Front. Neurosci. 2018, 11, 731. [Google Scholar] [CrossRef] [PubMed]
- Paez-Colasante, X.; Figueroa-Romero, C.; Sakowski, S.A.; Goutman, S.A.; Feldman, E.L. Amyotrophic lateral sclerosis: mechanisms and therapeutics in the epigenomic era. Nat. Rev. Neurol. 2015, 11, 266–279. [Google Scholar] [CrossRef] [PubMed]
- Hoye, M.L.; Koval, E.D.; Wegener, A.J.; Hyman, T.S.; Yang, C.; O’Brien, D.R.; Miller, R.L.; Cole, T.; Schoch, K.M.; Shen, T.; et al. MicroRNA profiling reveals marker of motor neuron disease in ALS models. J. Neurosci. 2017, 37, 5574–5586. [Google Scholar] [CrossRef] [PubMed]
- Kos, A.; Olde Loohuis, N.F.; Wieczorek, M.L.; Glennon, J.C.; Martens, G.J.; Kolk, S.M.; Aschrafi, A. A potential regulatory role for intronic microRNA-338-3p for its host gene encoding apoptosis-associated tyrosine kinase. PLoS ONE 2012, 7, e31022. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Pan, M.; Han, L.; Lu, H.; Hao, X.; Dong, Q. miR-338-3p suppresses neuroblastoma proliferation, invasion and migration through targeting PREX2a. FEBS Lett. 2013, 587, 3729–3737. [Google Scholar] [CrossRef] [PubMed]
- Conti, A.; Aguennouz, M.; La Torre, D.; Tomasello, C.; Cardali, S.; Angileri, F.F.; Maio, F.; Cama, A.; Germanò, A.; Vita, G.; et al. miR-21 and 221 upregulation and miR-181b downregulation in human grade II-IV astrocytic tumors. J. Neurooncol. 2009, 93, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Johanson, C.E.; Stopa, E.G.; McMillan, P.N. The blood-cerebrospinal fluid barrier: Structure and functional significance. Methods Mol. Biol. 2011, 686, 101–131. [Google Scholar] [CrossRef] [PubMed]
- Spector, R.; Robert Snodgrass, S.; Johanson, C.E. A balanced view of the cerebrospinal fluid composition and functions: Focus on adult humans. Exp. Neurol. 2015, 273, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Vu, L.T.; Bowser, R. Fluid-based biomarkers for amyotrophic lateral sclerosis. Neurotherapeutics 2017, 14, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Toivonen, J.M.; Manzano, R.; Oliván, S.; Zaragoza, P.; García-Redondo, A.; Osta, R. MicroRNA-206: A potential circulating biomarker candidate for amyotrophic lateral sclerosis. PLoS ONE. 2014, 9, e89065. [Google Scholar] [CrossRef] [PubMed]
- Freischmidt, A.; Müller, K.; Zondler, L.; Weydt, P.; Volk, A.E.; Božič, A.L.; Walter, M.; Bonin, M.; Mayer, B.; von Arnim, C.A.; et al. Serum microRNAs in patients with genetic amyotrophic lateral sclerosis and pre-manifest mutation carriers. Brain 2014, 137, 2938–2950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freischmidt, A.; Müller, K.; Zondler, L.; Weydt, P.; Mayer, B.; von Arnim, C.A.; Hübers, A.; Dorst, J.; Otto, M.; Holzmann, K.; et al. Serum microRNAs in sporadic amyotrophic lateral sclerosis. Neurobiol. Aging 2015, 36, 2660.e15–2660.e20. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, I.; Hama, Y.; Matsushima, M.; Hirotani, M.; Kano, T.; Hohzen, H.; Yabe, I.; Utsumi, J.; Sasaki, H. Identification of plasma microRNAs as a biomarker of sporadic amyotrophic lateral Sclerosis. Mol. Brain 2015, 8, 67. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wei, Q.; Chen, X.; Li, C.; Cao, B.; Ou, R.; Hadano, S.; Shang, H.F. Aberration of miRNAs expression in leukocytes from sporadic amyotrophic lateral sclerosis. Front. Mol. Neurosci. 2016, 9, 69. [Google Scholar] [CrossRef] [PubMed]
