Cilia Distal Domain: Diversity in Evolutionarily Conserved Structures

 ,
,  , ,
, ,
Abstract
:1. Introduction
The Discovery of the Basic Cilia Structure at a Glance
2. CILIA: Variations of a Basic Structure to Fulfill Myriad Functions
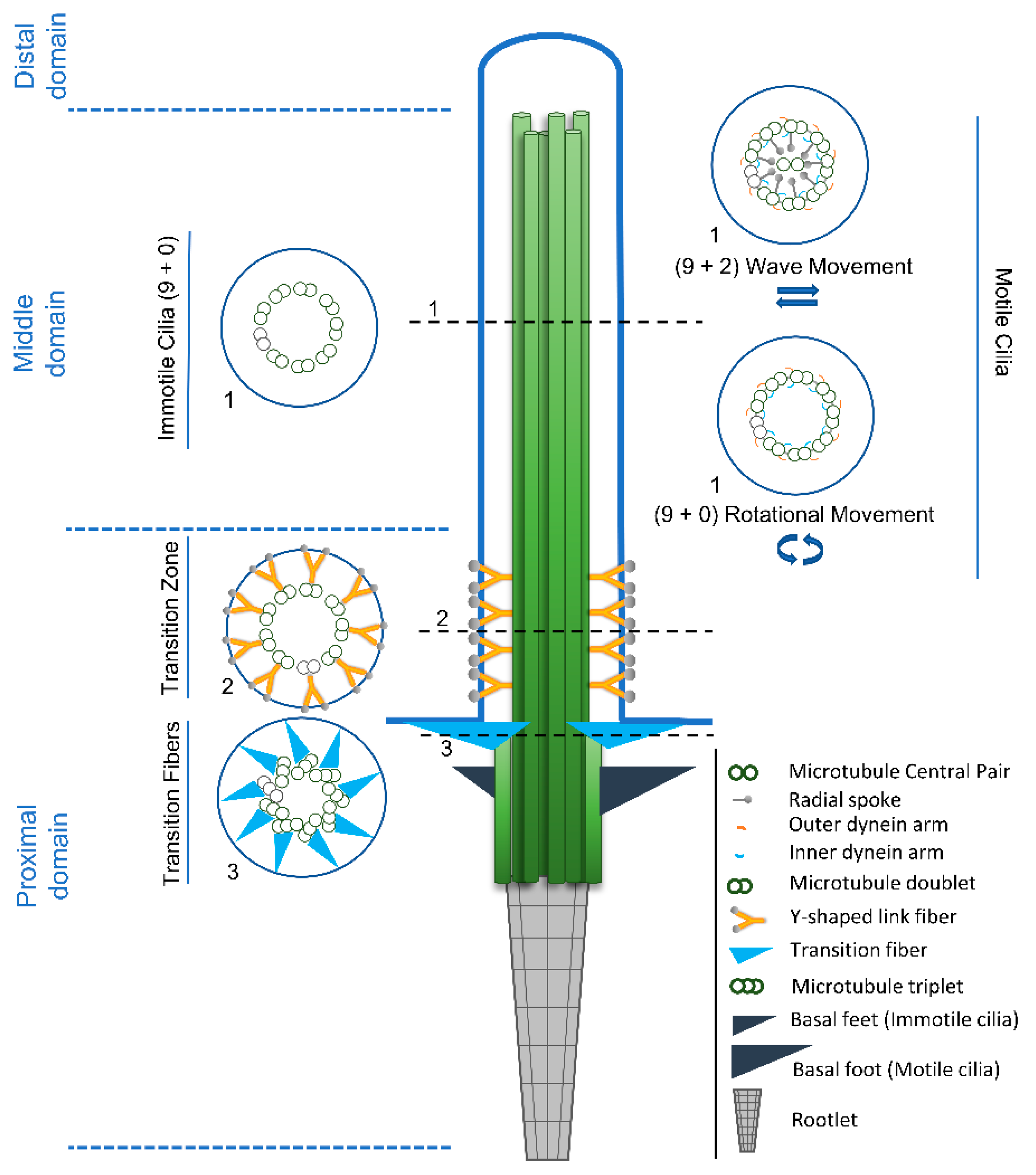
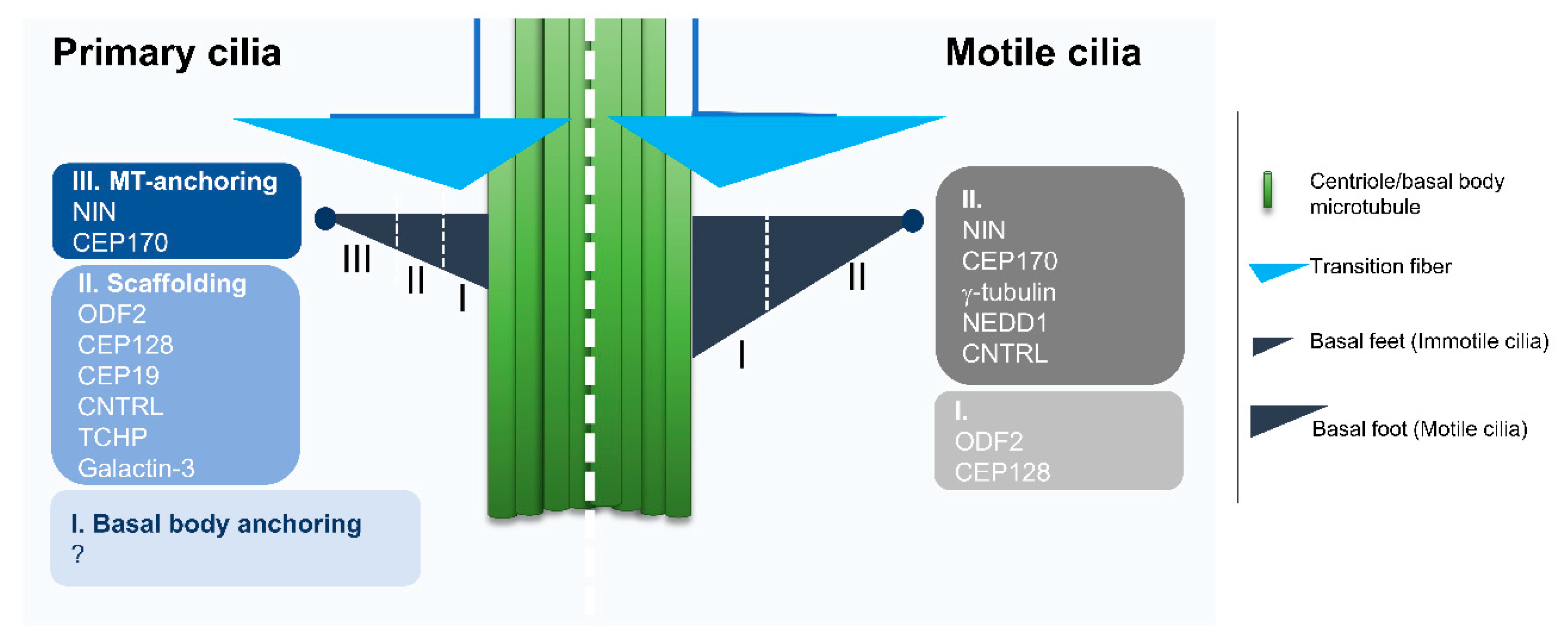
2.1. Diversity Starts at the Base: An Overview
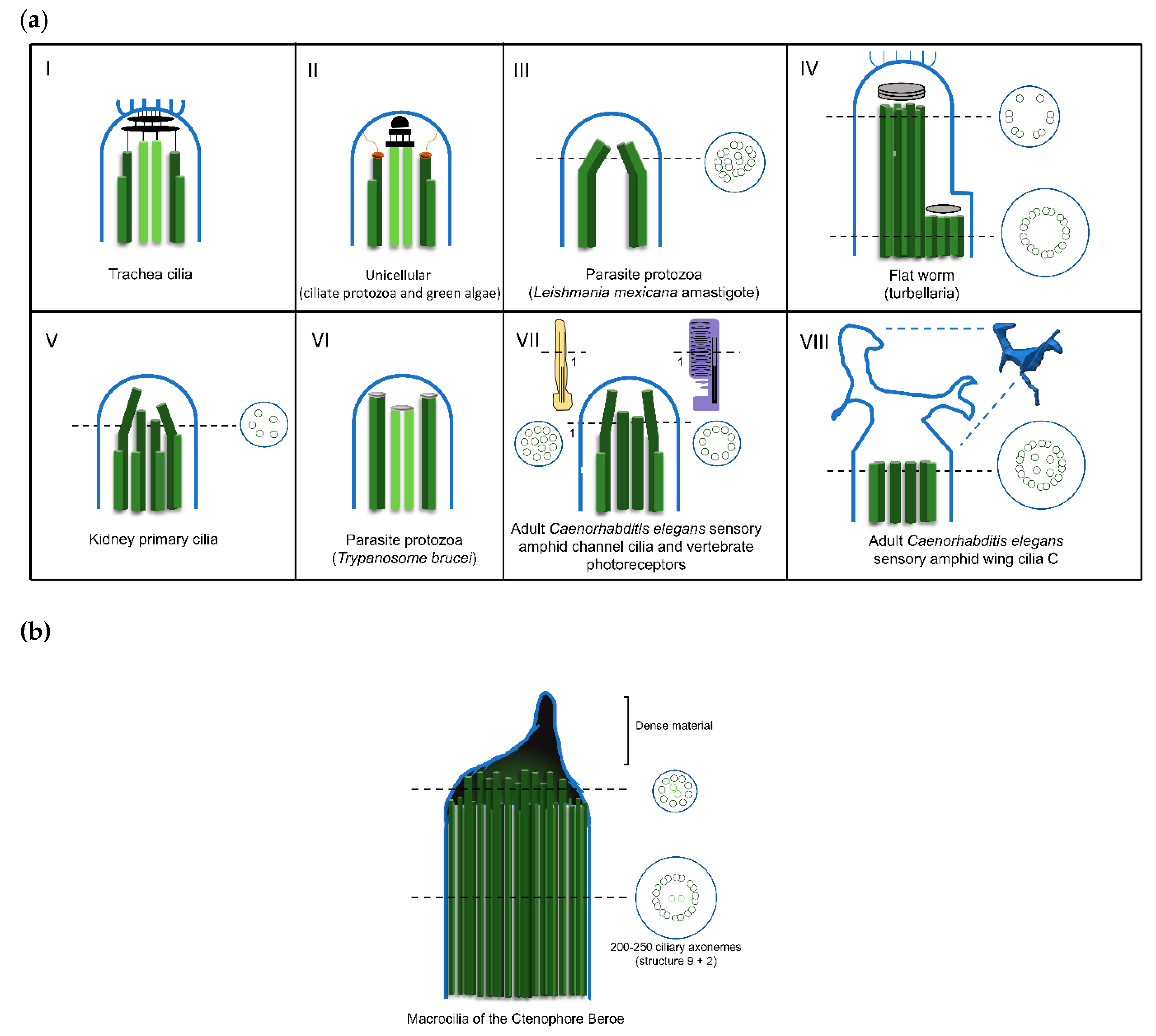
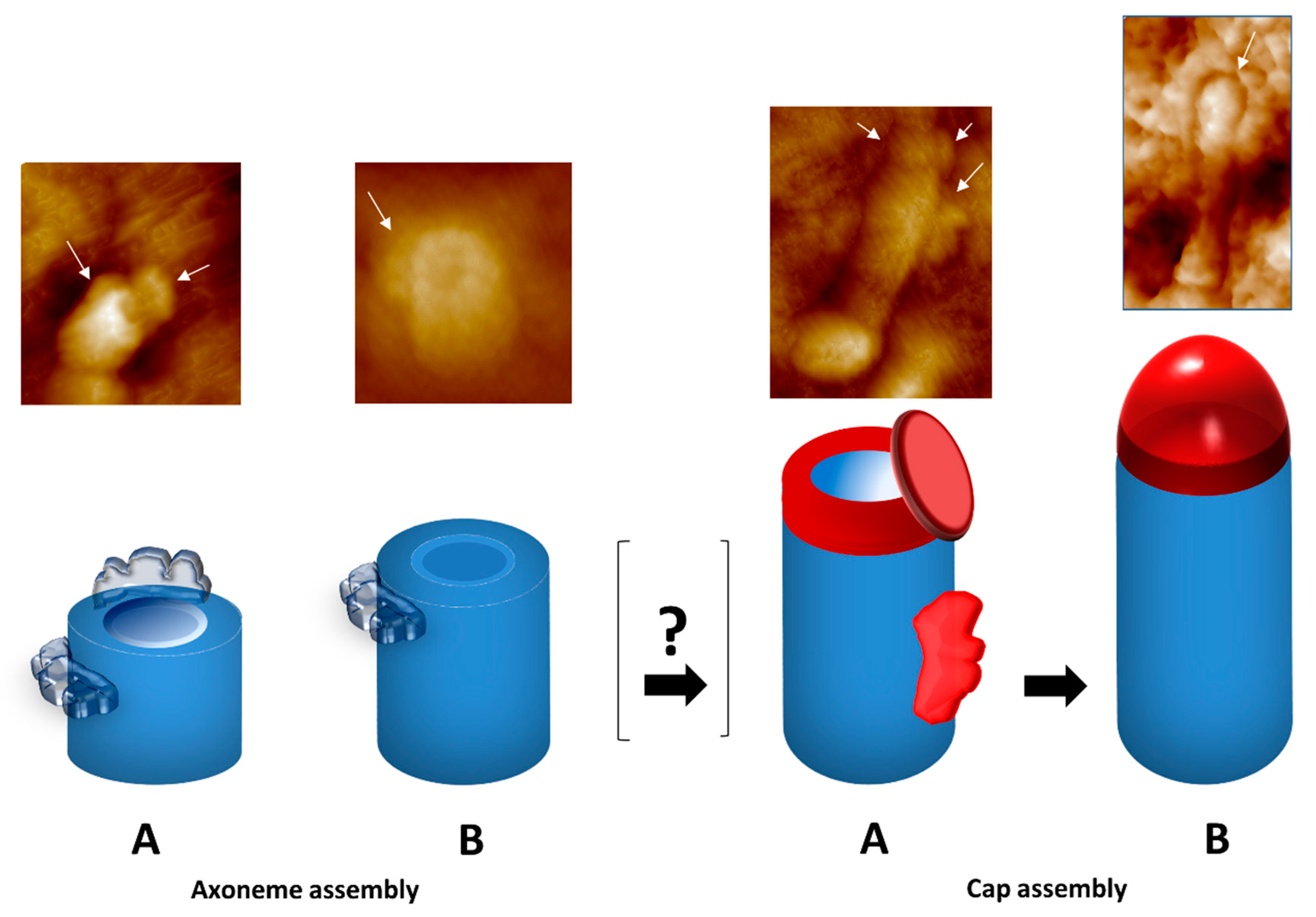
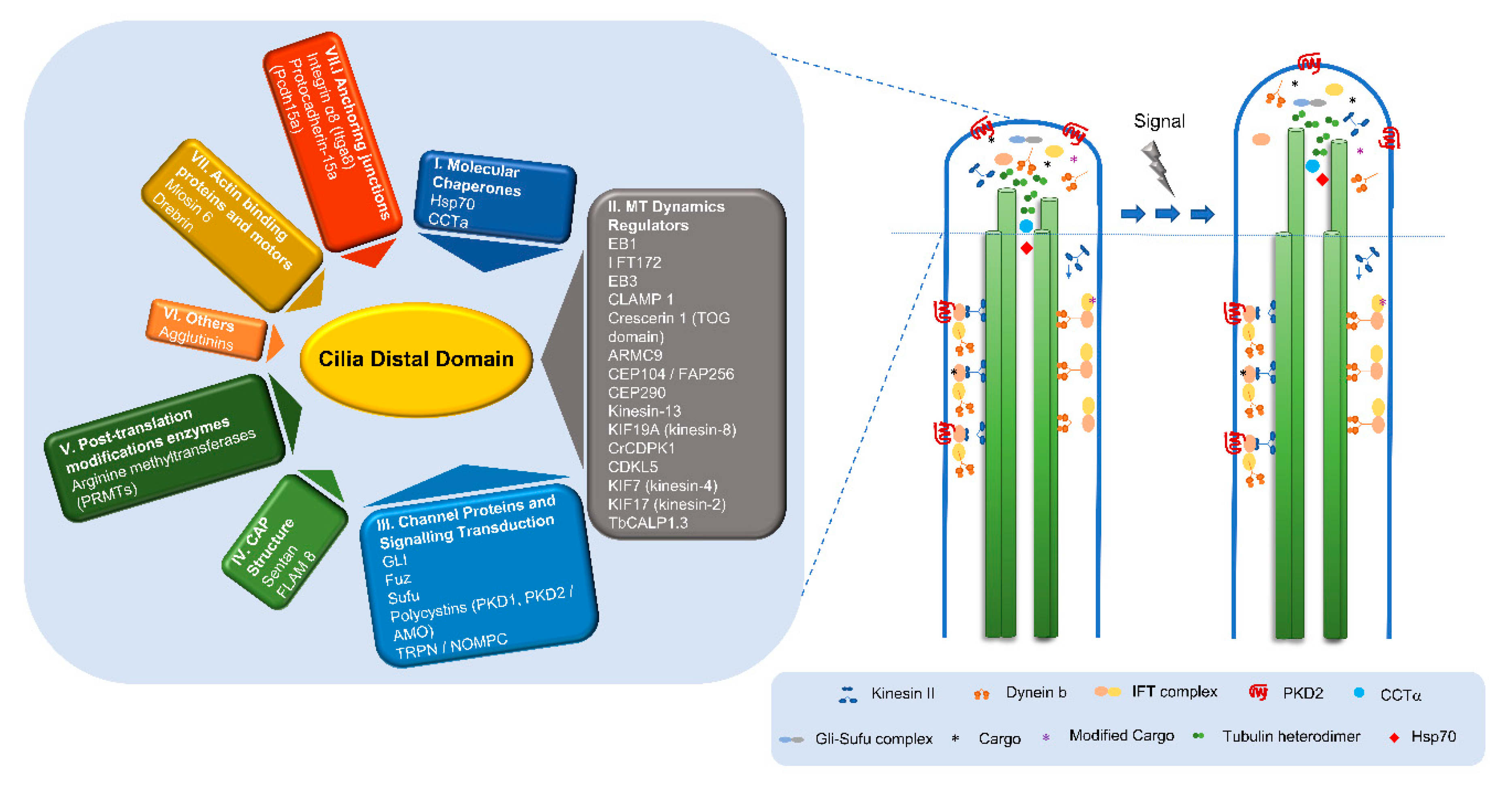
2.2. The Amazing Architecture Diversity of the Cilia Distal Domain: Cilia Tips Segments and Caps
2.3. The Distal Domain of Cilia: Components and Functions
2.4. What Can We Learn about the Functions of the Distal Domain from Proteins Residing There?
2.4.1. Molecular Chaperones and Cilia Distal Domain Dynamics
2.4.2. Regulators of Microtubule Dynamics at the Cilium Distal Domain
+TIPs and Related Proteins
Kinesin Proteins
Other Proteins Associated with Microtubules
2.4.3. Channel Proteins and Cilia Signaling Transduction
2.4.4. Post-Translational Modifications at the Cilia Distal Domain
2.4.5. Cap Structural Proteins
3. Concluding Remarks
Funding
Conflicts of Interest
References
- Waters, A.M.; Beales, P.L. Ciliopathies: An expanding disease spectrum. Pediatr. Nephrol. 2011, 26, 1039–1056. [Google Scholar] [CrossRef] [PubMed]
