Aquaporin Activity to Improve Crop Drought Tolerance


Abstract
: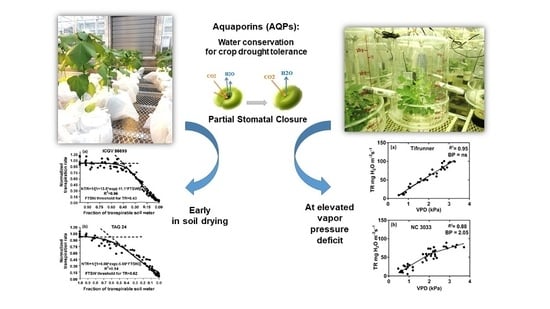
{kind=link}
{kind=link}
{kind=link}
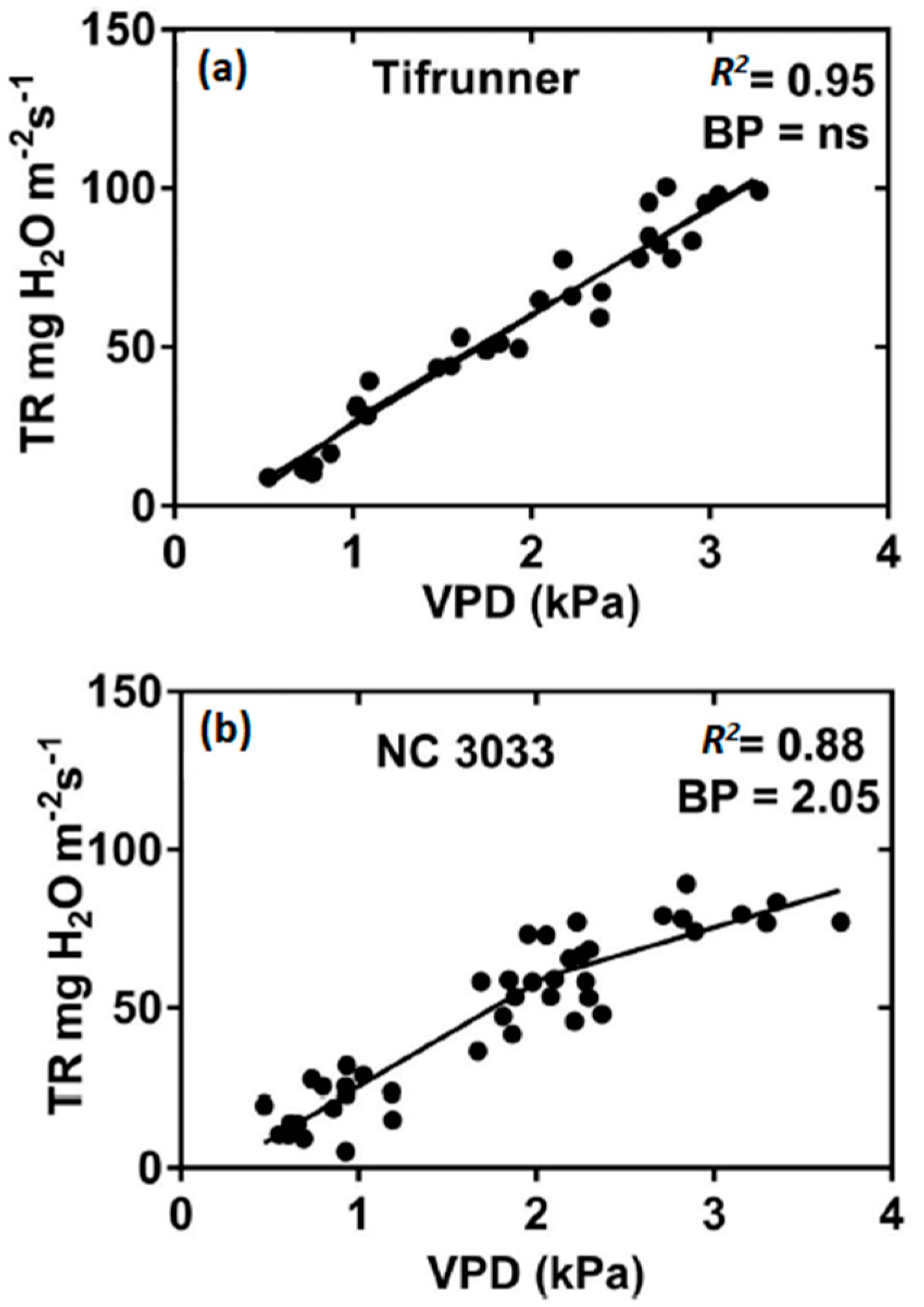
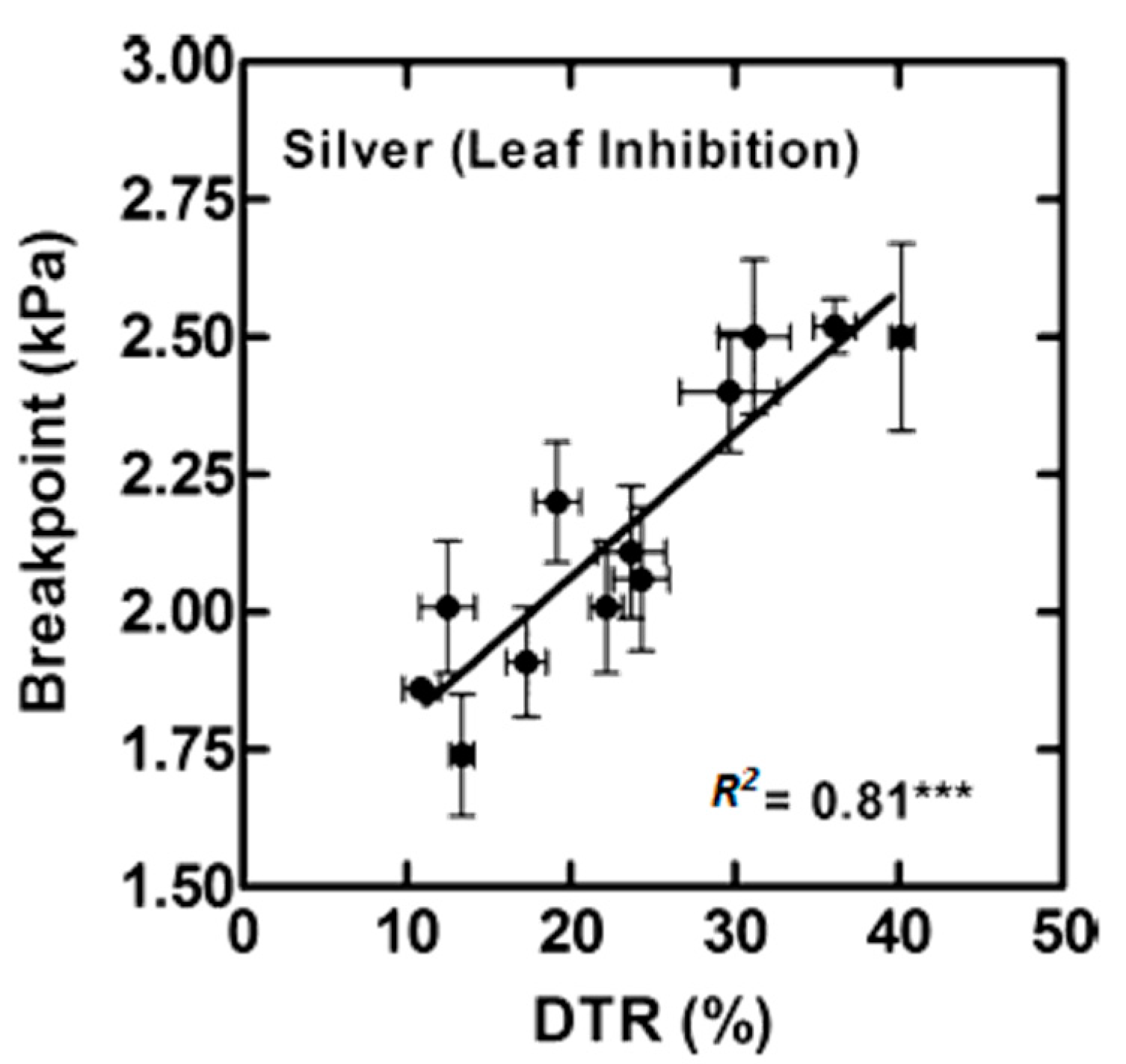{kind=link}
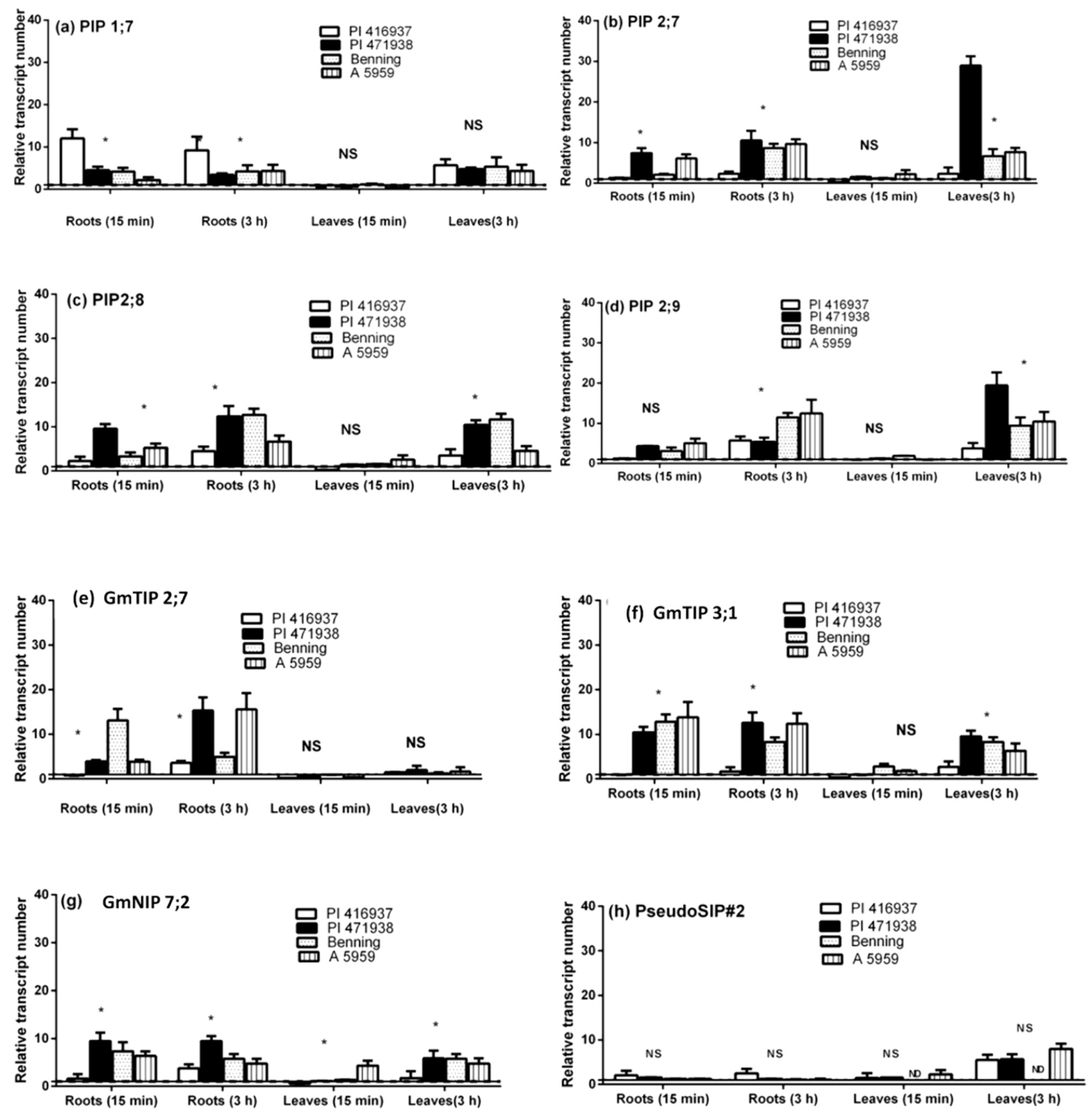{kind=link}
1. Introduction
2. Plant Water Use
3. Hydraulic Conductance
4. Soil Drying
5. High Evaporative Demand [or High Vapor Pressure Deficit (VPD)]
6. Molecular Observations
7. Conclusions
Funding
Conflicts of Interest
References
- Li, J.; Ban, L.; Wen, H.; Wang, Z.; Dzyubenko, N.; Chapurin, V.; Gao, H.; Wang, X. An aquaporin protein is associated with drought stress tolerance. Biochem. Biophys. Res. Commun. 2015, 459, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.; Gho, C.; Leafgren, R.; Tang, T.; Messina, C.D. Breeding drought-tolerant maize hybrids for the US corn-belt: Discovery to product. J. Exp. Bot. 2014, 65, 6191–6204. [Google Scholar] [CrossRef] [PubMed]
- Federal Emergency Management Agency. National Mitigation Strategy—Partnerships for Building Safer Communities; Mitigation Directorate; FEMA: Washington, DC, USA, 1995.