- De Andrade, H.M.; de Albuquerque, M.; Avansini, S.H.; de S Rocha, C.; Dogini, D.B.; Nucci, A.; Carvalho, B.; Lopes-Cendes, I.; França, M.C., Jr. MicroRNAs-424 and 206 are potential prognostic markers in spinal onset amyotrophic lateral sclerosis. J. Neurol. Sci. 2016, 368, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Tasca, E.; Pegoraro, V.; Merico, A.; Angelini, C. circulating microRNAs as biomarkers of muscle differentiation and atrophy in ALS. Clin. Neuropathol. 2016, 35, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Sheinerman, K.S.; Toledo, J.B.; Tsivinsky, V.G.; Irwin, D.; Grossman, M.; Weintraub, D.; Hurtig, H.I.; Chen-Plotkin, A.; Wolk, D.A.; McCluskey, L.F.; et al. Circulating brain-enriched microRNAs as novel biomarkers for detection and differentiation of neurodegenerative diseases. Alzheimers Res. Ther. 2017, 9, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raheja, R.; Regev, K.; Healy, B.C.; Mazzola, M.A.; Beynon, V.; Von Glehn, F.; Paul, A.; Diaz-Cruz, C.; Gholipour, T.; Glanz, B.I.; et al. Correlating serum micrornas and clinical parameters in amyotrophic lateral sclerosis. Muscle Nerve 2018, 58, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Waller, R.; Goodall, E.F.; Milo, M.; Cooper-Knock, J.; Da Costa, M.; Hobson, E.; Kazoka, M.; Wollff, H.; Heath, P.R.; Shaw, P.J. Serum miRNAs miR-206, 143–3p and 374b-5p as potential biomarkers for amyotrophic lateral sclerosis (ALS). Neurobiol. Aging 2017, 55, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Vrabec, K.; Boštjančič, E.; Koritnik, B.; Leonardis, L.; Dolenc Grošelj, L.; Zidar, J.; Rogelj, B.; Glavač, D.; Ravnik-Glavač, M. Differential expression of several miRNAs and the host genes AATK and DNM2 in leukocytes of sporadic ALS patients. Front. Mol. Neurosci. 2018, 11, 106. [Google Scholar] [CrossRef] [PubMed]
- Matamala, J.M.; Arias-Carrasco, R.; Sanchez, C.; Uhrig, M.; Bargsted, L.; Matus, S.; Maracaja-Coutinho, V.; Abarzua, S.; van Zundert, B.; Verdugo, R. Genome-wide circulating microRNA expression profiling reveals potential biomarkers for amyotrophic lateral sclerosis. Neurobiol. Aging 2018, 64, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Zhao, Y.; Zhou, X.; Luan, J.; Cui, Y.; Han, J. Comparison of the extraction and determination of serum exosome and miRNA in serum and the detection of miR-27a-3p in serum exosome of ALS patients. Intractable Rare Dis. Res. 2018, 7, 13–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liguori, M.; Nuzziello, N.; Introna, A.; Consiglio, A.; Licciulli, F.; D’Errico, E.; Scarafino, A.; Distaso, E.; Simone, I.L. Dysregulation of MicroRNAs and target genes networks in peripheral blood of patients with sporadic amyotrophic lateral sclerosis. Front. Mol. Neurosci. 2018, 11, 288. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, Y.H.; Wang, H. Exploring microRNA biomarker for amyotrophic lateral sclerosis. Int. J. Mol. Sci. 2018, 19, 1318. [Google Scholar] [CrossRef] [PubMed]
