Transient Receptor Potential Channel A1 (TRPA1) Regulates Sulfur Mustard-Induced Expression of Heat Shock 70 kDa Protein 6 (HSPA6) In Vitro


Abstract
:1. Introduction
2. Materials & Methods
2.1. Chemicals
2.2. Cell Culture
2.3. Sample Preparation
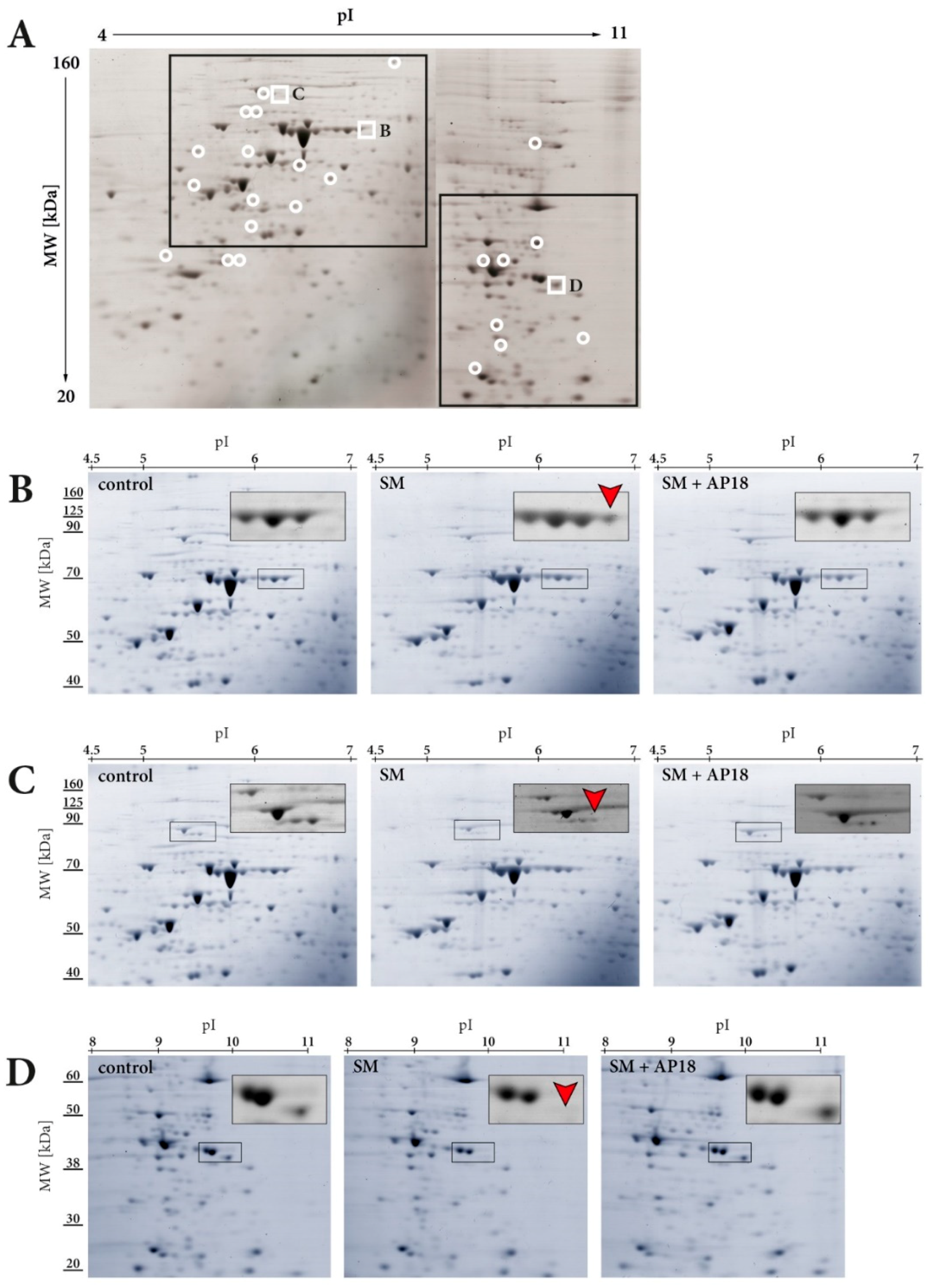
2.4. 2D Gel Electrophoresis and Image Analysis
2.5. MALDI-TOF MS(/MS) or NanoHPLC-ESI MS/MS Analysis
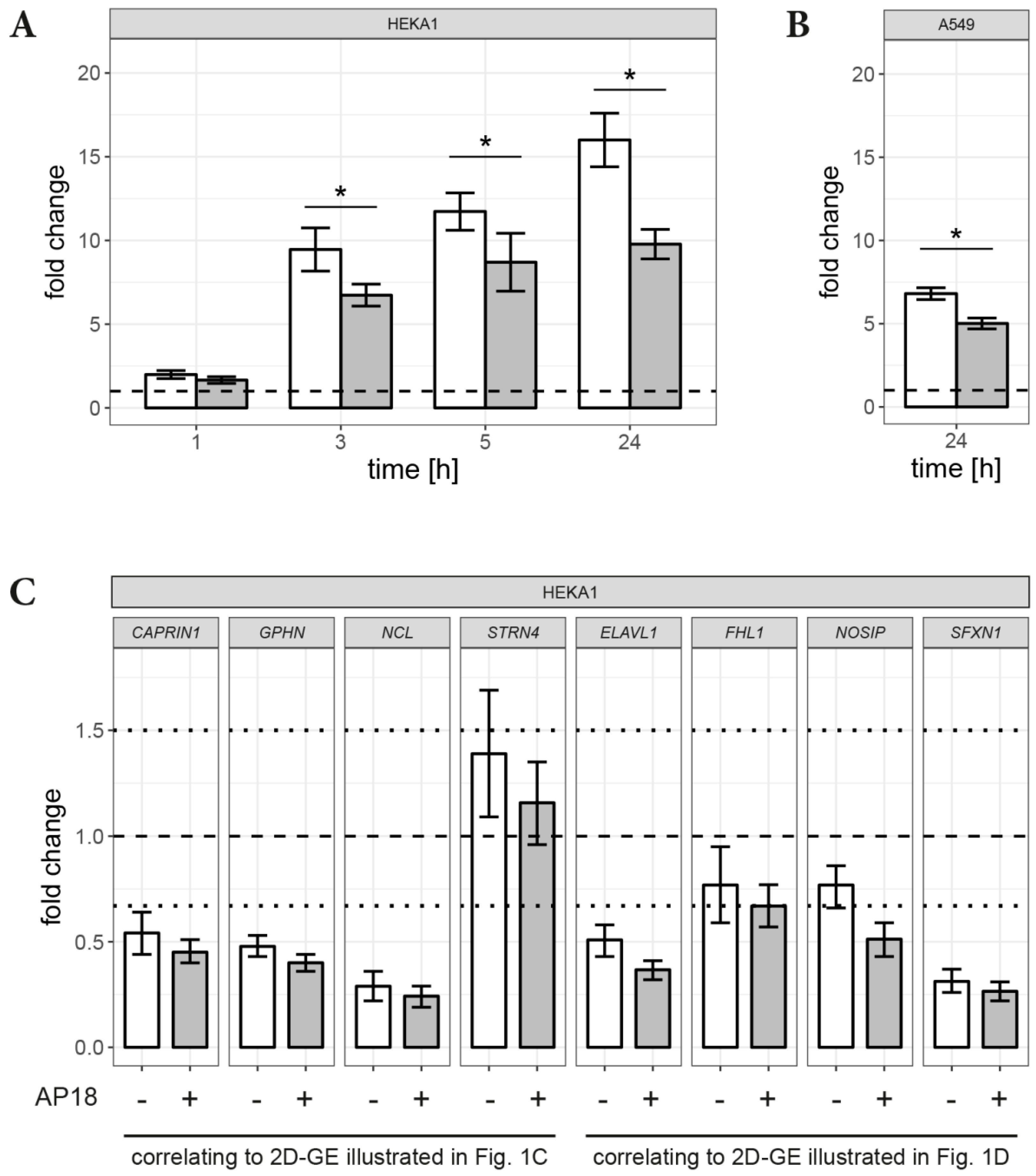
2.6. Real-Time qPCR
3. Results
3.1. 2D Gel Electrophoresis and Mass Spectrometry
3.2. RT-qPCR
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Steinritz, D.; Balszuweit, F.; Thiermann, H.; Kehe, K. Mustard: Pathophysiology and Therapeutic Approaches. In Chemical Warfare Toxicology; Worek, F., Jenner, J., Thiermann, H., Eds.; Royal Society of Chemistry: Cambridge, UK, 2016; pp. 120–156. [Google Scholar]
- Kehe, K.; Steinritz, D.; Balszuweit, F.; Thiermann, H. Long-Term Effects of the Chemical Warfare Agent Sulfur Mustard. In Chemical Warfare Toxicology; Worek, F., Jenner, J., Thiermann, H., Eds.; Royal Society of Chemistry: Cambridge, UK, 2016; pp. 179–190. [Google Scholar]
- Steinritz, D.; Thiermann, H. Sulfur Mustard. In Critical Care Toxicology: Diagnosis and Management of the Critically Poisoned Patient, 2nd ed.; Brent, J., Burkhart, K., Dargan, P.I., Hatten, B., Megarbane, B., Palmer, R., White, J., Eds.; Springer: Cham, Switzerland, 2017; pp. 2683–2712. [Google Scholar]
- Stenger, B.; Zehfuss, F.; Mückter, H.; Schmidt, A.; Balszuweit, F.; Schäfer, E.; Büch, T.; Gudermann, T.; Thiermann, H.; Steinritz, D. Activation of the chemosensing transient receptor potential channel A1 (TRPA1) by alkylating agents. Arch. Toxicol. 2015, 89, 1631–1643. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, I.S.; Delling, M.; Clapham, D.E. An introduction to TRP channels. Annu. Rev. Physiol. 