- Ostrowski, L.E.; Blackburn, K.; Radde, K.M.; Moyer, M.B.; Schlatzer, D.M.; Moseley, A.; Boucher, R.C. A proteomic analysis of human cilia: Identification of novel components. Mol. Cell Proteom. 2002, 1, 451–465. [Google Scholar] [CrossRef]
- Haimo, L.T.; Rosenbaum, J.L. Cilia, flagella, and microtubules. J. Cell Biol. 1981, 91, 125s–130s. [Google Scholar] [CrossRef] [PubMed]
- Dellinger, O.P. The cilium as a key to the structure of contractile protoplasm. J. Morphol. 1909, 20, 171–210. [Google Scholar] [CrossRef]
- Schmitt, F.O.; Hall, C.E.; Jakus, M.A. The Ultrastructure of Protoplasmic Fibrils. Biol. Symp. 1943, 10, 261–276. [Google Scholar]
- Harvey, E.B.; Anderson, T.F. The Spermatozoon And Fertilization Membrane Of Arbacia Punctulata As Shown By The Electron Microscope. Biol. Bull. 1943, 85, 151–156. [Google Scholar] [CrossRef]
- Longest, P.M. Structure of the cilia in Ectocarpus mitchellae and Codium decorticatum. J. Elisha Mitchell Sci. Soc. (Chapel Hill, NC) 1946, 62, 249–252. [Google Scholar]
- Jakus, M.A.; Hall, C.E. Electron microscope observations of the trichocysts and cilia of Paramecium. Biol. Bull. 1946, 91, 141–144. [Google Scholar] [CrossRef]
- Manton, I.; Clarke, B. Electron Microscope Observations on the Spermatozoid of Fucus. Nature 1950, 166, 973–974. [Google Scholar] [CrossRef]