- Bang, C.; Thum, T. Exosomes: New players in cell-cell communication. Int. J. Biochem. Cell. Biol. 2012, 44, 2060–2064. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Luan, J.; Li, H.; Zhou, X.; Han, J. Exosomes derived from mineralizing osteoblasts promote ST2 cell osteogenic differentiation by alteration of microRNA expression. FEBS Lett. 2016, 590, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Juvvuna, P.K.; Kukreti, H.; McFarlane, C. Mega roles of microRNAs in regulation of skeletal muscle health and disease. Front. Physiol. 2014, 5, 239. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Wang, Y.; Li, Y.; Cui, L.; Zhao, Y.; Zhao, B.; Li, K. MiR-206, a key modulator of skeletal muscle development and disease. Int. J. Biol. Sci. 2015, 11, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.H.; Valdez, G.; Moresi, V.; Qi, X.; McAnally, J.; Elliott, J.L.; Bassel-Duby, R.; Sanes, J.R.; Olson, E.N. MicroRNA-206 delays ALS progression and promotes regeneration of neuromuscular synapses in mice. Science 2009, 326, 1549–1554. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, J.P.; Picchiarelli, G.; Dupuis, L.; Gonzalez De Aguilar, J.L. The role of skeletal muscle in amyotrophic lateral sclerosis. Brain Pathol. 2016, 26, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Horak, M.; Novak, J.; Bienertova-Vasku, J. Muscle-specific microRNAs in skeletal muscle development. Dev. Biol. 2016, 410, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Di Pietro, L.; Lattanzi, W.; Bernardini, C. Skeletal muscle MicroRNAs as key players in the pathogenesis of amyotrophic lateral sclerosis. Int. J. Mol. Sci. 2018, 19, 1534. [Google Scholar] [CrossRef] [PubMed]
- Bruneteau, G.; Simonet, T.; Bauché, S.; Mandjee, N.; Malfatti, E.; Girard, E.; Tanguy, M.L.; Behin, A.; Khiami, F.; Sariali, E.; et al. Muscle histone deacetylase 4 upregulation in amyotrophic lateral sclerosis: potential role in reinnervation ability and disease progression. Brain 2013, 136, 2359–2368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, A.P.; Wada, S.; Vergani, L.; Hock, M.B.; Lamon, S.; Léger, B.; Ushida, T.; Cartoni, R.; Wadley, G.D.; Hespel, P.; et al. Disruption of skeletal muscle mitochondrial network genes and miRNAs in amyotrophic lateral sclerosis. Neurobiol. Dis. 2013, 49, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.; Jørgensen, L.H.; Bech, R.D.; Frandsen, U.; Schrøder, H.D. Skeletal muscle remodelling as a function of disease progression in amyotrophic lateral sclerosis. Biomed. Res. Int. 2016, 2016, 5930621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pegoraro, V.; Merico, A.; Angelini, C. Micro-RNAs in ALS muscle: Differences in gender, age at onset and disease duration. J. Neurol. Sci. 2017, 380, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Si, Y.; Cui, X.; Crossman, D.K.; Hao, J.; Kazamel, M.; Kwon, Y.; King, PH. Muscle microRNA signatures as biomarkers of disease progression in amyotrophic lateral sclerosis. Neurobiol. Dis. 2018, 114, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Kovanda, A.; Leonardis, L.; Zidar, J.; Koritnik, B.; Dolenc-Groselj, L.; Ristic Kovacic, S.; Curk, T.; Rogelj, B. Differential expression of microRNAs and other small RNAs in muscle tissue of patients with ALS and healthy age-matched controls. Sci. Rep. 2018, 8, 5609. [Google Scholar] [CrossRef] [PubMed]
- Di Pietro, L.; Baranzini, M.; Berardinelli, M.G.; Lattanzi, W.; Monforte, M.; Tasca, G.; Conte, A.; Logroscino, G.; Michetti, F.; Ricci, E.; et al. Potential therapeutic targets for ALS: MIR206, MIR208b and MIR499 are modulated during disease progression in the skeletal muscle of patients. Sci. Rep. 2017, 7, 9538. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| miRNA (Hsa-miR) | miRNA Expression Change | No. of Specimens | miRNA Detection Approach | Ref. |
|---|---|---|---|---|
| 150, 99b, 146a 27b, 328, 532-3p | ↑ in SALS ↑ in SALS and FALS | SALS: 10 FALS: 5 HCs: 10 | qRT-PCR | [54] |