2006, 68, 619–647. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Hackos, D.H. TRPA1 as a drug target—Promise and challenges. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2015, 388, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.J.M.; Balasuriya, D.; Jeggle, P.; Goetze, T.A.; McNaughton, P.A.; Reeh, P.W.; Edwardson, J.M. Direct evidence for functional TRPV1/TRPA1 heteromers. Pflug. Arch.-Eur. J. Physiol. 2014, 466, 2229–2241. [Google Scholar] [CrossRef] [PubMed]
- Madej, M.G.; Ziegler, C.M. Dawning of a new era in TRP channel structural biology by cryo-electron microscopy. Pflug. Arch.-Eur. J. Physiol. 2018, 470, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Macpherson, L.J.; Dubin, A.E.; Evans, M.J.; Marr, F.; Schultz, P.G.; Cravatt, B.F.; Patapoutian, A. Noxious compounds activate TRPA1 ion channels through covalent modification of cysteines. Nature 2007, 445, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Bessac, B.F.; Jordt, S.-E. Sensory detection and responses to toxic gases: Mechanisms, health effects, and countermeasures. Proc. Am. Thorac. Soc. 2010, 7, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Jordt, S.-E.; Bautista, D.M.; Chuang, H.-H.; McKemy, D.D.; Zygmunt, P.M.; Högestätt, E.D.; Meng, I.D.; Julius, D. Mustard oils and cannabinoids excite sensory nerve fibres through the TRP channel ANKTM1. Nature 2004, 427, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Achanta, S.; Jordt, S.-E. TRPA1: Acrolein meets its target. Toxicol. Appl. Pharmacol. 2017, 324, 45–50. [Google Scholar] [CrossRef] [PubMed]
- De La Roche, J.; Eberhardt, M.J.; Klinger, A.B.; Stanslowsky, N.; Wegner, F.; Koppert, W.; Reeh, P.W.; Lampert, A.; Fischer, M.J.M.; Leffler, A. The molecular basis for species-specific activation of human TRPA1 protein by protons involves poorly conserved residues within transmembrane domains 5 and 6. J. Biol. Chem. 2013, 288, 20280–20292. [Google Scholar] [CrossRef] [PubMed]
