Telomeres in Plants and Humans: Not So Different, Not So Similar


Abstract
:1. Introduction
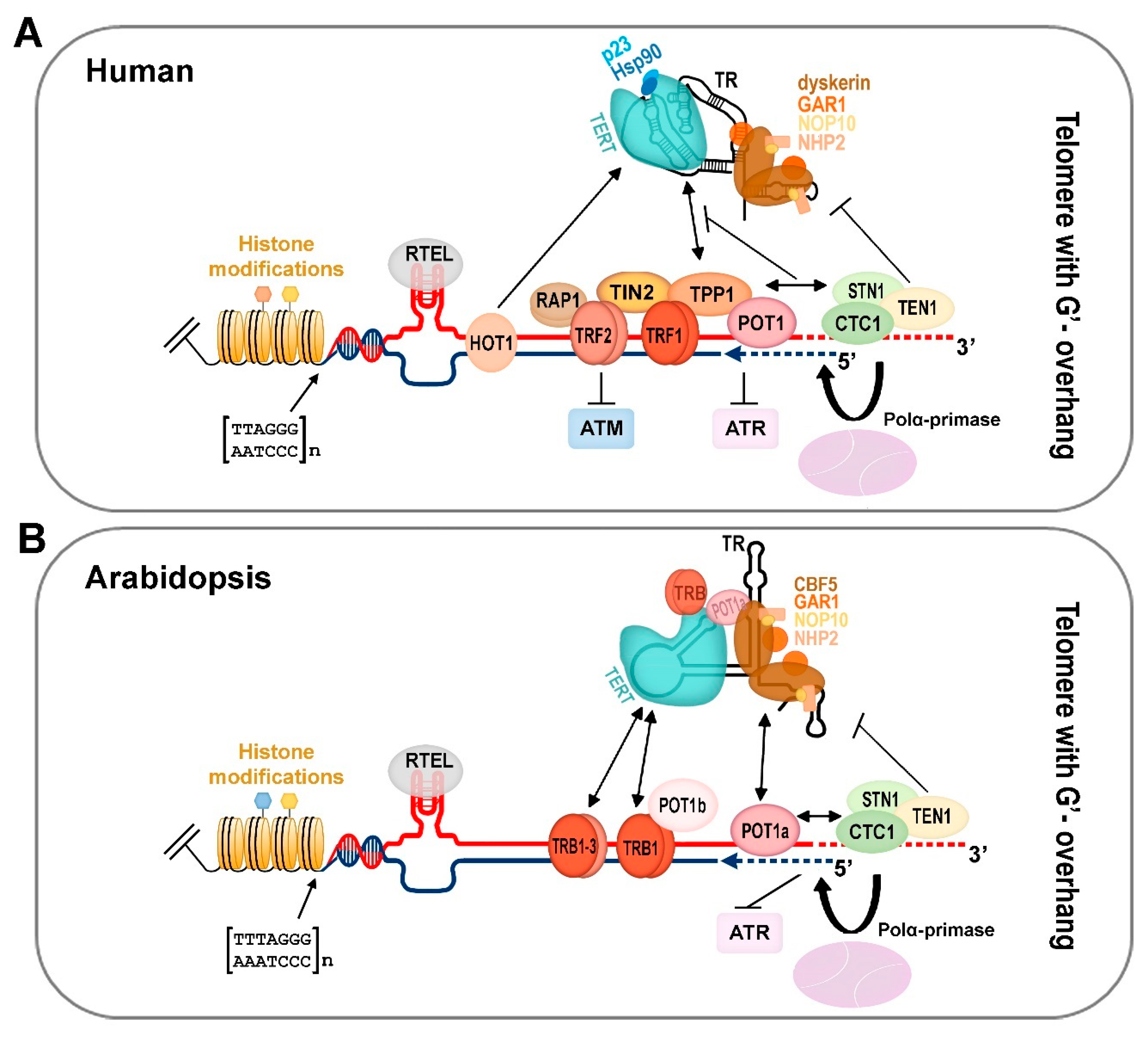
2. Telomerase Core Components
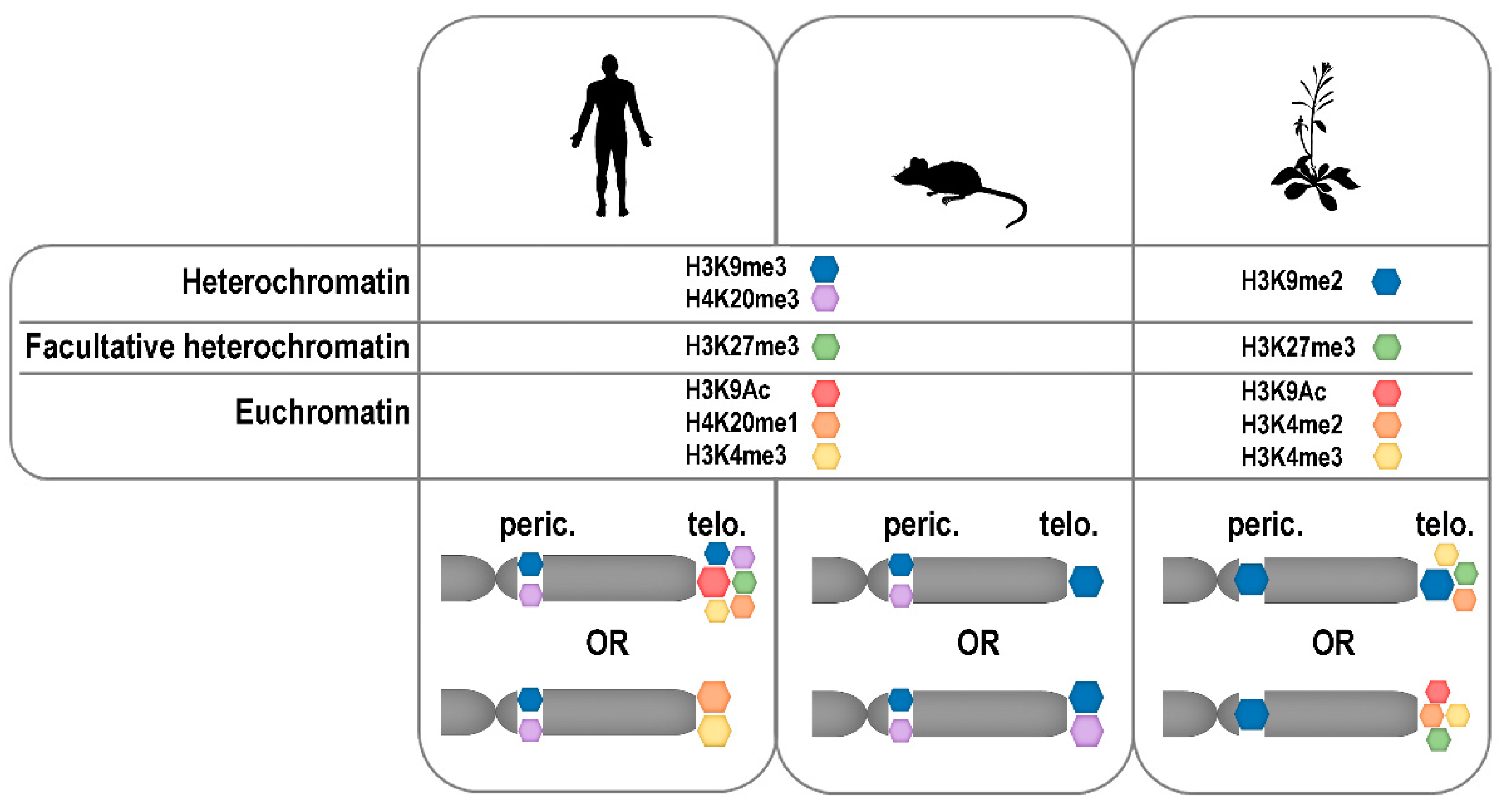
3. Telomere Chromatin Composition
4. Telomere Epigenetics
5. Telomere 3′-Overhangs, Blunt Ends, and Loops
6. Cellular Aging and the Immortal DNA Strand Hypothesis
7. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Olovnikov, A.M. Principle of marginotomy in template synthesis of polynucleotides. Dokl. Akad. Nauk. SSSR 1971, 201, 1496–1499. [Google Scholar]
- Muller, H.J. The remaking of chromosomes. Collect. Net. 1938, 13, 181–195. [Google Scholar]
- McClintock, B. The fusion of broken chromosome ends of sister half-chromatids following chromatid breakage at meiotic anaphases. Mo. Agric. Exp. Stn. Res. Bull. 1938, 290, 1–48. [Google Scholar]
- McClintock, B. The stability of broken ends of chromosomes in Zea mays. Genetics 1941, 26, 234–282. [Google Scholar]
- Von Zglinicki, T. Oxidative stress shortens telomeres. Trends Biochem. Sci. 2002, 27, 339–344. [Google Scholar] [CrossRef]
- Hoelzl, F.; Cornils, J.S.; Smith, S.; Moodley, Y.; Ruf, T. Telomere dynamics in free-living edible dormice (Glis glis): The impact of hibernation and food supply. J. Exp. Biol. 2016, 219, 2469–2474. [Google Scholar] [CrossRef]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [Green Version]
- Simons, M.J.P. Questioning causal involvement of telomeres in aging. Ageing Res. Rev. 2015, 24, 191–196. [Google Scholar] [CrossRef]
- Steenstrup, T.; Kark, J.D.; Verhulst, S.; Thinggaard, M.; Hjelmborg, J.V.B.; Dalgard, C.; Kyvik, K.O.; Christiansen, L.; Mangino, M.; Spector, T.D.; et al. Telomeres and the natural lifespan limit in humans. Aging US 2017, 9, 1130–1142. [Google Scholar] [CrossRef] [Green Version]
- Factor-Litvak, P.; Susser, E.; Kezios, K.; McKeague, I.; Kark, J.D.; Hoffman, M.; Kimura, M.; Wapner, R.; Aviv, A. Leukocyte Telomere Length in Newborns: Implications for the Role of Telomeres in Human Disease. Pediatrics 2016, 137, e20153927. [Google Scholar] [CrossRef]
- Robin, J.D.; Ludlow, A.T.; Batten, K.; Magdinier, F.; Stadler, G.; Wagner, K.R.; Shay, J.W.; Wright, W.E. Telomere position effect: Regulation of gene expression with progressive telomere shortening over long distances. Genes Dev. 2014, 28, 2464–2476. [Google Scholar] [CrossRef]
- Victorelli, S.; Passos, J.F. Telomeres and Cell Senescence—Size Matters Not. Ebiomedicine 2017, 21, 14–20. [Google Scholar] [CrossRef]
- Abdallah, P.; Luciano, P.; Runge, K.W.; Lisby, M.; Geli, V.; Gilson, E.; Teixeira, M.T. A two-step model for senescence triggered by a single critically short telomere. Nat. Cell Biol. 2009, 11, 988. [Google Scholar] [CrossRef]
- Hemann, M.T.; Strong, M.A.; Hao, L.Y.; Greider, C.W. The shortest telomere, not average telomere length, is critical for cell viability and chromosome stability. Cell 2001, 107, 67–77. [Google Scholar] [CrossRef]
- Kaul, Z.; Cesare, A.J.; Huschtscha, L.I.; Neumann, A.A.; Reddel, R.R. Five dysfunctional telomeres predict onset of senescence in human cells. Embo Rep. 2012, 13, 52–59. [Google Scholar] [CrossRef]
- Watson, J.M.; Riha, K. Telomeres, aging, and plants: From weeds to Methuselah—A mini-review. Gerontology 2011, 57, 129–136. [Google Scholar] [CrossRef]
- Barsov, E.V. Telomerase and primary T cells: Biology and immortalization for adoptive immunotherapy. Immunotherapy 2011, 3, 407–421. [Google Scholar] [CrossRef]
- Shalaby, T.; Hiyama, E.; Grotzer, M.A. Telomere Maintenance as Therapeutic Target in Embryonal Tumours. Anti-Cancer Agents Med. Chem. 2010, 10, 196–212. [Google Scholar] [CrossRef]
- Fajkus, J.; Kovarik, A.; Kralovics, R. Telomerase activity in plant cells. Febs Lett. 1996, 391, 307–309. [Google Scholar] [CrossRef] [Green Version]
- Fajkus, J.; Fulneckova, J.; Hulanova, M.; Berkova, K.; Riha, K.; Matyasek, R. Plant cells express telomerase activity upon transfer to callus culture, without extensively changing telomere lengths. Mol. Gen. Genet. 1998, 260, 470–474. [Google Scholar] [CrossRef]
- Fitzgerald, M.S.; McKnight, T.D.; Shippen, D.E. Characterization and developmental patterns of telomerase expression in plants. Proc. Natl. Acad. Sci. USA 1996, 93, 14422–14427. [Google Scholar] [CrossRef] [Green Version]
- Fajkus, J.; Sykorova, E.; Leitch, A.R. Telomeres in evolution and evolution of telomeres. Chromosome Res. 2005, 13, 469–479. [Google Scholar] [CrossRef]
- Louis, E.J. Are Drosophila telomeres an exception or the rule? Genome Biol. 2002, 3. [Google Scholar] [CrossRef]
- Fajkus, P.; Peska, V.; Sitova, Z.; Fulneckova, J.; Dvorackova, M.; Gogela, R.; Sykorova, E.; Hapala, J.; Fajkus, J. Allium telomeres unmasked: The unusual telomeric sequence (CTCGGTTATGGG)(n) is synthesized by telomerase. Plant J. 2016, 85, 337–347. [Google Scholar] [CrossRef]
- Peska, V.; Fajkus, P.; Fojtova, M.; Dvorackova, M.; Hapala, J.; Dvoracek, V.; Polanska, P.; Leitch, A.R.; Sykorova, E.; Fajkus, J. Characterisation of an unusual telomere motif (TTTTTTAGGG)n in the plant Cestrum elegans (Solanaceae), a species with a large genome. Plant J. 2015, 82, 644–654. [Google Scholar] [CrossRef]
- Peska, V.; Sitova, Z.; Fajkus, P.; Fajkus, J. BAL31-NGS approach for identification of telomeres de novo in large genomes. Methods 2017, 114, 16–27. [Google Scholar] [CrossRef]
- Tran, T.D.; Cao, H.X.; Jovtchev, G.; Neumann, P.; Novak, P.; Fojtova, M.; Vu, G.T.H.; Macas, J.; Fajkus, J.; Schubert, I.; et al. Centromere and telomere sequence alterations reflect the rapid genome evolution within the carnivorous plant genus Genlisea. Plant J. 2015, 84, 1087–1099. [Google Scholar] [CrossRef] [Green Version]
- Wright, W.E.; Piatyszek, M.A.; Rainey, W.E.; Byrd, W.; Shay, J.W. Telomerase activity in human germline and embryonic tissues and cells. Dev. Genet. 1996, 18, 173–179. [Google Scholar] [CrossRef]
- Ramirez, R.D.; Wright, W.E.; Shay, J.W.; Taylor, R.S. Telomerase activity concentrates in the mitotically active segments of human hair follicles. J. Investig. Dermatol. 1997, 108, 113–117. [Google Scholar] [CrossRef]
- Hiyama, E.; Hiyama, K.; Yokoyama, T.; Shay, J.W. Immunohistochemical detection of telomerase (hTERT) protein in human cancer tissues and a subset of cells in normal tissues. Neoplasia 2001, 3, 17–26. [Google Scholar] [CrossRef]
- Hiyama, E.; Hiyama, K. Telomere and telomerase in stem cells. Br. J. Cancer 2007, 96, 1020–1024. [Google Scholar] [CrossRef] [Green Version]
- Hiyama, K.; Hirai, Y.; Kyoizumi, S.; Akiyama, M.; Hiyama, E.; Piatyszek, M.A.; Shay, J.W.; Ishioka, S.; Yamakido, M. Activation of Telomerase in Human-Lymphocytes and Hematopoietic Progenitor Cells. J. Immunol. 1995, 155, 3711–3715. [Google Scholar]
- Yui, J.; Chiu, C.P.; Lansdorp, P.M. Telomerase activity in candidate stem cells from fetal liver and adult bone marrow. Blood 1998, 91, 3255–3262. [Google Scholar]
- Ito, H.; Kyo, S.; Kanaya, T.; Takakura, M.; Inoue, M.; Namiki, M. Expression of human telomerase subunits and correlation with telomerase activity in urothelial cancer. Clin. Cancer Res. 1998, 4, 1603–1608. [Google Scholar]
- Kyo, S.; Takakura, M.; Kohama, T.; Inoue, M. Telomerase activity in human endometrium. Cancer Res. 1997, 57, 610–614. [Google Scholar]
- Jureckova, J.F.; Sykorova, E.; Hafidh, S.; Honys, D.; Fajkus, J.; Fojtova, M. Tissue-specific expression of telomerase reverse transcriptase gene variants in Nicotiana tabacum. Planta 2017, 245, 549–561. [Google Scholar] [CrossRef]
- Ogrocka, A.; Sykorova, E.; Fajkus, J.; Fojtova, M. Developmental silencing of the AtTERT gene is associated with increased H3K27me3 loading and maintenance of its euchromatic environment. J. Exp. Bot. 2012, 63, 4233–4241. [Google Scholar] [CrossRef] [Green Version]
- Riha, K.; Fajkus, J.; Siroky, J.; Vyskot, B. Developmental control of telomere lengths and telomerase activity in plants. Plant Cell 1998, 10, 1691–1698. [Google Scholar] [CrossRef]
- Zachova, D.; Fojtova, M.; Dvorackova, M.; Mozgova, I.; Lermontova, I.; Peska, V.; Schubert, I.; Fajkus, J.; Sykorova, E. Structure-function relationships during transgenic telomerase expression in Arabidopsis. Physiol. Plant. 2013, 149, 114–126. [Google Scholar] [CrossRef]
- Winter, D.; Vinegar, B.; Nahal, H.; Ammar, R.; Wilson, G.V.; Provart, N.J. An “Electronic Fluorescent Pictograph” Browser for Exploring and Analyzing Large-Scale Biological Data Sets. PLoS ONE 2007, 2. [Google Scholar] [CrossRef]
- Greider, C.W.; Blackburn, E.H. Identification of a Specific Telomere Terminal Transferase-Activity in Tetrahymena Extracts. Cell 1985, 43, 405–413. [Google Scholar] [CrossRef]