- Zhao, C.X.; Shao, H.B.; Chu, L.Y. Aquaporin structure-function relationships: Water flow through plant living cells. Colloid Surf. B Biointerfaces 2008, 62, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Steudle, E.; Schulze, E. The regulation of plant water at the cell, tissue, and organ level: Role of active processes and of compartmentation. In Flux Control in Biological Systems: From Enzymes to Populations and Ecosystems; Schultze, E.D., Ed.; Academic Press: San Diego, CA, USA, 1994; pp. 237–299. [Google Scholar]
- Siefritz, F.; Tyree, M.T.; Lovisolo, C.; Schubert, A.; Kaldenhoff, R. PIP1 plasma membrane aquaporins in tobacco: From cellular effects to function in plants. Plant Cell 2002, 4, 869–876. [Google Scholar] [CrossRef]
- Aharon, A.; Shahak, Y.; Wininger, S.; Bendov, R.; Kapulnik, Y.; Galili, G. Overexpression of a plasma membrane aquaporin in transgenic tobacco improves plant vigor under favorable growth conditions but not under drought or salt stress. Plant Cell 2003, 15, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Moshelion, M.; Halperin, F.; Wallach, R.; Oren, R.; Way, D.A. Role of aquaporins in determining transpiration and photosynthesis in water-stressed plants: Crop water-use efficiency, growth and yield. Plant Cell Environ. 2015, 38, 1785–1793. [Google Scholar] [CrossRef] [PubMed]
- Sakuarai-Ishikawa, J.; Mursi-Hatano, M.; Hayashi, H.; Ahamed, A.; Fukushi, K.; Matsumoto, T.; Kitagawa, Y. Transpiration from shoots triggers diurnal changes in root aquaporin expression. Plant Cell Environ. 2011, 34, 1150–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinclair, T.R. Challenges in breeding for yield increase for drought. Trends Plant Sci. 2011, 16, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, T.R. Water-Conservation Traits to Increase Crop Yields in Water-Deficit Environments: Case Studies; Springer Briefs in Environmental Science; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Sinclair, T.R.; Messina, C.D.; Beatty, A.; Samples, A. Assessment across the United States of the benefits of altered soybean drought traits. Agron. J. 2010, 102, 475–482. [Google Scholar] [CrossRef]
- Messina, C.D.; Sinclair, T.R.; Hammer, G.L.; Curan, D.; Thompson, J.; Oler, Z.; Gho, C.; Cooper, C. Limited-Transpiration Trait May Increase Maize Drought Tolerance in the US Corn Belt. Agron. J. 2015, 107, 1978–1986. [Google Scholar] [CrossRef]
- Lian, H.L.; Yu, X.; Lane, D.; Sun, W.N.; Tang, Z.C.; Su, W.A. Upland rice and lowland rice exhibited different PIP expression under water deficit and ABA treatment. Cell Res. 2006, 16, 651–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaman-Allah, M.; Jenkinson, D.M.; Vadez, V. Chickpea genotypes contrasting for seed yield under terminal drought stress in the field differ for traits related to the control of water use. Funct. Plant Biol. 2011, 38, 270–281. [Google Scholar] [CrossRef] [Green Version]
- Shekoofa, A.; Rosas-Anderson, P.; Sinclair, T.R.; Balota, M.; Isleib, T.G. Measurement of limited-transpiration trait under high vapor pressure deficit for peanut in chambers and in field. Agron. J. 2015, 107, 1019–1024. [Google Scholar] [CrossRef]
- Shekoofa, A.; Sinclair, T.R.; Messina, C.D.; Cooper, M. Variation among maize hybrids in response to high vapor pressure deficit at high temperatures. Crop Sci. 2016, 55, 392–396. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Devi, J.; Shekoofa, A.; Choudhary, S.; Sadok, W.; Vadez, V.; Riar, M.; Rufty, T. Limited-transpiration response to high vapor pressure deficit in crop species. Plant Sci. 2017, 260, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Aroca, R.; Porcel, R.; Ruiz-Lozano, J.M. Regulation of root water uptake under abiotic stress conditions. J. Exp. Bot. 2012, 63, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Pou, A.; Medrano, H.; Flexas, J.; Tyerman, S.D. A putative role for TIP and PIP aquaporins in dynamics of leaf hydraulic and stomatal conductances in grapevine under water stress and re-watering. Plant Cell Environ. 2013, 36, 828–843. [Google Scholar] [CrossRef] [PubMed]