- Bessac, B.F.; Sivula, M.; Von Hehn, C.A.; Escalera, J.; Cohn, L.; Jordt, S.-E. TRPA1 is a major oxidant sensor in murine airway sensory neurons. J. Clin. Investig. 2008, 118, 1899–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hox, V.; Vanoirbeek, J.A.; Alpizar, Y.A.; Voedisch, S.; Callebaut, I.; Bobic, S.; Sharify, A.; De Vooght, V.; Van Gerven, L.; Devos, F.; et al. Crucial role of transient receptor potential ankyrin 1 and mast cells in induction of nonallergic airway hyperreactivity in mice. Am. J. Respir. Crit. Care Med. 2013, 187, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Arenas, O.M.; Zaharieva, E.E.; Para, A.; Vásquez-Doorman, C.; Petersen, C.P.; Gallio, M. Activation of planarian TRPA1 by reactive oxygen species reveals a conserved mechanism for animal nociception. Nat. Neurosci. 2017, 20, 1686–1693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, A.-H.; Liu, M.-H.; Ko, H.-K.; Perng, D.-W.; Lee, T.-S.; Kou, Y.R. Lung Epithelial TRPA1 Transduces the Extracellular ROS into Transcriptional Regulation of Lung Inflammation Induced by Cigarette Smoke: The Role of Influxed Ca2+. Mediat. Inflamm. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, G.; Hoffmeister, C.; Rossato, M.F.; Oliveira, S.M.; Silva, M.A.; Silva, C.R.; Fusi, C.; Tonello, R.; Minocci, D.; Guerra, G.P.; et al. TRPA1 receptor stimulation by hydrogen peroxide is critical to trigger hyperalgesia and inflammation in a model of acute gout. Free Radic. Biol. Med. 2014, 72, 200–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenger, B.; Popp, T.; John, H.; Siegert, M.; Tsoutsoulopoulos, A.; Schmidt, A.; Mückter, H.; Gudermann, T.; Thiermann, H.; Steinritz, D. N-Acetyl-L-cysteine inhibits sulfur mustard-induced and TRPA1-dependent calcium influx. Arch. Toxicol. 2017, 91, 2179–2189. [Google Scholar] [CrossRef] [PubMed]