| 132-5p, 132-3p, 143-3p 143-5p, 574-5p | ↓ ↑ | SALS: 22 HCs: 24 | qRT-PCR | [55] |
| 338-3p | ↑ | SALS: 10 HCs: 10 | qRT-PCR | [56] |
| 181a-5p 21-5p, 195-5p, 148-3p, 15b-5p, let7a-5p, let7b-5p, let7f-5p | ↑ ↓ | SALS: 24 HCs: 24 | qRT-PCR | [57] |
| 124-3p, 127-3p, 143-3p, 125b-2-3p, 9-5p, 27b-3p 486-5p, let7f-5p, 16-5p, 28-3p, 146a-3p, 150-5p, 378a-3p, 142-5p, 92a-5p | ↑ ↓ | SALS: 32 HCs: 10 NCs: 6 | NGS | [58] |
| miRNA (Has-miR) Expression Change | Source | miRNA Detection Approach | No. of Specimens for miRNAs Validation | Ref. |
|---|---|---|---|---|
↑: 338-3p | Leukocytes | Microarray→ miRNAs validation with qRT-PCR | SALS: 14 HCs: 14 | [51] |
| ↑: 27a, 155, 142-5p, 223, 30b, 532-3p | Monocytes (CD14+ CD16-) | Nanostring nCounter 1 → miRNAs validation with qRT-PCR | SALS: 22 FALS: 4 HCs: 24 | [54] |
| ↓: 132-3p, 132-5p, 143-3p, 143-5p, let-7b | Serum | Nine TARDBP binding miRNAs and miR-9-5p → qRT-PCR | SALS: 22 HCs: 24 | [55] |
| ↑: 206, 106b | Serum | Microarray 1 → miRNAs validation with qRT-PCR | SALS: 12 HCs: 12 | [67] |
| ↑: 338-3p | Leukocytes and serum | miR-338-3p → qRT-PCR | SALS: 10 HCs: 10 | [56] |
| ↓ in FALS/SALS: 4745-5p, 3665, 4530 ↓ in FALS: 1915-3p | Serum | Microarray → miRNAs validation with qRT-PCR | FALS: 23 HCs: 24 SALS: 14 HCs: 14 | [68] |
| ↓ in FALS/SALS: 1825 ↓ in SALS: 1234-3p | Serum | Microarray→ miRNAs validation with qRT-PCR | SALS: 20 HCs: 20 FALS: 13 HCs: 13 | [69] |
| ↑: 4649-5p ↓: 4299 | Plasma | Microarray → miRNAs validation with qRT-PCR | SALS: 48 HCs: 47 | [70] |
| ↓: 183, 193b, 451, 3935 | Leukocytes | Microarray → miRNAs validation with qRT-PCR | SALS: 83 HCs: 61 | [71] |
| ↑: 424, 206 | Plasma | Microarray 2 → miRNAs validation with qRT-PCR | SALS: 39 HCs: 39 | [72] |
| ↑: 206, 133a,133b ↓: 146a, 149*, 27a | Serum | Preselected myo-miRNAs, inflammatory and angiogenic miRNA → qRT-PCR | SALS: 14 HCs: 8 | [73] |
| ↑: 206 Deregulated MicroRN pairs: 206/338-3p 9*/129-3p 335-5p/338-3p | Plasma | Thirty seven brain-enriched and inflammation-associated microRNAs → qRT-PCR | ALS: 50 HCs: 50 | [74] |
| ↑ ┼: 1, 133a-3p, 133b, 144-5p, 192-3p, 195-5p, 19a-3p ↓ ┼: let-7d-3p, 320a, 320b, 320c, 425-5p, 139-5p | Serum | qRT-PCR array | SALS: 20 FALS: 3 HCs: 30 NCs: 103 | [75] |
| ↑: 206, 143-3p ↓: 374b-5p | Serum | qRT-PCR array → miRNAs validation with qRT-PCR | SALS: 23 CRL: 22 | [76] |
| ↑: 9, 338, 638, 663a, 124a, 451a, 132, 206, let-7b | Leukocytes | Preselected 10 miRNAs → miRNAs validation with qRT-PCR | SALS: 84 HCs: 27 | [77] |
| ↑: 142-3p ↓: 1249-3p | Serum | Microarray 1 → miRNAs validation with qRT-PCR | SALS: 20 HCs: 20 | [78] |
| ↓: 27a-3p | Serum exosomes | miR-27a-3p → qRT-PCR | ALS: 10 HCs: 20 | [79] |
| ↓: let-7a-5p, let-7d-5p, let-7f-5p, let-7g-5p, let-7i-5p, 103a-3p, 106b-3p, 128-3p, 130a-3p, 130b-3p, 144-5p, 148a-3p, 148b-3p, 15a-5p, 15b-5p, 151a-5p, 151b, 16-5p, 182-5p, 183-5p, 186-5p, 22-3p, 221-3p, 223-3p, 23a-3p, 26a-5p, 26b-5p, 27b-3p, 28-3p, 30b-5p, 30c-5p, 342-3p, 425-5p, 451a, 532-5p, 550a-3p, 584-5p, 93-5p | Whole blood | NGS → qRT-PCR | SALS: 50 HCs: 15 | [80] |
| miRNAs (Has-miR) | miRNA Change | Role in ALS | Ref. |
|---|---|---|---|
| 206 | ↑ | Myo-miRNA: muscle proliferation, repair and regeneration. It promotes neuromuscular connectivity and enhances reinnervation | [67,72,73,74,76,77] |