- Fawcett, D.W.; Porter, K.R. A study of the fine structure of ciliated epithelia. J. Morphol. 1954, 94, 221–281. [Google Scholar] [CrossRef]
- Bloodgood, R.A. From Central to Rudimentary to Primary: The History of an Underappreciated Organelle Whose Time Has Come.The Primary Cilium. In Methods in Cell Biology; Academic Press: Cambridge, MA, USA, 2009. [Google Scholar] [CrossRef]
- Heidemann, S.R.; Kirschner, M.W. Aster formation in eggs of Xenopus laevis. Induction by isolated basal bodies. J. Cell Biol. 1975, 67, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Afzelius, B. Electron microscopy of the sperm tail; results obtained with a new fixative. J. Biophys. Biochem. Cytol. 1959, 5, 269–278. [Google Scholar] [CrossRef]
- Gibbons, I.R.; Grimstone, A.V. On flagellar structure in certain flagellates. J. Biophys. Biochem. Cytol. 1960, 7, 697–716. [Google Scholar] [CrossRef] [PubMed]
- Ringo, D.L. Flagellar motion and fine structure of the flagellar apparatus in Chlamydomonas. J. Cell Biol. 1967, 33, 543–571. [Google Scholar] [CrossRef] [PubMed]
- Satir, P. STUDIES ON CILIA: II. Examination of the Distal Region of the Ciliary Shaft and the Role of the Filaments in Motility. J. Cell Biol. 1965, 26, 805–834. [Google Scholar] [CrossRef]