- Macpherson, L.J.; Xiao, B.; Kwan, K.Y.; Petrus, M.J.; Dubin, A.E.; Hwang, S.; Cravatt, B.; Corey, D.P.; Patapoutian, A. An ion channel essential for sensing chemical damage. J. Neurosci. 2007, 27, 11412–11415. [Google Scholar] [CrossRef] [PubMed]
- Achanta, S.; Chintagari, N.R.; Brackmann, M.; Balakrishna, S.; Jordt, S.-E. TRPA1 and CGRP antagonists counteract vesicant-induced skin injury and inflammation. Toxicol. Lett. 2018, 293, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Huang, C.; Zhong, S.; Wortley, M.A.; Luo, Y.; Luo, W.; Xie, Y.; Lai, K.; Zhong, N. Cigarette smoke extract (CSE) induces transient receptor potential ankyrin 1(TRPA1) expression via activation of HIF1αin A549 cells. Free Radic. Biol. Med. 2016, 99, 498–507. [Google Scholar] [CrossRef] [PubMed]
- Büch, T.R.H.; Schäfer, E.A.M.; Demmel, M.-T.; Boekhoff, I.; Thiermann, H.; Gudermann, T.; Steinritz, D.; Schmidt, A. Functional expression of the transient receptor potential channel TRPA1, a sensor for toxic lung inhalants, in pulmonary epithelial cells. Chem.-Biol. Interact. 2013, 206, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, I.; Gomes, P.; Aranake, S.; Shetty, M.; Karnik, P.; Damle, M.; Kuruganti, S.; Thorat, S.; Khairatkar-Joshi, N. Expression of functional TRPA1 receptor on human lung fibroblast and epithelial cells. J. Recept. Signal Transduct. 2011, 31, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Qiagen. Data Analysis Center. 2018. Available online: https://www.qiagen.com/de/shop/genes-and-pathways/data-analysis-center-overview-page/?akamai-feo=off (accessed on 10 July 2018).