- Bunce, J.A. How do leaf hydraulics limit stomatal conductance at high water vapour pressure deficits? Plant Cell Environ. 2006, 29, 1644–1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Sperry, J.S.; Shao, M. Hydraulic conductance and vulnerability to cavitation in corn (Zea mays L.) hybrids of differing drought resistance. Environ. Exp. Bot. 2009, 66, 341–346. [Google Scholar] [CrossRef]
- Pangle, R.E.; Limousin, J.M.; Plaut, J.A.; Yepez, E.A.; Hudson, P.J.; Boutz, A.I.; Gehres, N.; Pockman, W.T.; McDowell, N.G. Prolonged experimental drought reduces plant hydraulic conductance and transpiration and increases mortality in a pinon-juniper woodland. Ecol. Evol. 2015, 5, 1618–1638. [Google Scholar] [CrossRef] [PubMed]
- Trillo, N.; Fernandez, R.J. Wheat plant hydraulic properties under prolonged experimental drought: Stronger decline to root system conductivity than in leaf area. Plant Soil 2005, 277, 277–284. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Zwieniecki, M.A.; Holbrook, N.M. Low leaf hydraulic conductance associated with drought tolerance in soybean. Phys. Plant 2008, 132, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Schoppach, R.; Wauthelet, D.; Jeanguenin, L.; Sadok, W. Conservative water use under high evaporative demand associated with smaller root metaxylem and limited transmembrane water transport in wheat. Funct. Plant Biol. 2014, 41, 257–269. [Google Scholar] [CrossRef]
- Cochard, H.; Venisse, J.S.; Barigah, T.S.; Brunel, N.; Herbette, S.; Guilliot, A.; Tyree, M.T.; Sakr, S. Putative role of aquaporins in variable hydraulic conductance of leaves in response to light. Plant Physiol. 2007, 143, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Locke, A.M.; Ort, D.R. Diurnal depression in leaf hydraulic conductance at ambient and elevated [CO2] reveals anisohydric water management in field-grown soybean and possible involvement of aquaporins. Environ. Exp. Bot. 2015, 116, 39–46. [Google Scholar] [CrossRef]
- Choudhary, S.; Sinclair, T.R.; Messina, C.D. Hydraulic conductance of maize hybrids differing in transpiration response to vapor pressure deficit. Crop Sci. 2014, 54, 1147–1152. [Google Scholar]
- Sinclair, T.R.; Shekoofa, A.; Isleib, T.G.; Balota, M.; Hou, Z. Identification of virginia-type peanut genotypes for water-deficit conditions based on early stomatal closure with soil drying. Crop Sci. 2019. in review. [Google Scholar]
- Sinclair, T.R. Theoretical analysis of soil and plant traits influencing daily plant water flux on drying soils. Agron. J. 2005, 97, 1148–1152. [Google Scholar] [CrossRef]
- Devi, J.; Sinclair, T.R.; Vadez, V.; Krishnamurthy, L. Peanut genotypic variation in transpiration efficiency and decreased transpiration during progressive soil drying. Field Crops Res. 2009, 114, 280–285. [Google Scholar] [CrossRef]
- Nada, R.M.; Abogadallah, G.M. Aquaporins are major determinants of water use efficiency of rice plants in the field. Plant Sci. 2014, 227, 165–180. [Google Scholar] [CrossRef] [PubMed]