- Bessis, M.; Nomarski, G.; Thiery, J.P.; Breton-Gorius, J. [Sickling of erythrocytes studied with polarized light & electron microscopes. II. Erythrocyte internal structure; comparison with intra-erythrocytic crystals]. Rev. Hematol. 13, 249–270.
- Ledbetter, M.C.; Porter, K.R. A “Microtubule” In Plant Cell Fine Structure. J. Cell Biol. 1963, 19, 239–250. [Google Scholar] [CrossRef]
- Slautterback, D.B. Cytoplasmic Microtubules. I. Hydra. J. Cell Biol. 1963, 18, 367–388. [Google Scholar] [CrossRef]
- Watson, M.R.; Hopkins, J.M. Isolated cilia from Tetrahymena pyriformis. Exp. Cell Res. 1962, 28, 280–295. [Google Scholar] [CrossRef]
- Pease, D.C. The Ultrastructure Of Flagellar Fibrils. J. Cell Biol. 1963, 18, 313–326. [Google Scholar] [CrossRef]
- Ledbetter, M.C.; Porter, K.R. Morphology of Microtubules of Plant Cell. Science 1964, 144, 872–874. [Google Scholar] [CrossRef] [PubMed]
- Stephens, R.E.; Renaud, F.L.; Gibbons, I.R.; Stevens, R.E. Guanine nucleotide associated with the protein of the outer fibers of flagella and cilia. Science 1967, 156, 1606–1608. [Google Scholar] [CrossRef]
- Borisy, G.G.; Taylor, E.W. The mechanism of action of colchicine. Binding of colchincine-3H to cellular protein. J. Cell Biol. 1967, 34, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Shelanski, M.L.; Taylor, E.W. Isolation of a protein subunit from microtubules. J. Cell Biol. 1967, 34, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, I.R.; Rowe, A.J. Dynein: A Protein with Adenosine Triphosphatase Activity from Cilia. Science 1965, 149, 424–426. [Google Scholar] [CrossRef] [PubMed]