- Protein Isoelectric Point Calculator. 2018. Available online: http://isoelectric.org/index.html (accessed on 23 August 2018).
- Protein Isoelectric Point Calculator. 2018. Available online: http://www.bioinformatics.org/sms2/index.html (accessed on 23 August 2018).
- Kehe, K.; Balszuweit, F.; Steinritz, D.; Thiermann, H. Molecular toxicology of sulfur mustard-induced cutaneous inflammation and blistering. Toxicology 2009, 263, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Fu, D.; Calvo, J.A.; Samson, L.D. Balancing repair and tolerance of DNA damage caused by alkylating agents. Nat. Rev. Cancer 2012, 12, 104–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dacre, J.C.; Goldman, M. Toxicology and pharmacology of the chemical warfare agent sulfur mustard. Pharmacol. Rev. 1996, 48, 289–326. [Google Scholar] [PubMed]
- Deans, A.J.; West, S.C. DNA interstrand crosslink repair and cancer. Nat. Rev. Cancer 2011, 11, 467–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Li, L. DNA crosslinking damage and cancer—A tale of friend and foe. Transl. Cancer Res. 2013, 2, 144–154. [Google Scholar] [PubMed]
- Kondo, N.; Takahashi, A.; Ono, K.; Ohnishi, T. DNA damage induced by alkylating agents and repair pathways. J. Nucleic Acids 2010, 2010. [Google Scholar] [CrossRef] [PubMed]
- Steinritz, D.; Elischer, A.; Balszuweit, F.; Gonder, S.; Heinrich, A.; Bloch, W.; Thiermann, H.; Kehe, K. Sulphur mustard induces time- and concentration-dependent regulation of NO-synthesizing enzymes. Toxico. Lett. 2009, 188, 263–269. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, H.C.; Orlicky, D.J.; Hendry-Hofer, T.B.; Loader, J.E.; Day, B.J.; White, C.W. Role of reactive oxygen and nitrogen species in olfactory epithelial injury by the sulfur mustard analogue 2-chloroethyl ethyl sulfide. Am. J. Respir. Cell Mol. Biol. 2011, 45, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Brent, J.; Burkhart, K.; Dargan, P.I.; Hatten, B.; Megarbane, B.; Palmer, R.; White, J. Critical Care Toxicology. Diagnosis and Management of the Critically Poisoned Patient, 2nd ed.; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Jain, A.K.; Tewari-Singh, N.; Gu, M.; Inturi, S.; White, C.W.; Agarwal, R. Sulfur mustard analog, 2-chloroethyl ethyl sulfide-induced skin injury involves DNA damage and induction of inflammatory mediators, in part via oxidative stress, in SKH-1 hairless mouse skin. Toxicol. Lett. 2011, 205, 293–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tewari-Singh, N.; Jain, A.K.; Inturi, S.; Agarwal, C.; White, C.W.; Agarwal, R. Silibinin attenuates sulfur mustard analog-induced skin injury by targeting multiple pathways connecting oxidative stress and inflammation. PLoS ONE 2012, 7, e46149. [Google Scholar] [CrossRef] [PubMed]
- Steinritz, D.; Weber, J.; Balszuweit, F.; Thiermann, H.; Schmidt, A. Sulfur mustard induced nuclear translocation of glyceraldehyde-3-phosphate-dehydrogenase (GAPDH). Chem.-Biol. Interact. 2013, 206, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Steinritz, D.; Zehfuß, F.; Stenger, B.; Schmidt, A.; Popp, T.; Kehe, K.; Mückter, H.; Thiermann, H.; Gudermann, T. Zinc chloride-induced TRPA1 activation does not contribute to toxicity in vitro. Toxicol. Lett. 2018, 293, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, E.A.M.; Stohr, S.; Meister, M.; Aigner, A.; Gudermann, T.; Buech, T.R.H. Stimulation of the chemosensory TRPA1 cation channel by volatile toxic substances promotes cell survival of small cell lung cancer cells. Biochem. Pharmacol. 2013, 85, 426–438. [Google Scholar] [CrossRef] [PubMed]