- Greider, C.W.; Blackburn, E.H. A Telomeric Sequence in the Rna of Tetrahymena Telomerase Required for Telomere Repeat Synthesis. Nature 1989, 337, 331–337. [Google Scholar] [CrossRef]
- Chan, H.; Wang, Y.Q.; Feigon, J. Progress in Human and Tetrahymena Telomerase Structure Determination. Annu. Rev. Biophys. 2017, 46, 199–225. [Google Scholar] [CrossRef]
- Nguyen, T.H.D.; Tam, J.; Wu, R.A.; Greber, B.J.; Toso, D.; Nogales, E.; Collins, K. Cryo-EM structure of substrate-bound human telomerase holoenzyme. Nature 2018, 557, 190. [Google Scholar] [CrossRef]
- Lermontova, I.; Schubert, V.; Bornke, F.; Macas, J.; Schubert, I. Arabidopsis CBF5 interacts with the H/ACA snoRNP assembly factor NAF1. Plant Mol. Biol. 2007, 65, 615–626. [Google Scholar] [CrossRef]
- Pendle, A.F.; Clark, G.P.; Boon, R.; Lewandowska, D.; Lam, Y.W.; Andersen, J.; Mann, M.; Lamond, A.I.; Brown, J.W.S.; Shaw, P.J. Proteomic analysis of the Arabidopsis nucleolus suggests novel nucleolar functions. Mol. Biol. Cell 2005, 16, 260–269. [Google Scholar] [CrossRef]
- Rossignol, P.; Collier, S.; Bush, M.; Shaw, P.; Doonan, J.H. Arabidopsis POT1A interacts with TERT-V(18), an N-terminal splicing variant of telomerase. J. Cell Sci. 2007, 120, 3678–3687. [Google Scholar] [CrossRef]
- Nakamura, T.M.; Morin, G.B.; Chapman, K.B.; Weinrich, S.L.; Andrews, W.H.; Lingner, J.; Harley, C.B.; Cech, T.R. Telomerase catalytic subunit homologs from fission yeast and human. Science 1997, 277, 955–959. [Google Scholar] [CrossRef]
- Oguchi, K.; Liu, H.T.; Tamura, K.; Takahashi, H. Molecular cloning and characterization of AtTERT, a telomerase reverse transcriptase homolog in Arabidopsis thaliana. Febs Lett. 1999, 457, 465–469. [Google Scholar] [CrossRef]
- Baumann, P.; Cech, T.R. Pot1, the putative telomere end-binding protein in fission yeast and humans. Science 2001, 292, 1171–1175. [Google Scholar] [CrossRef]
- Houghtaling, B.R.; Cuttonaro, L.; Chang, W.; Smith, S. A dynamic molecular link between the telomere length regulator TRF1 and the chromosome end protector TRF2. Curr. Biol. 2004, 14, 1621–1631. [Google Scholar] [CrossRef]
- Liu, D.; Safari, A.; O’Connor, M.S.; Chan, D.W.; Laegeler, A.; Qin, J.; Zhou, S.Y. PTOP interacts with POT1 and regulates its localization to telomeres. Nat. Cell Biol. 2004, 6, 673–680. [Google Scholar] [CrossRef]
- Ye, J.Z.S.; Hockemeyer, D.; Krutchinsky, A.N.; Loayza, D.; Hooper, S.M.; Chait, B.T.; de Lange, T. POT1-interacting protein PIP1: A telomere length regulator that recruits POT1 to the TIN2/TRF1 complex. Genes Dev. 2004, 18, 1649–1654. [Google Scholar] [CrossRef]
- Chen, L.Y.; Redon, S.; Lingner, J. The human CST complex is a terminator of telomerase activity. Nature 2012, 488, 540. [Google Scholar] [CrossRef]
- Tani, A.; Murata, M. Alternative splicing of Pot1 (Protection of telomere)-like genes in Arabidopsis thaliana. Genes Genet. Syst. 2005, 80, 41–48. [Google Scholar] [CrossRef]
- Cifuentes-Rojas, C.; Kannan, K.; Tseng, L.; Shippen, D.E. Two RNA subunits and POT1a are components of Arabidopsis telomerase. Proc. Natl. Acad. Sci. USA 2011, 108, 73–78. [Google Scholar] [CrossRef]
- Kannan, K.; Nelson, A.D.L.; Shippen, D.E. Dyskerin is a component of the Arabidopsis telomerase RNP required for telomere maintenance. Mol. Cell. Biol. 2008, 28, 2332–2341. [Google Scholar] [CrossRef]
- Arora, A.; Beilstein, M.A.; Shippen, D.E. Evolution of Arabidopsis protection of telomeres 1 alters nucleic acid recognition and telomerase regulation. Nucleic Acids Res. 2016, 44, 9821–9830. [Google Scholar] [CrossRef]
- Van Steensel, B.; de Lange, T. Control of telomere length by the human telomeric protein TRF1. Nature 1997, 385, 740–743. [Google Scholar] [CrossRef]
- Kim, S.H.; Kaminker, P.; Campisi, J. TIN2, a new regulator of telomere length in human cells. Nat. Genet. 1999, 23, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.Z.S.; de Lange, T. TIN2 is a tankyrase 1 PARP modulator in the TRF1 telomere length control complex. Nat. Genet. 2004, 36, 618–623. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.Z.; Lu, K.P. The Pin2/TRF1-interacting: Protein PinX1 is a potent telomerase inhibitor. Cell 2001, 107, 347–359. [Google Scholar] [CrossRef]
- Wu, Y.; Xiao, S.; Zhu, X.D. MRE11-RAD50-NBS1 and ATM function as co-mediators of TRF1 in telomere length control. Nat. Struct. Mol. Biol. 2007, 14, 832–840. [Google Scholar] [CrossRef]
- Schrumpfova, P.; Kuchar, M.; Mikova, G.; Skrisovska, L.; Kubicarova, T.; Fajkus, J. Characterization of two Arabidopsis thaliana myb-like proteins showing affinity to telomeric DNA sequence. Genome 2004, 47, 316–324. [Google Scholar] [CrossRef]
- Schrumpfova, P.P.; Kuchar, M.; Palecek, J.; Fajkus, J. Mapping of interaction domains of putative telomere-binding proteins AtTRB1 and AtPOT1b from Arabidopsis thaliana. Febs Lett. 2008, 582, 1400–1406. [Google Scholar] [CrossRef]
- Schrumpfova, P.P.; Vychodilova, I.; Dvorackova, M.; Majerska, J.; Dokladal, L.; Schorova, S.; Fajkus, J. Telomere repeat binding proteins are functional components of Arabidopsis telomeres and interact with telomerase. Plant J. 2014, 77, 770–781. [Google Scholar] [CrossRef] [Green Version]
- Schrumpfova, P.P.; Vychodilova, I.; Hapala, J.; Schorova, S.; Dvoracek, V.; Fajkus, J. Telomere binding protein TRB1 is associated with promoters of translation machinery genes in vivo. Plant Mol. Biol. 2016, 90, 189–206. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, Y.J.; Krause, K.; Yang, T.T.; Dongus, J.A.; Zhang, Y.J.; Turck, F. Telobox motifs recruit CLF/SWN-PRC2 for H3K27me3 deposition via TRB factors in Arabidopsis. Nat. Genet. 2018, 50, 638. [Google Scholar] [CrossRef]
- Dokladal, L.; Benkova, E.; Honys, D.; Dupl’akova, N.; Lee, L.Y.; Gelvin, S.B.; Sykorova, E. An armadillo-domain protein participates in a telomerase interaction network. Plant Mol. Biol. 2018, 97, 407–420. [Google Scholar] [CrossRef]
- Lee, W.K.; Cho, M.H. Telomere-binding protein regulates the chromosome ends through the interaction with histone deacetylases in Arabidopsis thaliana. Nucleic Acids Res. 2016, 44, 4610–4624. [Google Scholar] [CrossRef]
- Tan, L.M.; Zhang, C.J.; Hou, X.M.; Shao, C.R.; Lu, Y.J.; Zhou, J.X.; Li, Y.Q.; Li, L.; Chen, S.; He, X.J. The PEAT protein complexes are required for histone deacetylation and heterochromatin silencing. Embo J. 2018, 37, e98770. [Google Scholar] [CrossRef] [PubMed]
- Van Steensel, B.; Smogorzewska, A.; de Lange, T. TRF2 protects human telomeres from end-to-end fusions. Cell 1998, 92, 401–413. [Google Scholar] [CrossRef]
- Kabir, S.; Sfeir, A.; de Lange, T. Taking apart Rap1 An adaptor protein with telomeric and non-telomeric functions. Cell Cycle 2010, 9, 4061–4067. [Google Scholar] [CrossRef] [PubMed]
- Rai, R.; Hu, C.; Broton, C.; Chen, Y.; Lei, M.; Chang, S. NBS1 Phosphorylation Status Dictates Repair Choice of Dysfunctional Telomeres. Mol. Cell 2017, 65, 801. [Google Scholar] [CrossRef]
- O’Connor, M.S.; Safari, A.; Liu, D.; Qin, J.; Zhou, S.Y. The human Rap1 protein complex and modulation of telomere length. J. Biol. Chem. 2004, 279, 28585–28591. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, Y.T.; van Overbeek, M.; Donigian, J.R.; Baciu, P.; de Lange, T.; Lei, M. A shared docking motif in TRF1 and TRF2 used for differential recruitment of telomeric proteins. Science 2008, 319, 1092–1096. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Jung, D.; Jung, Y.; Lee, S.G.; Lee, I. Interaction of human Ku70 with TRF2. Febs Lett. 2000, 481, 81–85. [Google Scholar] [CrossRef] [Green Version]
- Gomez, M.; Wu, J.; Schreiber, V.; Dunlap, J.; Dantzer, F.; Wang, Y.S.; Liu, Y. PARP1 is a TRF2-associated poly(ADP-ribose)polymerase and protects eroded telomere. Mol. Biol. Cell 2006, 17, 1686–1696. [Google Scholar] [CrossRef]
- Dantzer, F.; Giraud-Panis, M.J.; Jaco, I.; Ame, J.C.; Schultz, I.; Blasco, M.; Koering, C.E.; Gilson, E.; Menissier-de Murcia, J.; de Murcia, G.; et al. Functional interaction between poly(ADP-ribose) polymerase 2 (PARP-2) and TRF2: PARP activity negatively regulates TRF2. Mol. Cell. Biol. 2004, 24, 1595–1607. [Google Scholar] [CrossRef]
- Wu, Y.L.; Mitchell, T.R.H.; Zhu, X.D. Human XPF controls TRF2 and telomere length maintenance through distinctive mechanisms. Mech. Ageing Dev. 2008, 129, 602–610. [Google Scholar] [CrossRef]
- Chen, C.M.; Wang, C.T.; Ho, C.H. A plant gene encoding a Myb-like protein that binds telomeric GGTTTAG repeats in vitro. J. Biol. Chem. 2001, 276, 16511–16519. [Google Scholar] [CrossRef]