| 338 | ↑ | Involvement in different pathways such as apoptosis, neurodegeneration, and/or glutamate clearance | [51,67,74,77] |
| 133a | ↑ | Myo-miRNA: muscle proliferation, repair and regeneration | [73,75] |
| 133b | ↑ | Myo-miRNA: muscle proliferation, repair and regeneration | [73,75] |
| 142 | ↑ | miRNA predicted to target a specific set of genes associated to the pathophysiology of ALS, including TARDBP and C9orf72. | [54,78] |
| 183 | ↓ | miRNA involved in neurodegenerative signaling pathway, including PI3K-Akt and MAPK pathway. miR-183/mTOR pathway contributes to spinal muscular atrophy pathology | [71,80] |
| 27a | ↓ | miRNA involved in muscle growth, myoblast proliferation acting on myostatin. It is present in myoblast-derived exosomes | [73,79] |
| let-7d | ↓ | Involvement in apoptosis by the Hippo signaling pathway | [75,80] |
| miRNA (Hsa-miR) | miRNA Expression Change | Type of Muscle | No. of Muscle Biopsies | miRNA Detection Approach | Ref. |
|---|---|---|---|---|---|
| 206 | ↑ | Deltoid, anconeus | FALS: 1 SALS: 10 HCs: 6 | mir-206 → qRT-PCR | [90] |
| 23a, 29b, 206, 455, 31 | ↑ | Vastus lateralis | ALS: 14 HCs: 10 | Myo-miRNAs and miRNAs dysregulated in human muscle disease → qRT-PCR | [91] |
| 1, 26a, 133a, 455 | ↓ | Vastus lateralis | ALS: 5 HCs: 7 | Myo-miRNAs → qRT-PCR | [92] |
| 424, 214, 206 | ↑ | Biceps brachii | ALS: 5 HCs: 5 | Microarray → miRNAs validation with qRT-PCR | [72] |
| 1, 206, 133a, 133b, 27a, 155, 146a, 221 | ↑ | Quadriceps femoris | SALS: 13 HCs: 5 | Inflammatory/angiogenic miRNAs and myo-miRNAs → qRT-PCR | [93] |
| 1, 10b-5p, 100-5p, 133a-3p, 133b-3p | ↓ | Biceps, deltoid, tibialis anterior, vastus lateralis | ALS: 19 HCs: 9 | NGS 1 and qRT-PCR 1 → qRT-PCR | [94] |
| 100-5p, 10a, 125a-5p, 133a-1/-2-3p, 362, 500a-3p, 542-5p, 99a-5p 1303-3p, 150-5p, 26a-1/-2-5p, 486-1/-2-5p, | ↑ ↓ | Vastus lateralis | FALS: 2 SALS: 9 HCs: 11 | NGS | [95] |
| miRNA (Hsa-miR) | miRNA Expression Change in Specific ALS Patients’ Group | Type of Muscle | No. of Muscle Biopsies | miRNA Detection Approach | Ref. |
|---|---|---|---|---|---|
| 133a, 29c, 9, 208b 1, 208b 133a, 133b, 206, 29c, 9, 155, 23a | ↑in ALS slow group 1 ↓ in ALS rapid group 2 ↑ in early stage group 3 | Deltoid and quadriceps | FALS: 3 SALS: 11 HCs: 24 Slow group 1: 6 Rapid group 2: 5 Early group 3: 4 Late group 4:9 | Eleven skeletal muscle related miRNAs → qRT-PCR | [96] |
| 100-5p, 199a-1/-2, 199b-3p, 27a-5p, 3607-3p, 424-5p, 450a-1/-2-5p, 450b-5p, 501-3p, 502-3p, 542-5p, 660-5p 1303-3p, 133a-1/-2-3p, 150-5p, 378, 486-1/-2-5p, 502-3p, 855-3p | ↑ in higher disease severity 5 ↓ in higher disease severity 5 | Vastus lateralis | Higher disease group 5: 7 HCs: 11 | NGS | [95] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ricci, C.; Marzocchi, C.; Battistini, S. MicroRNAs as Biomarkers in Amyotrophic Lateral Sclerosis. Cells 2018, 7, 219. https://doi.org/10.3390/cells7110219
Ricci C, Marzocchi C, Battistini S. MicroRNAs as Biomarkers in Amyotrophic Lateral Sclerosis. Cells. 2018; 7(11):219. https://doi.org/10.3390/cells7110219
Chicago/Turabian StyleRicci, Claudia, Carlotta Marzocchi, and Stefania Battistini. 2018. "MicroRNAs as Biomarkers in Amyotrophic Lateral Sclerosis" Cells 7, no. 11: 219. https://doi.org/10.3390/cells7110219
APA StyleRicci, C., Marzocchi, C., & Battistini, S. (2018). MicroRNAs as Biomarkers in Amyotrophic Lateral Sclerosis. Cells, 7(11), 219. https://doi.org/10.3390/cells7110219