- Everley, P.A.; Dillman, J.F. A large-scale quantitative proteomic approach to identifying sulfur mustard-induced protein phosphorylation cascades. Chem. Res. Toxicol. 2010, 23, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.V.; Choi, Y.W.; Kiser, R.C.; Babin, M.C.; Casillas, R.P.; Schlager, J.J.; Sabourin, C.L.K. Microarray analysis of gene expression in murine skin exposed to sulfur mustard. J. Biochem. Mol. Toxicol. 2004, 18, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Petrus, M.; Peier, A.M.; Bandell, M.; Hwang, S.W.; Huynh, T.; Olney, N.; Jegla, T.; Patapoutian, A. A role of TRPA1 in mechanical hyperalgesia is revealed by pharmacological inhibition. Mol. Pain 2007, 3, 40. [Google Scholar] [CrossRef] [PubMed]
- Solomon, S.; Xu, Y.; Wang, B.; David, M.D.; Schubert, P.; Kennedy, D.; Schrader, J.W. Distinct structural features of caprin-1 mediate its interaction with G3BP-1 and its induction of phosphorylation of eukaryotic translation initiation factor 2α, entry to cytoplasmic stress granules, and selective interaction with a subset of mRNAs. Mol. Cell. Biol. 2007, 27, 2324–2342. [Google Scholar] [CrossRef] [PubMed]
- Castets, F.; Rakitina, T.; Gaillard, S.; Moqrich, A.; Mattei, M.G.; Monneron, A. Zinedin, SG2NA, and striatin are calmodulin-binding, WD repeat proteins principally expressed in the brain. J. Biol. Chem. 2000, 275, 19970–19977. [Google Scholar] [CrossRef] [PubMed]
- Losfeld, M.-E.; Khoury, D.E.; Mariot, P.; Carpentier, M.; Krust, B.; Briand, J.-P.; Mazurier, J.; Hovanessian, A.G.; Legrand, D. The cell surface expressed nucleolin is a glycoprotein that triggers calcium entry into mammalian cells. Exp. Cell Res. 2009, 315, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Cong, R.; Das, S.; Douet, J.; Wong, J.; Buschbeck, M.; Mongelard, F.; Bouvet, P. macroH2A1 histone variant represses rDNA transcription. Nucleic Acids Res. 2014, 42, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Reiss, J.; Lenz, U.; Aquaviva-Bourdain, C.; Joriot-Chekaf, S.; Mention-Mulliez, K.; Holder-Espinasse, M. A GPHN point mutation leading to molybdenum cofactor deficiency. Clin. Genet. 2011, 80, 598–599. [Google Scholar] [CrossRef] [PubMed]
- Chu, P.-H.; Chen, J. The novel roles of four and a half LIM proteins 1 and 2 in the cardiovascular system. Chang Gung Med. J. 2011, 34, 127–134. [Google Scholar] [PubMed]
- Doller, A.; Akool, E.-S.; Huwiler, A.; Müller, R.; Radeke, H.H.; Pfeilschifter, J.; Eberhardt, W. Posttranslational modification of the AU-rich element binding protein HuR by protein kinase Cdelta elicits angiotensin II-induced stabilization and nuclear export of cyclooxygenase 2 mRNA. Mol. Cell. Biol. 2008, 28, 2608–2625. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.; Maurer, F.; Nagamine, Y. Stabilization of Urokinase and Urokinase Receptor mRNAs by HuR Is Linked to Its Cytoplasmic Accumulation Induced by Activated Mitogen-Activated Protein Kinase-Activated Protein Kinase 2. Mol. Cell. Biol. 2003, 23, 7177–7188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshikumi, Y.; Mashima, H.; Ueda, N.; Ohno, H.; Suzuki, J.; Tanaka, S.; Hayashi, M.; Sekine, N.; Ohnishi, H.; Yasuda, H.; et al. Roles of CTPL/Sfxn3 and Sfxn family members in pancreatic islet. J. Cell. Biochem. 2005, 95, 1157–1168. [Google Scholar] [CrossRef] [PubMed]