- Kuchar, M.; Fajkus, J. Interactions of putative telomere-binding proteins in Arabidopsis thaliana: Identification of functional TRF2 homolog in plants. Febs Lett. 2004, 578, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Karamysheva, Z.N.; Surovtseva, Y.V.; Vespa, L.; Shakirov, E.V.; Shippen, D.E. A C-terminal Myb extension domain defines a novel family of double-strand telomeric DNA-binding proteins in Arabidopsis. J. Biol. Chem. 2004, 279, 47799–47807. [Google Scholar] [CrossRef] [PubMed]
- Majerska, J.; Schrumpfova, P.P.; Dokladal, L.; Schorova, S.; Stejskal, K.; Oboril, M.; Honys, D.; Kozakova, L.; Polanska, P.S.; Sykorova, E. Tandem affinity purification of AtTERT reveals putative interaction partners of plant telomerase in vivo. Protoplasma 2017, 254, 1547–1562. [Google Scholar] [CrossRef]
- Jeong, S.A.; Kim, K.; Lee, J.H.; Cha, J.S.; Khadka, P.; Cho, H.S.; Chung, K. Akt-mediated phosphorylation increases the binding affinity of hTERT for importin alpha to promote nuclear translocation. J. Cell Sci. 2015, 128, 2287–2301. [Google Scholar] [CrossRef] [PubMed]
- Khurts, S.; Masutomi, K.; Delgermaa, L.; Arai, K.; Oishi, N.; Mizuno, H.; Hayashi, N.; Hahn, W.C.; Murakami, S. Nucleolin interacts with telomerase. J. Biol. Chem. 2004, 279, 51508–51515. [Google Scholar] [CrossRef] [PubMed]
- Pontvianne, F.; Abou-Ellail, M.; Douet, J.; Comella, P.; Matia, I.; Chandrasekhara, C.; DeBures, A.; Blevins, T.; Cooke, R.; Medina, F.J.; et al. Nucleolin Is Required for DNA Methylation State and the Expression of rRNA Gene Variants in Arabidopsis thaliana. PLoS Genet. 2010, 6, e1001225. [Google Scholar] [CrossRef]
- Pontvianne, F.; Carpentier, M.C.; Durut, N.; Pavlistova, V.; Jaske, K.; Schorova, S.; Parrinello, H.; Rohmer, M.; Pikaard, C.S.; Fojtova, M.; et al. Identification of Nucleolus-Associated Chromatin Domains Reveals a Role for the Nucleolus in 3D Organization of the A. thaliana Genome. Cell Rep. 2016, 16, 1574–1587. [Google Scholar] [CrossRef] [Green Version]
- Venteicher, A.S.; Meng, Z.J.; Mason, P.J.; Veenstra, T.D.; Artandi, S.E. Identification of ATPases pontin and reptin as telomerase components essential for holoenzyme assembly. Cell 2008, 132, 945–957. [Google Scholar] [CrossRef]
- Holt, B.F.; Boyes, D.C.; Ellerstrom, M.; Siefers, N.; Wiig, A.; Kauffman, S.; Grant, M.R.; Dangl, J.L. An evolutionarily conserved mediator of plant disease resistance gene function is required for normal Arabidopsis development. Dev. Cell 2002, 2, 807–817. [Google Scholar] [CrossRef]
- Giannone, R.J.; McDonald, H.W.; Hurst, G.B.; Shen, R.F.; Wang, Y.S.; Liu, Y. The Protein Network Surrounding the Human Telomere Repeat Binding Factors TRF1, TRF2, and POT1. PLoS ONE 2010, 5, e12407. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.Y.; Wu, F.H.; Hsu, C.T.; Shen, S.C.; Yeh, H.Y.; Liao, D.C.; Fang, M.J.; Liu, N.T.; Yen, Y.C.; Dokladal, L.; et al. Screening a cDNA Library for Protein-Protein Interactions Directly in Planta. Plant Cell 2012, 24, 1746–1759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kappei, D.; Butter, F.; Benda, C.; Scheibe, M.; Draskovic, I.; Stevense, M.; Novo, C.L.; Basquin, C.; Araki, M.; Araki, K.; et al. HOT1 is a mammalian direct telomere repeat-binding protein contributing to telomerase recruitment. Embo J. 2013, 32, 1681–1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, W.H.; Ford, L.P.; Lenertz, L.; Wright, W.E.; Shay, J.W. Human Ku70/80 associates physically with telomerase through interaction with hTERT. J. Biol. Chem. 2002, 277, 47242–47247. [Google Scholar] [CrossRef]
- Fell, V.L.; Schild-Poulter, C. The Ku heterodimer: Function in DNA repair and beyond. Mutat. Res. Rev. Mutat. Res. 2015, 763, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Bundock, P.; van Attikum, H.; Hooykaas, P. Increased telomere length and hypersensitivity to DNA damaging agents in an Arabidopsis KU70 mutant. Nucleic Acids Res. 2002, 30, 3395–3400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riha, K.; Watson, J.M.; Parkey, J.; Shippen, D.E. Telomere length deregulation and enhanced sensitivity to genotoxic stress in Arabidopsis mutants deficient in Ku70. Embo J. 2002, 21, 2819–2826. [Google Scholar] [CrossRef] [Green Version]
- West, C.E.; Waterworth, W.M.; Story, G.W.; Sunderland, P.A.; Jiang, Q.; Bray, C.M. Disruption of the Arabidopsis AtKu80 gene demonstrates an essential role for AtKu80 protein in efficient repair of DNA double-strand breaks in vivo. Plant J. 2002, 31, 517–528. [Google Scholar] [CrossRef]
- Gallego, M.E.; Jalut, N.; White, C.I. Telomerase dependence of telomere lengthening in Ku80 mutant Arabidopsis. Plant Cell 2003, 15, 782–789. [Google Scholar] [CrossRef]
- Cifuentes-Rojas, C.; Nelson, A.D.L.; Boltz, K.A.; Kannan, K.; She, X.T.; Shippen, D.E. An alternative telomerase RNA in Arabidopsis modulates enzyme activity in response to DNA damage. Genes Dev. 2012, 26, 2512–2523. [Google Scholar] [CrossRef] [Green Version]
- Valuchova, S.; Fulnecek, J.; Prokop, Z.; Stolt-Bergner, P.; Janouskova, E.; Hofr, C.; Riha, K. Protection of Arabidopsis blunt-ended telomeres is mediated by a physical association with the Ku heterodimer. Plant Cell 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holt, S.E.; Aisner, D.L.; Baur, J.; Tesmer, V.M.; Dy, M.; Ouellette, M.; Trager, J.B.; Morin, G.B.; Toft, D.O.; Shay, J.W.; et al. Functional requirement of p23 and Hsp90 in telomerase complexes. Genes Dev. 1999, 13, 817–826. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Zhong, D.B.; Monteiro, A. Comparative genomics and evolution of the HSP90 family of genes across all kingdoms of organisms. BMC Genom. 2006, 7, 156. [Google Scholar]
- Zhang, Z.M.; Sullivan, W.; Felts, S.J.; Prasad, B.D.; Toft, D.O.; Krishna, P. Characterization of plant p23-like proteins for their co-chaperone activities. Cell Stress Chaperones 2010, 15, 703–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wortman, M.J.; Johnson, E.M.; Bergemann, A.D. Mechanism of DNA binding and localized strand separation by Pur alpha and comparison with Pur family member, Pur beta. Biochim. Biophys. Acta-Mol. Cell Res. 2005, 1743, 64–78. [Google Scholar] [CrossRef] [PubMed]
- Mermoud, J.E.; Rowbotham, S.P.; Varga-Weisz, P.D. Keeping chromatin quiet How nucleosome remodeling restores heterochromatin after replication. Cell Cycle 2011, 10, 4017–4025. [Google Scholar] [CrossRef]
- Dona, M.; Scheid, O.M. DNA Damage Repair in the Context of Plant Chromatin. Plant Physiol. 2015, 168, 1206–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, D.; St-Sauveur, V.G.; Bergeron, D.; Dupuis-Sandoval, F.; Scott, M.S.; Bachand, F. A Polyadenylation-Dependent 3′ End Maturation Pathway Is Required for the Synthesis of the Human Telomerase RNA. Cell Rep. 2015, 13, 2244–2257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dokladal, L.; Honys, D.; Rana, R.; Lee, L.Y.; Gelvin, S.B.; Sykorova, E. cDNA Library Screening Identifies Protein Interactors Potentially Involved in Non-Telomeric Roles of Arabidopsis Telomerase. Front. Plant Sci. 2015, 6, 985. [Google Scholar] [CrossRef]
- Ma, H.L.; Su, L.; Yue, H.W.; Yin, X.L.; Zhao, J.; Zhang, S.L.; Kung, H.F.; Xu, Z.G.; Miao, J.Y. HMBOX1 interacts with MT2A to regulate autophagy and apoptosis in vascular endothelial cells. Sci. Rep. 2015, 5, 15121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.Y.; Luo, Z.H.; Jiang, S.; Li, F.; Han, X.; Hu, Y.; Wang, D.; Zhao, Y.; Ma, W.B.; Liu, D.; et al. The telomere-associated homeobox-containing protein TAH1/HMBOX1 participates in telomere maintenance in ALT cells. J. Cell Sci. 2013, 126, 3982–3989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamartine, J.; Seri, M.; Cinti, R.; Heitzmann, F.; Creaven, M.; Radomski, N.; Jost, E.; Lenoir, G.M.; Romeo, G.; Sylla, B.S. Molecular cloning and mapping of a human cDNA (PA2G4) that encodes a protein highly homologous to the mouse cell cycle protein p38-2G4. Cytogenet. Cell Genet. 1997, 78, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.L.; Funk, W.D.; Wang, S.S.; Weinrich, S.L.; Avilion, A.A.; Chiu, C.P.; Adams, R.R.; Chang, E.; Allsopp, R.C.; Yu, J.H.; et al. The Rna Component of Human Telomerase. Science 1995, 269, 1236–1241. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.B.; Graham, M.E.; Lovrecz, G.O.; Bache, N.; Robinson, P.J.; Reddel, R.R. Protein composition of catalytically active human telomerase from immortal cells. Science 2007, 315, 1850–1853. [Google Scholar] [CrossRef]
- Heiss, N.S.; Knight, S.W.; Vulliamy, T.J.; Klauck, S.M.; Wiemann, S.; Mason, P.J.; Poustka, A.; Dokal, I. X-linked dyskeratosis congenita is caused by mutations in a highly conserved gene with putative nucleolar functions. Nat. Genet. 1998, 19, 32–38. [Google Scholar] [CrossRef]