- Kaldenhoff, R.; Ribas-Carbo, M.; Flexas Sans, J.; Lovisolo, C.; Heckwolf, M.; Uehlein, N. Aquaporins and plant water balance. Plant Cell Environ. 2008, 31, 658–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fletcher, A.L.; Sinclair, T.R.; Allen, L.H., Jr. Transpiration responses to vapor pressure deficit in well-watered “slow-wilting” and commercial soybean. Environ. Exp. Bot. 2007, 61, 145–151. [Google Scholar] [CrossRef]
- Shekoofa, A.; Choudhary, S. Maize. In Water-Conservation Traits to Increase Crop Yields in Water-Deficit Environments: Case Studies; Sinclair, T.R., Ed.; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Sinclair, T.R.; Pradhan, D.; Shekoofa, A. Update on Inheritance of Limited-Transpiration Trait in Peanut. J. Crop Improv. 2018, 32, 281–286. [Google Scholar] [CrossRef]
- Javot, H.; Maurel, C. The role of aquaporins in root water uptake. Ann. Bot. (Lond.) 2002, 90, 301–313. [Google Scholar] [CrossRef]
- Sadok, W.; Sinclair, T.R. Transpiration response of ‘slow-wilting’ and commercial soybean (Glycine max (L.) Merr.) genotypes to three aquaporin inhibitors. J. Exp. Bot. 2010, 61, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Kastori, R.; Petrovic, M.; Petrovic, N. Effect of excess lead, cadmium, copper, and zinc on water relations in sunflower. J. Plant Nutr. 1992, 15, 2427–2439. [Google Scholar] [CrossRef]
- Sagardoy, R.; Morales, F.; López-Millán, A.F.; Abadía, A.; Abadía, J. Effects of zinc toxicity on sugar beet (Beta vulgaris L.) plants grown in hydroponics. Plant Biol. 2009, 11, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Devi, J.; Sinclair, T.R.; Tliercio, E. Silver and zinc inhibitors influence transpiration rate and aquaporin transcript abundance in intact soybean plants. Environ. Exp. Bot. 2016, 122, 168–175. [Google Scholar] [CrossRef]
- Choudhary, S.; Sinclair, T.R.; Messina, C.D.; Cai, W.; Warner, D.; Cooper, M. Inhibitor screen for limited-transpiration trait among maize hybrids. Environ. Exp. Bot. 2015, 109, 161–167. [Google Scholar] [CrossRef]
- Gholipoor, M.; Choudhary, S.; Sinclair, T.R.; Messina, C.D.; Cooper, M. Transpiration response of maize hybrids to atmospheric vapor pressure deficit. J. Agron. Crop Sci. 2013, 199, 155–160. [Google Scholar] [CrossRef]
- Shekoofa, A.; Devi, M.J.; Sinclair, T.R.; Isleib, T.G. Divergence in drought-resistance traits among parents of recombinant peanut inbred lines. Crop Sci. 2013, 53, 2569–2576. [Google Scholar] [CrossRef]
- Choudhary, S.; Sinclair, T.R.; Prasad, P.V. Hydraulic conductance of intact plants of two contrasting sorghum lines, SC15 and SC1205. Funct. Plant Biol. 2013, 40, 730–738. [Google Scholar] [CrossRef]
- Shekoofa, A.; Sinclair, T.R.; Aninbon, C.; Holbrook, C.C.; Isleib, T.G.; Ozias-Akins, P.; Chu, Y. Expression of the limited transpiration trait under high vapor pressure deficit in peanut RIL populations: Runner and Virginia type. J. Agron. Crop Sci. 2017, 203, 295–300. [Google Scholar] [CrossRef]
- Nardini, A.; Salleo, S. Water stress-induced modifications of leaf hydraulic architecture in sunflower: Co-ordination with gas exchange. Environ. Exp. Bot. 2005, 422, 3093–3101. [Google Scholar] [CrossRef] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shekoofa, A.; Sinclair, T.R. Aquaporin Activity to Improve Crop Drought Tolerance. Cells 2018, 7, 123. https://doi.org/10.3390/cells7090123
Shekoofa A, Sinclair TR. Aquaporin Activity to Improve Crop Drought Tolerance. Cells. 2018; 7(9):123. https://doi.org/10.3390/cells7090123
Chicago/Turabian StyleShekoofa, Avat, and Thomas R. Sinclair. 2018. "Aquaporin Activity to Improve Crop Drought Tolerance" Cells 7, no. 9: 123. https://doi.org/10.3390/cells7090123
APA StyleShekoofa, A., & Sinclair, T. R. (2018). Aquaporin Activity to Improve Crop Drought Tolerance. Cells, 7(9), 123. https://doi.org/10.3390/cells7090123