- Xi, D.; He, Y.; Sun, Y.; Gou, X.; Yang, S.; Mao, H.; Deng, W. Molecular cloning, sequence identification and tissue expression profile of three novel genes Sfxn1, Snai2 and Cno from Black-boned sheep (Ovis aries). Mol. Biol. Rep. 2011, 38, 1883–1887. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, J.; Schleicher, M.; Tappe, A.; Schilling, K.; Kuner, T.; Kusumawidijaja, G.; Müller-Esterl, W.; Oess, S.; Kuner, R. Nitric oxide synthase (NOS)-interacting protein interacts with neuronal NOS and regulates its distribution and activity. J. Neurosci. 2004, 24, 10454–10465. [Google Scholar] [CrossRef] [PubMed]
- Black, A.T.; Hayden, P.J.; Casillas, R.P.; Heck, D.E.; Gerecke, D.R.; Sinko, P.J.; Laskin, D.L.; Laskin, J.D. Regulation of Hsp27 and Hsp70 expression in human and mouse skin construct models by caveolae following exposure to the model sulfur mustard vesicant, 2-chloroethyl ethyl sulfide. Toxicol. Appl. Pharmacol. 2011, 253, 112–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liberek, K.; Lewandowska, A.; Zietkiewicz, S. Chaperones in control of protein disaggregation. EMBO J. 2008, 27, 328–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maytin, E.V. Heat Shock Proteins and Molecular Chaperones: Implication for Adaptive Responses in the Skin. J. Investig. Dermatol. 1995, 104, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Trautinger, F. Heat shock proteins in the photobiology of human skin. J. Photochem. Photobiol. B Biol. 2001, 63, 70–77. [Google Scholar] [CrossRef]
- Macario, A.J.L. Molecular chaperones: Multiple functions, pathologies, and potential applications. Front. Biosci. 2007, 12, 2588–2600. [Google Scholar] [CrossRef] [PubMed]
- Madamanchi, N.R.; Li, S.; Patterson, C.; Runge, M.S. Reactive Oxygen Species Regulate Heat-Shock Protein 70 via the JAK/STAT Pathway. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 321–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brookes, P.S.; Yoon, Y.; Robotham, J.L.; Anders, M.W.; Sheu, S.-S. Calcium, ATP, and ROS: A mitochondrial love-hate triangle. Am. J. Physiol.-Cell Physiol. 2004, 287, C817–C833. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.-L.; Yoshioka, T. Role of TRP channels in the induction of heat shock proteins (Hsps) by heating skin. Biophysics 2015, 11, 25–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gould, N.S.; White, C.W.; Day, B.J. A role for mitochondrial oxidative stress in sulfur mustard analog 2-chloroethyl ethyl sulfide-induced lung cell injury and antioxidant protection. J. Pharmacol. Exp. Ther. 2009, 328, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Ray, R.; Legere, R.H.; Majerus, B.J.; Petrali, J.P. Sulfur mustard-induced increase in intracellular free calcium level and arachidonic acid release from cell membrane. Toxicol. Appl. Pharmacol. 1995, 131, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Sawale, S.D.; Ambhore, P.D.; Pawar, P.P.; Pathak, U.; Deb, U.; Satpute, R.M. Ameliorating effect of S-2(ω-aminoalkylamino) alkylaryl sulfide (DRDE-07) on sulfur mustard analogue, 2-chloroethyl ethyl sulfide-induced oxidative stress and inflammation. Toxicol. Mech. Methods 2013, 23, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Mei, Y.; Wang, T.; Liu, F.; Jiang, N.; Zhou, W.; Zhang, Y. Early oxidative stress, DNA damage and inflammation resulting from subcutaneous injection of sulfur mustard into mice. Environ. Toxicol. Pharmacol. 