- Henras, A.; Henry, Y.; Bousquet-Antonelli, C.; Noaillac-Depeyre, J.; Gelugne, J.P.; Caizergues-Ferrer, M. Nhp2p and Nop10p are essential for the function of H/ACA snoRNPs. Embo J. 1998, 17, 7078–7090. [Google Scholar] [CrossRef] [Green Version]
- Saito, H.; Fujiwara, T.; Shin, S.; Okui, K.; Nakamura, Y. Cloning and mapping of a human novel cDNA (NHP2L1) that encodes a protein highly homologous to yeast nuclear protein NHP2. Cytogenet. Cell Genet. 1996, 72, 191–193. [Google Scholar] [CrossRef] [PubMed]
- Watkins, N.J.; Gottschalk, A.; Neubauer, G.; Kastner, B.; Fabrizio, P.; Mann, M.; Luhrmann, R. Cbf5p, a potential pseudouridine synthase, and Nhp2p, a putative RNA-binding protein, are present together with Gar1p in all H BOX/ACA-motif snoRNPs and constitute a common bipartite structure. RNA 1998, 4, 1549–1568. [Google Scholar] [CrossRef] [Green Version]
- Fatica, A.; Dlakic, M.; Tollervey, D. Naf1p is a box H/ACA snoRNP assembly factor. RNA 2002, 8, 1502–1514. [Google Scholar]
- Ting, N.S.Y.; Yu, Y.P.; Pohorelic, B.; Lees-Miller, S.P.; Beattie, T.L. Human Ku70/80 interacts directly with hTR, the RNA component of human telomerase. Nucleic Acids Res. 2005, 33, 2090–2098. [Google Scholar] [CrossRef] [Green Version]
- Sexton, A.N.; Collins, K. The 5′ Guanosine Tracts of Human Telomerase RNA Are Recognized by the G-Quadruplex Binding Domain of the RNA Helicase DHX36 and Function To Increase RNA Accumulation. Mol. Cell. Biol. 2011, 31, 736–743. [Google Scholar] [CrossRef] [PubMed]
- Moon, D.H.; Segal, M.; Boyraz, B.; Guinan, E.; Hofmann, I.; Cahan, P.; Tai, A.K.; Agarwal, S. Poly(A)-specific ribonuclease (PARN) mediates 3′-end maturation of the telomerase RNA component. Nat. Genet. 2015, 47, 1482. [Google Scholar] [CrossRef]
- Chiba, Y.; Johnson, M.A.; Lidder, P.; Vogel, J.T.; van Erp, H.; Green, P.J. AtPARN is an essential poly(A) ribonuclease in Arabidopsis. Gene 2004, 328, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Venteicher, A.S.; Abreu, E.B.; Meng, Z.J.; McCann, K.E.; Terns, R.M.; Veenstra, T.D.; Terns, M.P.; Artandi, S.E. A Human Telomerase Holoenzyme Protein Required for Cajal Body Localization and Telomere Synthesis. Science 2009, 323, 644–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sykorova, E.; Fajkus, J. Structure-function relationships in telomerase genes. Biol. Cell 2009, 101, 375–392. [Google Scholar] [CrossRef]
- Sykorova, E.; Fulneckova, J.; Mokros, P.; Fajkus, J.; Fojtova, M.; Peska, V. Three TERT genes in Nicotiana tabacum. Chromosome Res. 2012, 20, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, K.; Pearson, M.; Grate, L.; Sterne-Weiler, T.; Deans, J.; Donohue, J.P.; Ares, M. Structural RNAs of known and unknown function identified in malaria parasites by comparative genomics and RNA analysis. RNA 2007, 13, 1923–1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, C.J.; Zakian, V.A. Identification and characterization of the Schizosaccharomyces pombe TER1 telomerase RNA. Nat. Struct. Mol. Biol. 2008, 15, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, J.; Box, J.A.; Bunch, J.T.; Baumann, P. TER1, the RNA subunit of fission yeast telomerase. Nat. Struct. Mol. Biol. 2008, 15, 26–33. [Google Scholar] [CrossRef]
- Xie, M.Y.; Mosig, A.; Qi, X.; Li, Y.; Stadler, P.F.; Chen, J.J.L. Structure and function of the smallest vertebrate telomerase RNA from teleost fish. J. Biol. Chem. 2008, 283, 2049–2059. [Google Scholar] [CrossRef]
- Kachouri-Lafond, R.; Dujon, B.; Gilson, E.; Westhof, E.; Fairhead, C.; Teixeira, M.T. Large telomerase RNA, telomere length heterogeneity and escape from senescence in Candida glabrata. Febs Lett. 2009, 583, 3605–3610. [Google Scholar] [CrossRef]
- Gunisova, S.; Elboher, E.; Nosek, J.; Gorkovoy, V.; Brown, Y.; Lucier, J.F.; Laterreur, N.; Wellinger, R.J.; Tzfati, Y.; Tomaska, L. Identification and comparative analysis of telomerase RNAs from Candida species reveal conservation of functional elements. RNA-A Publ. RNA Soc. 2009, 15, 546–559. [Google Scholar] [CrossRef] [Green Version]
- Waldl, M.; Thiel, B.C.; Ochsenreiter, R.; Holzenleiter, A.; de Araujo Oliveira, J.V.; Walter, M.; Wolfinger, M.T.; Stadler, P.F. TERribly Difficult: Searching for Telomerase RNAs in Saccharomycetes. Genes (Basel) 2018, 9, 372. [Google Scholar] [CrossRef] [PubMed]
- Sykorova, E.; Lim, K.Y.; Kunicka, Z.; Chase, M.W.; Bennett, M.D.; Fajkus, J.; Leitch, A.R. Telomere variability in the monocotyledonous plant order Asparagales. Proc. R. Soc. B-Biol. Sci. 2003, 270, 1893–1904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sykorova, E.; Fajkus, J.; Meznikova, M.; Lim, K.Y.; Neplechova, K.; Blattner, F.R.; Chase, M.W.; Leitch, A.R. Minisatellite telomeres occur in the family Alliaceae but are lost in Allium. Am. J. Bot. 2006, 93, 814–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lange, T. Shelterin: The protein complex that shapes and safeguards human telomeres. Genes Dev. 2005, 19, 2100–2110. [Google Scholar] [CrossRef]
- De Lange, T. What I got wrong about shelterin. J. Biol. Chem. 2018, 293, 10453–10456. [Google Scholar] [CrossRef]
- Palm, W.; de Lange, T. How Shelterin Protects Mammalian Telomeres. Annu. Rev. Genet. 2008, 42, 301–334. [Google Scholar] [CrossRef]
- Sfeir, A.; de Lange, T. Removal of Shelterin Reveals the Telomere End-Protection Problem. Science 2012, 336, 593–597. [Google Scholar] [CrossRef] [Green Version]
- Kibe, T.; Zimmermann, M.; de Lange, T. TPP1 Blocks an ATR-Mediated Resection Mechanism at Telomeres. Mol. Cell 2016, 61, 236–246. [Google Scholar] [CrossRef]
- Zimmermann, M.; Lottersberger, F.; Buonomo, S.B.; Sfeir, A.; de Lange, T. 53BP1 Regulates DSB Repair Using Rif1 to Control 5 ‘ End Resection. Science 2013, 339, 700–704. [Google Scholar] [CrossRef] [PubMed]
- Dalby, A.B.; Hofr, C.; Cech, T.R. Contributions of the TEL-patch Amino Acid Cluster on TPP1 to Telomeric DNA Synthesis by Human Telomerase. J. Mol. Biol. 2015, 427, 1291–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latrick, C.M.; Cech, T.R. POT1-TPP1 enhances telomerase processivity by slowing primer dissociation and aiding translocation. Embo J. 2010, 29, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Nandakumar, J.; Bell, C.F.; Weidenfeld, I.; Zaug, A.J.; Leinwand, L.A.; Cech, T.R. The TEL patch of telomere protein TPP1 mediates telomerase recruitment and processivity. Nature 2012, 492, 285. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.C.; Cech, T.R. Human telomerase: Biogenesis, trafficking, recruitment, and activation. Genes Dev. 2015, 29, 1095–1105. [Google Scholar] [CrossRef] [PubMed]
- Griffith, J.D.; Comeau, L.; Rosenfield, S.; Stansel, R.M.; Bianchi, A.; Moss, H.; de Lange, T. Mammalian telomeres end in a large duplex loop. Cell 1999, 97, 503–514. [Google Scholar] [CrossRef]
- Stansel, R.M.; de Lange, T.; Griffith, J.D. T-loop assembly in vitro involves binding of TRF2 near the 3’ telomeric overhang. Embo J. 2001, 20, 5532–5540. [Google Scholar] [CrossRef]
- Sfeir, A.; Kosiyatrakul, S.T.; Hockemeyer, D.; MacRae, S.L.; Karlseder, J.; Schildkraut, C.L.; de Lange, T. Mammalian Telomeres Resemble Fragile Sites and Require TRF1 for Efficient Replication. Cell 2009, 138, 90–103. [Google Scholar] [CrossRef] [Green Version]
- Tong, A.S.; Stern, J.L.; Sfeir, A.; Kartawinata, M.; de Lange, T.; Zhu, X.D.; Bryan, T.M. ATM and ATR Signaling Regulate the Recruitment of Human Telomerase to Telomeres. Cell Rep. 2015, 13, 1633–1646. [Google Scholar] [CrossRef] [Green Version]
- Cesare, A.J.; Quinney, N.; Willcox, S.; Subramanian, D.; Griffith, J.D. Telomere looping in P-sativum (common garden pea). Plant J. 2003, 36, 271–279. [Google Scholar] [CrossRef]
- Mozgova, I.; Schrumpfova, P.P.; Hofr, C.; Fajkus, J. Functional characterization of domains in AtTRB1, a putative telomere-binding protein in Arabidopsis thaliana. Phytochemistry 2008, 69, 1814–1819. [Google Scholar] [CrossRef] [PubMed]
- Marian, C.O.; Bordoli, S.J.; Goltz, M.; Santarella, R.A.; Jackson, L.P.; Danilevskaya, O.; Beckstette, M.; Meeley, R.; Bass, H.W. The maize Single myb histone 1 gene, Smh1, belongs to a novel gene family and encodes a protein that binds telomere DNA repeats in vitro. Plant Physiol. 2003, 133, 1336–1350. [Google Scholar] [CrossRef]
- Bilaud, T.; Koering, C.E.; BinetBrasselet, E.; Ancelin, K.; Pollice, A.; Gasser, S.M.; Gilson, E. The telobox, a Myb-related telomeric DNA binding motif found in proteins from yeast, plants and human. Nucleic Acids Res. 1996, 24, 1294–1303. [Google Scholar] [CrossRef]