- Munger, B.L. A light and electron microscopic study of cellular differentiation in the pancreatic islets of the mouse. Am. J. Anat. 1958, 103, 275–311. [Google Scholar] [CrossRef] [PubMed]
- Currie, A.R.; Wheatley, D.N. Cilia of a Distinctive Structure (9+0) in Endocrine and other Tissues. Postgrad Med. J. 1966, 42, 403. [Google Scholar] [CrossRef]
- Wheatley, D.N. Cilia in cell-cultured fibroblasts. I. On their occurrence and relative frequencies in primary cultures and established cell lines. J. Anat. 1969, 105, 351–362. [Google Scholar]
- Sorokin, S.P. Reconstructions of centriole formation and ciliogenesis in mammalian lungs. J. Cell Sci. 1968, 3, 207–230. [Google Scholar]
- Poole, C.A.; Flint, M.H.; Beaumont, B.W. Analysis of the morphology and function of primary cilia in connective tissues: A cellular cybernetic probe? Cell Motil. 1985, 5, 175–193. [Google Scholar] [CrossRef]
- Wheatley, D.N. The primary cilium—Once a “rudimentary” organelle that is now a ubiquitous sensory cellular structure involved in many pathological disorders. J. Cell Commun. Signal. 2018, 12, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Kozminski, K.G.; Johnson, K.A.; Forscher, P.; Rosenbaum, J.L. A motility in the eukaryotic flagellum unrelated to flagellar beating. Proc. Natl. Acad. Sci. USA 1993, 90, 5519–5523. [Google Scholar] [CrossRef]