2017, 55, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Steinritz, D.; Schmidt, A.; Simons, T.; Ibrahim, M.; Morguet, C.; Balszuweit, F.; Thiermann, H.; Kehe, K.; Bloch, W.; Bölck, B. Chlorambucil (nitrogen mustard) induced impairment of early vascular endothelial cell migration—Effects of α-linolenic acid and N-acetylcysteine. Chem.-Biol. Interact. 2014, 219, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, D.S.; Simbulan-Rosenthal, C.M.; Iyer, S.; Spoonde, A.; Smith, W.; Ray, R.; Smulson, M.E. Sulfur mustard induces markers of terminal differentiation and apoptosis in keratinocytes via a Ca2+-calmodulin and caspase-dependent pathway. J. Investig. Dermatol. 1998, 111, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Simbulan-Rosenthal, C.M.; Ray, R.; Benton, B.; Soeda, E.; Daher, A.; Anderson, D.; Smith, W.J.; Rosenthal, D.S. Calmodulin mediates sulfur mustard toxicity in human keratinocytes. Toxicology 2006, 227, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, M.G.; Dorandeu, F.M.; McCaffery, M.; Lundy, P.M.; Sawyer, T.W. Modification of cytosolic free calcium concentrations in human keratinocytes after sulfur mustard exposure. Toxicol. In Vitro 1998, 12, 365–372. [Google Scholar] [CrossRef]
- Mosser, D.D.; Caron, A.W.; Bourget, L.; Meriin, A.B.; Sherman, M.Y.; Morimoto, R.I.; Massie, B. The Chaperone Function of hsp70 Is Required for Protection against Stress-Induced Apoptosis. Mol. Cell. Biol. 2000, 20, 7146–7159. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Protein Name | UniProt ID | MW[kDa] | pI[Calculated] | Number of Identified Peptides | MS/MS Score | |
|---|---|---|---|---|---|---|---|
| Figure 1C | CAPRIN1 | Caprin-1 | Q14444 | 78.5 | 5.0 | 17 | 864 |
| STRN4 | Striatin 4 | Q9NRL3 | 81.3 | 5.1 | 13 | 787 | |
| NCL | Nucleolin | P19338 | 76.6 | 4.5 | 10 | 443 | |
| GPHN | Gephyrin | Q9NQX3 | 80.4 | 5.1 | 7 | 372 | |
| Figure 1D | SFXN1 | Sideroflexin 1 | Q9H9B4 | 35.9 | 9.4 | 10 | 689 |
| FHL1 | Four and a half LIM domain protein 1 | Q13642 | 38.0 | 9.2 | 9 | 592 | |
| NOSIP | Nitric oxide synthase interacting protein | Q9Y314 | 33.7 | 9.1 | 11 | 554 | |
| ELAVL1 | ELAV like protein 1 | Q15717 | 36.2 | 9.6 | 8 | 445 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lüling, R.; John, H.; Gudermann, T.; Thiermann, H.; Mückter, H.; Popp, T.; Steinritz, D. Transient Receptor Potential Channel A1 (TRPA1) Regulates Sulfur Mustard-Induced Expression of Heat Shock 70 kDa Protein 6 (HSPA6) In Vitro. Cells 2018, 7, 126. https://doi.org/10.3390/cells7090126
Lüling R, John H, Gudermann T, Thiermann H, Mückter H, Popp T, Steinritz D. Transient Receptor Potential Channel A1 (TRPA1) Regulates Sulfur Mustard-Induced Expression of Heat Shock 70 kDa Protein 6 (HSPA6) In Vitro. Cells. 2018; 7(9):126. https://doi.org/10.3390/cells7090126
Chicago/Turabian StyleLüling, Robin, Harald John, Thomas Gudermann, Horst Thiermann, Harald Mückter, Tanja Popp, and Dirk Steinritz. 2018. "Transient Receptor Potential Channel A1 (TRPA1) Regulates Sulfur Mustard-Induced Expression of Heat Shock 70 kDa Protein 6 (HSPA6) In Vitro" Cells 7, no. 9: 126. https://doi.org/10.3390/cells7090126
APA StyleLüling, R., John, H., Gudermann, T., Thiermann, H., Mückter, H., Popp, T., & Steinritz, D. (2018). Transient Receptor Potential Channel A1 (TRPA1) Regulates Sulfur Mustard-Induced Expression of Heat Shock 70 kDa Protein 6 (HSPA6) In Vitro. Cells, 7(9), 126. https://doi.org/10.3390/cells7090126