- Peska, V.; Schrumpfova, P.P.; Fajkus, J. Using the Telobox to Search for Plant Telomere Binding Proteins. Curr. Protein Pept. Sci. 2011, 12, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Schrumpfova, P.P.; Schorova, S.; Fajkus, J. Telomere- and Telomerase-Associated Proteins and Their Functions in the Plant Cell. Front. Plant Sci. 2016, 7, 851. [Google Scholar]
- Zhou, Y.; Hartwig, B.; James, G.V.; Schneeberger, K.; Turck, F. Complementary Activities of TELOMERE REPEAT BINDING Proteins and Polycomb Group Complexes in Transcriptional Regulation of Target Genes. Plant Cell 2016, 28, 87–101. [Google Scholar] [CrossRef]
- El Mai, M.; Wagner, K.D.; Michiels, J.F.; Ambrosetti, D.; Borderie, A.; Destree, S.; Renault, V.; Djerbi, N.; Giraud-Panis, M.J.; Gilson, E.; et al. The Telomeric Protein TRF2 Regulates Angiogenesis by Binding and Activating the PDGFR beta Promoter. Cell Rep. 2014, 9, 1047–1060. [Google Scholar] [CrossRef]
- Krutilina, R.I.; Oei, S.L.; Buchlow, G.; Yau, P.M.; Zalensky, A.O.; Zalenskaya, I.A.; Bradbury, E.M.; Tomilin, N.V. A negative regulator of telomere-length protein TRF1 is associated with interstitial (TTAGGG)n blocks in immortal Chinese hamster ovary cells. Biochem. Biophys. Res. Commun. 2001, 280, 471–475. [Google Scholar] [CrossRef]
- Martinez, P.; Thanasoula, M.; Carlos, A.R.; Gomez-Lopez, G.; Tejera, A.M.; Schoeftner, S.; Dominguez, O.; Pisano, D.G.; Tarsounas, M.; Blasco, M.A. Mammalian Rap1 controls telomere function and gene expression through binding to telomeric and extratelomeric sites. Nat. Cell Biol. 2010, 12, 768. [Google Scholar] [CrossRef]
- Morse, R.H. RAP, RAP, open up! New wrinkles for RAP1 in yeast. Trends Genet. 2000, 16, 51–53. [Google Scholar] [CrossRef]
- Rizzo, A.; Iachettini, S.; Salvati, E.; Zizza, P.; Maresca, C.; D’Angelo, C.; Benarroch-Popivker, D.; Capolupo, A.; del Gaudio, F.; Cosconati, S.; et al. SIRT6 interacts with TRF2 and promotes its degradation in response to DNA damage. Nucleic Acids Res. 2017, 45, 1820–1834. [Google Scholar] [CrossRef]
- Simonet, T.; Zaragosi, L.E.; Philippe, C.; Lebrigand, K.; Schouteden, C.; Augereau, A.; Bauwens, S.; Ye, J.; Santagostino, M.; Giulotto, E.; et al. The human TTAGGG repeat factors 1 and 2 bind to a subset of interstitial telomeric sequences and satellite repeats. Cell Res. 2011, 21, 1028–1038. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Renault, V.M.; Jamet, K.; Gilson, E. Transcriptional outcome of telomere signalling. Nat. Rev. Genet. 2014, 15, 491–503. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Pazin, M.J.; Schwartz, C.M.; Becker, K.G.; Wersto, R.P.; Dilley, C.M.; Mattson, M.P. Nontelomeric TRF2-REST Interaction Modulates Neuronal Gene Silencing and Fate of Tumor and Stem Cells. Curr. Biol. 2008, 18, 1489–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fulcher, N.; Riha, K. Using Centromere Mediated Genome Elimination to Elucidate the Functional Redundancy of Candidate Telomere Binding Proteins in Arabidopsis thaliana. Front. Genet. 2016, 6, 349. [Google Scholar] [CrossRef]
- Perrault, S.D.; Hornsby, P.J.; Betts, D.H. Global gene expression response to telomerase in bovine adrenocortical cells. Biochem. Biophys. Res. Commun. 2005, 335, 925–936. [Google Scholar] [CrossRef]
- Majerska, J.; Sykorova, E.; Fajkus, J. Non-telomeric activities of telomerase. Mol. Biosyst. 2011, 7, 1013–1023. [Google Scholar] [CrossRef] [PubMed]
- Park, J.I.; Venteicher, A.S.; Hong, J.Y.; Choi, J.; Jun, S.; Shkreli, M.; Chang, W.; Meng, Z.J.; Cheung, P.; Ji, H.; et al. Telomerase modulates Wnt signalling by association with target gene chromatin. Nature 2009, 460, 66–U77. [Google Scholar] [CrossRef]
- Freeling, M. Bias in Plant Gene Content Following Different Sorts of Duplication: Tandem, Whole-Genome, Segmental, or by Transposition. Annu. Rev. Plant Biol. 2009, 60, 433–453. [Google Scholar] [CrossRef]
- Mandakova, T.; Lysak, M.A. Chromosomal Phylogeny and Karyotype Evolution in x=7 Crucifer Species (Brassicaceae). Plant Cell 2008, 20, 2559–2570. [Google Scholar] [CrossRef] [Green Version]
- Price, C.M.; Boltz, K.A.; Chaiken, M.F.; Stewart, J.A.; Beilstein, M.A.; Shippen, D.E. Evolution of CST function in telomere maintenance. Cell Cycle 2010, 9, 3157–3165. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.Y.; Hsu, S.J.; Bhattacharjee, A.; Wang, Y.Y.; Diao, J.J.; Price, C.M. CTC1-STN1 terminates telomerase while STN1-TEN1 enables C-strand synthesis during telomere replication in colon cancer cells. Nat. Commun. 2018, 9, 2827. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.Y.; Hsu, S.J.; Kasbek, C.; Chaiken, M.; Price, C.M. CTC1-mediated C-strand fill-in is an essential step in telomere length maintenance. Nucleic Acids Res. 2017, 45, 4281–4293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, J.A.; Wang, F.; Chaiken, M.F.; Kasbek, C.; Chastain, P.D.; Wright, W.E.; Price, C.M. Human CST promotes telomere duplex replication and general replication restart after fork stalling. Embo J. 2012, 31, 3537–3549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedoyan, J.K.; Lejnine, S.; Makarov, V.L.; Langmore, J.P. Condensation of rat telomere-specific nucleosomal arrays containing unusually short DNA repeats and histone H1. J. Biol. Chem. 1996, 271, 18485–18493. [Google Scholar] [CrossRef] [PubMed]
- Lejnine, S.; Makarov, V.L.; Langmore, J.P. Conserved Nucleoprotein Structure at the Ends of Vertebrate and Invertebrate Chromosomes. Proc. Natl. Acad. Sci. USA 1995, 92, 2393–2397. [Google Scholar] [CrossRef] [PubMed]
- Makarov, V.L.; Lejnine, S.; Bedoyan, J.; Langmore, J.P. Nucleosomal Organization of Telomere-Specific Chromatin in Rat. Cell 1993, 73, 775–787. [Google Scholar] [CrossRef]
- Tommerup, H.; Dousmanis, A.; Delange, T. Unusual Chromatin in Human Telomeres. Mol. Cell. Biol. 1994, 14, 5777–5785. [Google Scholar] [CrossRef] [PubMed]
- Fajkus, J.; Kovarik, A.; Kralovics, R.; Bezdek, M. Organization of Telomeric and Subtelomeric Chromatin in the Higher-Plant Nicotiana tabacum. Mol. Gen. Genet. 1995, 247, 633–638. [Google Scholar] [CrossRef]
- Dejardin, J.; Kingston, R.E. Purification of Proteins Associated with Specific Genomic Loci. Cell 2009, 136, 175–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fajkus, J.; Trifonov, E.N. Columnar packing of telomeric nucleosomes. Biochem. Biophys. Res. Commun. 2001, 280, 961–963. [Google Scholar] [CrossRef]
- Bosco, N.; de Lange, T. A TRF1-controlled common fragile site containing interstitial telomeric sequences. Chromosoma 2012, 121, 465–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Karow, J.K.; Hickson, I.D.; Maizels, N. The Bloom’s syndrome helicase unwinds G4 DNA. J. Biol. Chem. 1998, 273, 27587–27592. [Google Scholar] [CrossRef] [PubMed]
- Muftuoglu, M.; Wong, H.K.; Imam, S.Z.; Wilson, D.M.; Bohr, V.A.; Opresko, P.L. Telomere repeat binding factor 2 interacts with base excision repair proteins and stimulates DNA synthesis by DNA polymerase beta. Cancer Res. 2006, 66, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Tatsumi, Y.; Ezura, K.; Yoshida, K.; Yugawa, T.; Narisawa-Saito, M.; Kiyono, T.; Ohta, S.; Obuse, C.; Fujita, M. Involvement of human ORC and TRF2 in pre-replication complex assembly at telomeres. Genes Cells 2008, 13, 1045–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarek, G.; Vannier, J.B.; Panier, S.; Petrini, J.H.J.; Boulton, S.J. TRF2 Recruits RTEL1 to Telomeres in S Phase to Promote T-Loop Unwinding. Mol. Cell 2015, 57, 622–635. [Google Scholar] [CrossRef]
- Karlseder, J.; Hoke, K.; Mirzoeva, O.K.; Bakkenist, C.; Kastan, M.B.; Petrini, J.H.J.; de Lange, T. The telomeric protein TRF2 binds the ATM kinase and can inhibit the ATM-dependent DNA damage response. PLoS Biol. 2004, 2, 1150–1156. [Google Scholar] [CrossRef]
- Hwang, M.G.; Chung, I.K.; Kang, B.G.; Cho, M.H. Sequence-specific binding property of Arabidopsis thaliana telomeric DNA binding protein 1 (AtTBP1). Febs Lett. 2001, 503, 35–40. [Google Scholar] [CrossRef]
- Renfrew, K.B.; Song, X.Y.; Lee, J.R.; Arora, A.; Shippen, D.E. POT1a and Components of CST Engage Telomerase and Regulate Its Activity in Arabidopsis. PLoS Genet. 2014, 10, e1004738. [Google Scholar] [CrossRef]
- Wyatt, H.D.M.; Tsang, A.R.; Lobb, D.A.; Beattie, T.L. Human Telomerase Reverse Transcriptase (hTERT) Q169 Is Essential for Telomerase Function In Vitro and In Vivo. PLoS ONE 2009, 4, e7176. [Google Scholar] [CrossRef]
- Ganduri, S.; Lue, N.F. STN1-POLA2 interaction provides a basis for primase-pol alpha stimulation by human STN1. Nucleic Acids Res. 2017, 45, 9455–9466. [Google Scholar] [CrossRef] [PubMed]
- Miyake, Y.; Nakamura, M.; Nabetani, A.; Shimamura, S.; Tamura, M.; Yonehara, S.; Saito, M.; Ishikawa, F. RPA-like Mammalian Ctc1-Stn1-Ten1 Complex Binds to Single-Stranded DNA and Protects Telomeres Independently of the Pot1 Pathway. Mol. Cell 2009, 36, 193–206. [Google Scholar] [CrossRef]