- Cole, D.G.; Diener, D.R.; Himelblau, A.L.; Beech, P.L.; Fuster, J.C.; Rosenbaum, J.L. Chlamydomonas kinesin-II-dependent intraflagellar transport (IFT): IFT particles contain proteins required for ciliary assembly in Caenorhabditis elegans sensory neurons. J. Cell Biol. 1998, 141, 993–1008. [Google Scholar] [CrossRef]
- Pazour, G.J.; Wilkerson, C.G.; Witman, G.B. A dynein light chain is essential for the retrograde particle movement of intraflagellar transport (IFT). J. Cell Biol. 1998, 141, 979–992. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, S.; Tanaka, Y.; Okada, Y.; Takeda, S.; Harada, A.; Kanai, Y.; Kido, M.; Hirokawa, N. Randomization of left-right asymmetry due to loss of nodal cilia generating leftward flow of extraembryonic fluid in mice lacking KIF3B motor protein. Cell 1998, 95, 829–837. [Google Scholar] [CrossRef]
- Pazour, G.J.; Dickert, B.L.; Vucica, Y.; Seeley, E.S.; Rosenbaum, J.L.; Witman, G.B.; Cole, D.G. Chlamydomonas IFT88 and its mouse homologue, polycystic kidney disease gene tg737, are required for assembly of cilia and flagella. J. Cell Biol. 2000, 151, 709–718. [Google Scholar] [CrossRef] [PubMed]