- Derboven, E.; Ekker, H.; Kusenda, B.; Bulankova, P.; Riha, K. Role of STN1 and DNA Polymerase alpha in Telomere Stability and Genome-Wide Replication in Arabidopsis. PLoS Genet. 2014, 10, e1004682. [Google Scholar] [CrossRef] [PubMed]
- Leehy, K.A.; Lee, J.R.; Song, X.Y.; Renfrew, K.B.; Shippen, D.E. MERISTEM DISORGANIZATION1 Encodes TEN1, an Essential Telomere Protein That Modulates Telomerase Processivity in Arabidopsis. Plant Cell 2013, 25, 1343–1354. [Google Scholar] [CrossRef]
- Song, X.Y.; Leehy, K.; Warrington, R.T.; Lamb, J.C.; Surovtseva, Y.V.; Shippen, D.E. STN1 protects chromosome ends in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2008, 105, 19815–19820. [Google Scholar] [CrossRef]
- Surovtseva, Y.V.; Churikov, D.; Boltz, K.A.; Song, X.Y.; Lamb, J.C.; Warrington, R.; Leehy, K.; Heacock, M.; Price, C.M.; Shippen, D.E. Conserved Telomere Maintenance Component 1 Interacts with STN1 and Maintains Chromosome Ends in Higher Eukaryotes. Mol. Cell 2009, 36, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.H.; Kwon, C.; Lee, M.M.; Chung, I.K. Single-stranded DNA binding factor AtWHY1 modulates telomere length homeostasis in Arabidopsis. Plant J. 2007, 49, 442–451. [Google Scholar] [CrossRef]
- Kwon, C.; Chung, I.K. Interaction of an Arabidopsis RNA-binding protein with plant single-stranded telomeric DNA modulates telomerase activity. J. Biol. Chem. 2004, 279, 12812–12818. [Google Scholar] [CrossRef]
- Li, E.; Zhang, Y. DNA Methylation in Mammals. Cold Spring Harb. Perspect. Biol. 2014, 6, a019133. [Google Scholar] [CrossRef]
- Lister, R.; Pelizzola, M.; Dowen, R.H.; Hawkins, R.D.; Hon, G.; Tonti-Filippini, J.; Nery, J.R.; Lee, L.; Ye, Z.; Ngo, Q.M.; et al. Human DNA methylomes at base resolution show widespread epigenomic differences. Nature 2009, 462, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X. The epigenetic landscape of plants. Science 2008, 320, 489–492. [Google Scholar] [CrossRef]
- Cokus, S.J.; Feng, S.H.; Zhang, X.Y.; Chen, Z.G.; Merriman, B.; Haudenschild, C.D.; Pradhan, S.; Nelson, S.F.; Pellegrini, M.; Jacobsen, S.E. Shotgun bisulphite sequencing of the Arabidopsis genome reveals DNA methylation patterning. Nature 2008, 452, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Ogrocka, A.; Polanska, P.; Majerova, E.; Janeba, Z.; Fajkus, J.; Fojtova, M. Compromised telomere maintenance in hypomethylated Arabidopsis thaliana plants. Nucleic Acids Res. 2014, 42, 2919–2931. [Google Scholar] [CrossRef] [PubMed]
- Vrbsky, J.; Akimcheva, S.; Watson, J.M.; Turner, T.L.; Daxinger, L.; Vyskot, B.; Aufsatz, W.; Riha, K. siRNA-Mediated Methylation of Arabidopsis Telomeres. PLoS Genet. 2010, 6, e1000986. [Google Scholar] [CrossRef] [PubMed]
- Majerova, E.; Fojtova, M.; Mozgova, I.; Bittova, M.; Fajkus, J. Hypomethylating drugs efficiently decrease cytosine methylation in telomeric DNA and activate telomerase without affecting telomere lengths in tobacco cells. Plant Mol. Biol. 2011, 77, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Majerova, E.; Mandakova, T.; Vu, G.T.H.; Fajkus, J.; Lysak, M.A.; Fojtova, M. Chromatin features of plant telomeric sequences at terminal vs. internal positions. Front. Plant Sci. 2014, 5, 593. [Google Scholar] [CrossRef]
- Xie, X.Y.; Shippen, D.E. DDM1 guards against telomere truncation in Arabidopsis. Plant Cell Rep. 2018, 37, 501–513. [Google Scholar] [CrossRef]
- Fojtova, M.; Fajkus, J. Epigenetic Regulation of Telomere Maintenance. Cytogenet. Genome Res. 2014, 143, 125–135. [Google Scholar] [CrossRef]
- Fransz, P.; ten Hoopen, R.; Tessadori, F. Composition and formation of heterochromatin in Arabidopsis thaliana. Chromosome Res. 2006, 14, 71–82. [Google Scholar] [CrossRef]
- Roudier, F.; Ahmed, I.; Berard, C.; Sarazin, A.; Mary-Huard, T.; Cortijo, S.; Bouyer, D.; Caillieux, E.; Duvernois-Berthet, E.; Al-Shikhley, L.; et al. Integrative epigenomic mapping defines four main chromatin states in Arabidopsis. Embo J. 2011, 30, 1928–1938. [Google Scholar] [CrossRef] [PubMed]
- Schoeftner, S.; Blasco, M.A. A ‘higher order’ of telomere regulation: Telomere heterochromatin and telomeric RNAs. Embo J. 2009, 28, 2323–2336. [Google Scholar] [CrossRef] [PubMed]
- Cubiles, M.D.; Barroso, S.; Vaquero-Sedas, M.I.; Enguix, A.; Aguilera, A.; Vega-Palas, M.A. Epigenetic features of human telomeres. Nucleic Acids Res. 2018, 46, 2347–2355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenfeld, J.A.; Wang, Z.B.; Schones, D.E.; Zhao, K.; DeSalle, R.; Zhang, M.Q. Determination of enriched histone modifications in non-genic portions of the human genome. BMC Genom. 2009, 10, 143. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, R.J.; Kubicek, S.; Schreiber, S.L.; Karlseder, J. Reduced histone biosynthesis and chromatin changes arising from a damage signal at telomeres. Nat. Struct. Mol. Biol. 2010, 17, 1218. [Google Scholar] [CrossRef] [PubMed]
- Arnoult, N.; Van Beneden, A.; Decottignies, A. Telomere length regulates TERRA levels through increased trimethylation of telomeric H3K9 and HP1 alpha. Nat. Struct. Mol. Biol. 2012, 19, 948–956. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Cao, M.; O’Sullivan, R.; Peters, A.H.F.M.; Jenuwein, T.; Blasco, M.A. Epigenetic regulation of telomere length in mammalian cells by the Suv39h1 and Suv39h2 histone methyltransferases. Nat. Genet. 2004, 36, 94–99. [Google Scholar] [CrossRef]
- Saksouk, N.; Barth, T.K.; Ziegler-Birling, C.; Olova, N.; Nowak, A.; Rey, E.; Mateos-Langerak, J.; Urbach, S.; Reik, W.; Torres-Padilla, M.E.; et al. Redundant Mechanisms to Form Silent Chromatin at Pericentromeric Regions Rely on BEND3 and DNA Methylation. Mol. Cell 2014, 56, 580–594. [Google Scholar] [CrossRef]
- Benetti, R.; Gonzalo, S.; Jaco, I.; SChotta, G.; Klatt, P.; Jenuwein, T.; Blasco, M.A. Suv4-20h deficiency results in telomere elongation and derepression of telomere recombination. J. Cell Biol. 2007, 178, 925–936. [Google Scholar] [CrossRef] [Green Version]
- Gonzalo, S.; Jaco, I.; Fraga, M.F.; Chen, T.P.; Li, E.; Esteller, M.; Blasco, M.A. DNA methyltransferases control telomere length and telomere recombination in mammalian cells. Nat. Cell Biol. 2006, 8, 416. [Google Scholar] [CrossRef]
- Montero, J.J.; Lopez-Silanes, I.; Megias, D.; Fraga, M.F.; Castells-Garcia, A.; Blasco, M.A. TERRA recruitment of polycomb to telomeres is essential for histone trymethylation marks at telomeric heterochromatin. Nat. Commun. 2018, 9, 1548. [Google Scholar] [CrossRef]
- Sovakova, P.P.; Magdolenova, A.; Konecna, K.; Rajecka, V.; Fajkus, J.; Fojtova, M. Telomere elongation upon transfer to callus culture reflects the reprogramming of telomere stability control in Arabidopsis. Plant Mol. Biol. 2018, 98, 81–99. [Google Scholar] [CrossRef]
- Vaquero-Sedas, M.I.; Luo, C.Y.; Vega-Palas, M.A. Analysis of the epigenetic status of telomeres by using ChIP-seq data. Nucleic Acids Res. 2012, 40, e163. [Google Scholar] [CrossRef] [PubMed]
- Bulut-Karslioglu, A.; Perrera, V.; Scaranaro, M.; de la Rosa-Velazquez, I.A.; van de Nobelen, S.; Shukeir, N.; Popow, J.; Gerle, B.; Opravil, S.; Pagani, M.; et al. A transcription factor-based mechanism for mouse heterochromatin formation. Nat. Struct. Mol. Biol. 2012, 19, 1023. [Google Scholar] [CrossRef] [PubMed]
- Azzalin, C.M.; Reichenbach, P.; Khoriauli, L.; Giulotto, E.; Lingner, J. Telomeric repeat-containing RNA and RNA surveillance factors at mammalian chromosome ends. Science 2007, 318, 798–801. [Google Scholar] [CrossRef]
- Vaquero-Sedas, M.I.; Gamez-Arjona, F.M.; Vega-Palas, M.A. Arabidopsis thaliana telomeres exhibit euchromatic features. Nucleic Acids Res. 2011, 39, 2007–2017. [Google Scholar] [CrossRef] [PubMed]
- Tardat, M.; Dejardin, J. Telomere chromatin establishment and its maintenance during mammalian development. Chromosoma 2018, 127, 3–18. [Google Scholar] [CrossRef]
- Chai, W.H.; Du, Q.; Shay, J.W.; Wright, W.E. Human telomeres have different overhang sizes at leading versus lagging strands. Mol. Cell 2006, 21, 427–435. [Google Scholar] [CrossRef]
- Cimino-Reale, G.; Pascale, E.; Battiloro, E.; Starace, G.; Verna, R.; D’Ambrosio, E. The length of telomeric G-rich strand 3 ‘-overhang measured by oligonucleotide ligation assay. Nucleic Acids Res. 2001, 29, e35. [Google Scholar] [CrossRef]
- Makarov, V.L.; Hirose, Y.; Langmore, J.P. Long G tails at both ends of human chromosomes suggest a C strand degradation mechanism for telomere shortening. Cell 1997, 88, 657–666. [Google Scholar] [CrossRef]
- Wright, W.E.; Tesmer, V.M.; Huffman, K.E.; Levene, S.D.; Shay, J.W. Normal human chromosomes have long G-rich telomeric overhangs at one end. Genes Dev. 1997, 11, 2801–2809. [Google Scholar] [CrossRef] [Green Version]
- Oganesian, L.; Karlseder, J. Mammalian 5’ C-Rich Telomeric Overhangs Are a Mark of Recombination-Dependent Telomere Maintenance. Mol. Cell 2011, 42, 224–236. [Google Scholar] [CrossRef]
- Riha, K.; McKnight, T.D.; Fajkus, J.; Vyskot, B.; Shippen, D.E. Analysis of the G-overhang structures on plant telomeres: Evidence for two distinct telomere architectures. Plant J. 2000, 23, 633–641. [Google Scholar] [CrossRef]
- Kazda, A.; Zellinger, B.; Rossler, M.; Derboven, E.; Kusenda, B.; Riha, K. Chromosome end protection by blunt-ended telomeres. Genes Dev. 2012, 26, 1703–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fojtova, M.; Sykorova, E.; Najdekrova, L.; Polanska, P.; Zachova, D.; Vagnerova, R.; Angelis, K.J.; Fajkus, J. Telomere dynamics in the lower plant Physcomitrella patens. Plant Mol. Biol. 2015, 87, 591–601. [Google Scholar] [CrossRef]
- Riha, K.; Shippen, D.E. Ku is required for telomeric C-rich strand maintenance but not for end-to-end chromosome fusions in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 611–615. [Google Scholar] [CrossRef]
- Bryan, T.M.; Englezou, A.; DallaPozza, L.; Dunham, M.A.; Reddel, R.R. Evidence for an alternative mechanism for maintaining telomere length in human tumors and tumor-derived cell lines. Nat. Med. 1997, 3, 1271–1274. [Google Scholar] [CrossRef]
- Neumann, A.A.; Watson, C.M.; Noble, J.R.; Pickett, H.A.; Tam, P.P.L.; Reddel, R.R. Alternative lengthening of telomeres in normal mammalian somatic cells. Genes Dev. 2013, 27, 18–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruckova, E.; Friml, J.; Schrumpfova, P.P.; Fajkus, J. Role of alternative telomere lengthening unmasked in telomerase knock-out mutant plants. Plant Mol. Biol. 2008, 66, 637–646. [Google Scholar] [CrossRef]
- Zellinger, B.; Akimcheva, S.; Puizina, J.; Schirato, M.; Riha, K. Ku suppresses formation of telomeric circles and alternative telomere lengthening in Arabidopsis. Mol. Cell 2007, 27, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Karpenshif, Y.; Bernstein, K.A. From yeast to mammals: Recent advances in genetic control of homologous recombination. DNA Repair 2012, 11, 781–788. [Google Scholar] [CrossRef] [Green Version]
- Barber, L.J.; Youds, J.L.; Ward, J.D.; McIlwraith, M.J.; O’Neil, N.J.; Petalcorin, M.I.R.; Martin, J.S.; Collis, S.J.; Cantor, S.B.; Auclair, M.; et al. RTEL1 Maintains Genomic Stability by Suppressing Homologous Recombination. Cell 2008, 135, 261–271. [Google Scholar] [CrossRef] [Green Version]
- Uringa, E.J.; Lisaingo, K.; Pickett, H.A.; Brind’Amour, J.; Rohde, J.H.; Zelensky, A.; Essers, J.; Lansdorp, P.M. RTEL1 contributes to DNA replication and repair and telomere maintenance. Mol. Biol. Cell 2012, 23, 2782–2792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vannier, J.B.; Pavicic-Kaltenbrunner, V.; Petalcorin, M.I.R.; Ding, H.; Boulton, S.J. RTEL1 Dismantles T Loops and Counteracts Telomeric G4-DNA to Maintain Telomere Integrity. Cell 2012, 149, 795–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Guen, T.; Jullien, L.; Schertzer, M.; Lefebvre, A.; Kermasson, L.; de Villartay, J.P.; Londono-Vallejo, A.; Revy, P. RTEL1 (regulator of telomere elongation helicase 1), a DNA helicase essential for genome stability. Med. Sci. 2013, 29, 1138–1144. [Google Scholar]
- Vannier, J.B.; Sarek, G.; Boulton, S.J. RTEL1: Functions of a disease-associated helicase. Trends Cell Biol. 2014, 24, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Faure, G.; Revy, P.; Schertzer, M.; Londono-Vallejo, A.; Callebaut, I. The C-terminal extension of human RTEL1, mutated in Hoyeraal-Hreidarsson syndrome, contains Harmonin-N-like domains. Proteins-Struct. Funct. Bioinform. 2014, 82, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Margalef, P.; Kotsantis, P.; Borel, V.; Bellelli, R.; Panier, S.; Boulton, S.J. Stabilization of Reversed Replication Forks by Telomerase Drives Telomere Catastrophe. Cell 2018, 172, 439. [Google Scholar] [CrossRef]
- Hu, Z.B.; Cools, T.; Kalhorzadeh, P.; Heyman, J.; De Veylder, L. Deficiency of the Arabidopsis Helicase RTEL1 Triggers a SOG1-Dependent Replication Checkpoint in Response to DNA Cross-Links. Plant Cell 2015, 27, 149–161. [Google Scholar] [CrossRef]
- Recker, J.; Knoll, A.; Puchta, H. The Arabidopsis thaliana Homolog of the Helicase RTEL1 Plays Multiple Roles in Preserving Genome Stability. Plant Cell 2014, 26, 4889–4902. [Google Scholar] [CrossRef]
- Riha, K.; McKnight, T.D.; Griffing, L.R.; Shippen, D.E. Living with genome instability: Plant responses to telomere dysfunction. Science 2001, 291, 1797–1800. [Google Scholar] [CrossRef]
- Olivier, M.; Charbonnel, C.; Amiard, S.; White, C.I.; Gallego, M.E. RAD51 and RTEL1 compensate telomere loss in the absence of telomerase. Nucleic Acids Res. 2018, 46, 2432–2445. [Google Scholar] [CrossRef] [PubMed]
- Kamisugi, Y.; Whitaker, J.W.; Cuming, A.C. The Transcriptional Response to DNA-Double-Strand Breaks in Physcomitrella patens. PLoS ONE 2016, 11, e0161204. [Google Scholar] [CrossRef] [PubMed]
- Olsson, M.; Wapstra, E.; Friesen, C. Ectothermic telomeres: it’s time they came in from the cold. Philos. Trans. R. Soc. B-Biol. Sci. 2018, 373, 20160449. [Google Scholar] [CrossRef] [PubMed]
- Hoelzl, F.; Smith, S.; Cornils, J.S.; Aydinonat, D.; Bieber, C.; Ruf, T. Telomeres are elongated in older individuals in a hibernating rodent, the edible dormouse (Glis glis). Sci. Rep. 2016, 6, 36856. [Google Scholar] [CrossRef]
- Gomes, N.M.V.; Ryder, O.A.; Houck, M.L.; Charter, S.J.; Walker, W.; Forsyth, N.R.; Austad, S.N.; Venditti, C.; Pagel, M.; Shay, J.W.; et al. Comparative biology of mammalian telomeres: Hypotheses on ancestral states and the roles of telomeres in longevity determination. Aging Cell 2011, 10, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Lingner, J. Impact of oxidative stress on telomere biology. Differentiation 2018, 99, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.W.; Rane, G.; Dai, X.Y.; Shanmugam, M.K.; Arfuso, F.; Samy, R.P.; Lai, M.K.P.; Kappei, D.; Kumar, A.P.; Sethi, G. Ageing and the telomere connection: An intimate relationship with inflammation. Ageing Res. Rev. 2016, 25, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Bottcher, M.A.; Dingli, D.; Werner, B.; Traulsen, A. Replicative cellular age distributions in compartmentalized tissues. J. R. Soc. Interface 2018, 15, 20180272. [Google Scholar] [CrossRef]
- Cairns, J. Mutation Selection and Natural-History of Cancer. Nature 1975, 255, 197–200. [Google Scholar] [CrossRef]
- Conboy, M.J.; Karasov, A.O.; Rando, T.A. High incidence of non-random template strand segregation and asymmetric fate determination in dividing stem cells and their progeny. PLoS Biol. 2007, 5, 1120–1126. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Telomerase Calytic Subunit (TERT) Associated Proteins. | |||||
|---|---|---|---|---|---|
| Human TERT Associated Proteins | Protein Function and Direct Interactions | References | Arabidopsis TERT Associated Proteins | Protein Function and Direct Interactions | References |
| TERT | Catalytic subunit of telomerase | [48] | TERT | Catalytic subunit of telomerase | [49] |
| POT1 | Shelterin. Int.: telomeric ssDNA, TPP1 and CTC1. | [50,51,52,53,54] | POT1a | Shelterin-like. Int.: TERT, telomeric ssDNA, TER1, TRFL9, CBF5, RuvBL1, CTC1 and STN1. | [47,55,56,57,58] |
| TRF1 | Shelterin. Int.: telomeric dsDNA, TIN2, TANK1, PINX1, and ATM. | [59,60,61,62,63] | TRB1-3 | Shelterin-like. Int.: TERT, telomeric dsDNA, POT1b, RuvBL1 and RuvBL2a. | [64,65,66,67,68,69,70,71] |
| TRF2 | Shelterin. Int.: telomeric dsDNA; TIN2, NBS1, RAD50, Apollo, Ku70, PARP1, XPF-ERCC1, BLM, FEN1, POLB, ORC, RTEL1, ATM and HP1. | [61,72,73,74,75,76,77,78,79,80] | TRP1 | Possible non-telomeric functions of telomerase. Int.: TERT, telomere dsDNA in vitro, ARM, Ku70 and TRFL9. | [66,69,81,82,83] |
| TRFL2 | Possible non-telomeric functions of telomerase. Int.: TERT, telomere dsDNA in vitro and ARM. | [69,83] | |||
| TRFL11 | Associates with TERT. | [84] | |||
| KPNA1 | Promotes nuclear import of the TERT. | [85] | ImpA4 | Associates with TERT. | [84] |
| NCL | Involves nucleolar localization of TERT. | [86] | NUC-L1 | Role in telomere maintenance and telomere clustering. | [87,88] |
| pontin | Telomerase assembly. Int.: TERT and dyskerin. | [89] | RuvBL1 | Associates with TERT via TRBs, regulates telomerase activity. | [84,90] |
| reptin | Telomerase assembly. Int.: dyskerin. | [89] | RuvBL2a | Associates with TERT via TRBs, regulates telomerase activity. | [84] |
| ARMC6 | Int.: TRF2, telomerase. | [69,91] | ARM | May reflect posible non-telomeric functions of telomerase. Int.: TERT, TRP1, TRFL2, TRFL9 and CHR19. | [69,92] |
| TPP1 | Shelterin, mediates telomerase recruitment. Int.: TERT, POT1, TIN2, CTC1 and STN1. | [51,52,53,54,75] | n.a. | ||
| PINX1 | Potent telomerase inhibitor. Int.: TERT and TRF1. | [62] | n.a. | ||
| HOT1 | Int.: telomeric dsDNA, active telomerase. | [93] | n.a. | ||
| Ku70/80 | Int.: TERT, TR, TRF2 and RAP1. | [94,95] | Ku70/80 | Role in telomere length regulation, may protect blunt-ended telomeres Int.: TRP1, TER2 and TER2s. | [82,96,97,98,99,100,101] |
| Hsp90 | TERT assembly. Int.: TERT. | [102] | Hsp90 | NP_194150.1 | [103] |
| p23 | TERT assembly. Int.: TERT. | [102] | p23 | CAC16575, NP_683525 | [104] |
| Purα | p.h. Unwinds dsDNA telomeric oligonucleotides. | [105] | PURα1 | Associates with TERT. | [84] |
| SMARCAD1 | p.h. SWI/SNF-like protein that presumably associates with telomeres. | [106,107] | CHR19 | May reflect possible non-telomeric functions of telomerase. Int.: TERT, ARM, TRB1 and TRFL9. | [69] |
| PABPN1 | Promotes poly(A)-dependent TR 3′ end maturation. | [108] | RRM | Associates with TERT. | [92,109] |
| MT2A | p.h. Int.: HOT1. | [110,111] | MT2A | Associates with TERT. | [84,109] |
| PA2G4 | NP_006182.2 | [112] | G2p | Associates with TERT. | [84,109] |
| Telomerase RNA Associated Proteins | |||||
|---|---|---|---|---|---|
| Human TR Associated Proteins | Protein Function and Direct Interactions | References | Arabidopsis TR Associated Proteins | Protein Function and Direct Interactions | References |
| TR | RNA subunit of telomerase | [113] | TER1, TER2, TER2s | Putative RNA subunit of telomerase | [56,100] |
| TERT | Catalytic subunit of telomerase | [48,114] | TERT | Catalytic subunit of telomerase | [100] |
| Dyskerin | H/ACA snoRNPs, associated with nucleolus. Int.: TR, GAR1, NHP2, NOP10 and TCAB1. | [44,115] | CBF5 | H/ACA snoRNPs, Ath orthologue of Dyskerin, associated with nucleolus, subnuclear bodies and Cajal bodies, associated with telomerase RNP complex. Direct interaction with either of putative TERs not demonstrated. Int.: NAF1. | [45,57] |
| NOP10 | H/ACA snoRNPs, associates with nucleolus. Int.: TR and dyskerin. | [44,116] | NOP10 | H/ACA snoRNPs, Ath orthologue of NOP10, associates with nucleolus. | [45,46] |
| NHP2 | H/ACA snoRNPs, associates with nucleolus. Int.: TR, dyskerin and TCAB1. | [117,118] | NHP2 | H/ACA snoRNPs, Ath orthologue of NHP2, associates with nucleolus. | [45,46] |
| GAR1 | H/ACA snoRNPs, associated with nucleolus. Int.: dyskerin and TCAB1. | [44,118] | GAR1, 2 | H/ACA snoRNPs, Ath orthologues of GAR1, associate with nucleolus. | [45,46] |
| NAF1 | H/ACA snoRNPs, nucleolar shuttle - NAF1 is substituted by GAR1 during maturation of telomerase. Int.: dyskerin. | [119] | NAF1 | H/ACA snoRNPs, Ath orthologue of NAF1, associates with nucleolus and Cajal bodies. Int.: CBF5. | [45] |
| Ku70/80 | Int.: TR, TERT, TRF2 and RAP1. | [95,120] | Ku70/80 | Role in telomere length regulation, may protect blunt-ended telomeres Int.: TRP1, TER2 and TER2s. | [100] |
| pontin | Telomerase assembly. Int.: TERT and dyskerin. | [89] | RuvBL1 | Associates with TERT via TRBs, regulates telomerase activity. | Schorova et al., submitted |
| reptin | Telomerase assembly. Int.: dyskerin. | [89] | RuvBL2a | Associates with TERT via TRBs, regulates telomerase activity. | Schorova et al., submitted |
| RHAU | RNA helicase, unwinds a G4-quadruplex in human telomerase RNA. Int.: TR. | [121] | RHAU | NP_850255.1, NP_175298.2, NP_680142.2, NP_178223.2 | n.a. |
| PARN | Poly(A)-specific ribonuclease, 3′-end maturation of the TR. Int.: TR | [122] | PARN | Poly(A) degradation activity, essential gene first required during early development. | [123] |
| TCAB1 | H/ACA snoRNPs, driving telomerase to Cajal bodies. Int.: TR, dyskerin, NHP2 and GAR1. | [124] | TCAB1 | NP_193883.2 | n.a. |
| Telomeric dsDNA Associated Proteins | |||||
|---|---|---|---|---|---|
| Human Telomeric dsDNA Associated Proteins | Protein Function and Direct Interactions | References | Arabidopsis Telomeric dsDNA Associated Proteins | Protein Function and Direct Interactions | References |
| TRF1 | Shelterin. Int.: telomeric dsDNA, TIN2, TANK1 and PINX1. Non-telomeric: binding to ITS and chromatin and satellite DNA and modulation of their chromatin structure. Control of a common fragile site containing ITS. | [59,60,61,62] [162,182] | TRB1, 2, 3 | Shelterin-like. Int.: telomeric dsDNA, TERT, POT1b, RuvBL1 and RuvBL2a. Non-telomeric functions - a recruitment subunit of protein complexes involved in epigenetic regulations. Binding to ITSs. | [64,65,66]; Schorova et al., submitted [67,68,69,70,71] |
| TRF2 | Shelterin. Int.: telomeric dsDNA; TIN2, RAP1, NBS1, RAD50, Apollo, Ku70, PARP1, XPF-ERCC1, BLM, FEN1, POLB, ORC, RTEL1 and ATM. | [61,72,73,74,75,76,77,78,79,80,183,184,185,186,187] | TRP1 | Possible non-telomeric functions of telomerase. Int.: telomere dsDNA in vitro, TERT, ARM, Ku70, TRFL1 and TRFL9. | [66,69,81,82,83] |
| Non-telomeric function: transcriptional regulator. Binding to ITSs and satellite III. | [155,163] | TRFL2 | Possible non-telomeric functions of telomerase. Int.: telomere dsDNA in vitro, TERT and ARM. | [69,83] | |
| TRFL9 | Possible non-telomeric functions of telomerase. Int.: telomere dsDNA in vitro, TRP1 and POT1a. | [69,83] | |||
| TBP1, TRFL1, TRFL4 | Int.: telomere dsDNA in vitro. | [83,188] | |||
| HOT1 | Int.: telomeric dsDNA, active telomerase. | [93] | n.a. | ||
| Ku70/80 | The way of association with telomeric dsDNA is not fully elucidated. Int.: TRF2, RAP1, TR and TERT. | [95] | Ku70/80 | Role in telomere length regulation, may protect blunt-ended telomeres Int.: TRP1, TER2 and TER2s. | [82,96,97,98,99,101] |
| Telomeric ssDNA Associated Proteins | |||||
|---|---|---|---|---|---|
| Human Telomeric ssDNA Associated Proteins | Protein Function and Direct Interactions | References | Arabidopsis Telomeric ssDNA Associated Proteins | Protein Function and Direct Interactions | References |
| POT1 | Shelterin. Int.: telomeric ssDNA, TPP1 and CTC1. | [50,51,52,53,54] | POT1a | Shelterin-like. Int.: TERT, telomeric ssDNA, TER1, TRFL9, CBF5, RuvBL1, CTC1 and STN1. | [47,55,56,57,58,69,105,189] |
| POT1b | Shelterin-like. Int.: TRB1, TER2, TER2s. | [56,82,100] | |||
| POT1c | POT1 paralogue of unknown function. | [47] | |||
| TERT | Catalytic subunit of telomerase. | [190] | TERT | Catalytic subunit of telomerase. | |
| STN1 | CST complex subunit, prevents G-overhang overextension. Int.: CTC1, TEN1, TPP1 and POLA. | [54,172,191,192] | STN1 | CST complex subunit, controls access of telomerase and DDR, together with POLA may be involved in C-strand synthesis. Int.: CTC1, TEN1 and POT1a. Non-telomeric function. Facilitates re-replication at non-telomeric loci. | [189,193,194,195] |
| TEN1 | CST complex subunit, involves C-strand fill-in synthesis. Int.: STN1. | [172,192] | TEN1 | CST complex subunit, controls access of telomerase and DDR, coordinating synthesis of the C-strand. Int.: STN1. | [194] |
| CTC1 | CST complex subunit, prevents G-overhang overextension. Int.: telomeric ssDNA, STN1, TPP1 and POT1. | [54,192] | CTC1 | CST complex subunit, controls access of the telomerase and DDR, coordinating synthesis of the C-strand. Int.: STN1, POT1a and POLA. | [171,189,196] |
| Purα | p.h. Unwinds dsDNA telomeric oligonucleotides. | [105] | PURα1 | Associates with TERT. | [84] |
| n.a. | Why1 | Regulates telomere-length homeostasis. Int.: telomeric ssDNA. | [197] | ||
| n.a. | STEP1 | Truncated derivative of chloroplast RNA-binding protein, role in plant telomere biogenesis. Int.: telomeric ssDNA. | [198] | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Procházková Schrumpfová, P.; Fojtová, M.; Fajkus, J. Telomeres in Plants and Humans: Not So Different, Not So Similar. Cells 2019, 8, 58. https://doi.org/10.3390/cells8010058
Procházková Schrumpfová P, Fojtová M, Fajkus J. Telomeres in Plants and Humans: Not So Different, Not So Similar. Cells. 2019; 8(1):58. https://doi.org/10.3390/cells8010058
Chicago/Turabian StyleProcházková Schrumpfová, Petra, Miloslava Fojtová, and Jiří Fajkus. 2019. "Telomeres in Plants and Humans: Not So Different, Not So Similar" Cells 8, no. 1: 58. https://doi.org/10.3390/cells8010058
APA StyleProcházková Schrumpfová, P., Fojtová, M., & Fajkus, J. (2019). Telomeres in Plants and Humans: Not So Different, Not So Similar. Cells, 8(1), 58. https://doi.org/10.3390/cells8010058