- Cavalier-Smith, T. The phagotrophic origin of eukaryotes and phylogenetic classification of Protozoa. Int. J. Syst. Evol. Microbiol. 2002, 52, 297–354. [Google Scholar] [CrossRef] [PubMed]
- Carvalho-Santos, Z.; Azimzadeh, J.; Pereira-Leal, J.B.; Bettencourt-Dias, M. Tracing the origins of centrioles, cilia, and flagella. J. Cell Biol. 2011, 194, 165–175. [Google Scholar] [CrossRef]
- Mitchell, D.R. The Evolution of Eukaryotic Cilia and Flagella as Motile and Sensory Organelles. In Eukaryotic Membranes and Cytoskeleton; Springer New York: New York, NY, USA, 2007; pp. 130–140. [Google Scholar]
- Sharma, N.; Kosan, Z.A.; Stallworth, J.E.; Berbari, N.F.; Yoder, B.K. Soluble levels of cytosolic tubulin regulate ciliary length control. Mol. Biol. Cell 2011, 22, 806–816. [Google Scholar] [CrossRef]
- Nicastro, D.; Schwartz, C.; Pierson, J.; Gaudette, R.; Porter, M.E.; McIntosh, J.R. The Molecular Architecture of Axonemes Revealed by Cryoelectron Tomography. Science 2006, 313, 944–948. [Google Scholar] [CrossRef]
- Goetz, S.C.; Anderson, K.V. The primary cilium: A signalling centre during vertebrate development. Nat. Rev. Genet. 2010, 11, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Satir, P.; Christensen, S.T. Overview of Structure and Function of Mammalian Cilia. Annu Rev. Physiol. 2007, 69, 377–400. [Google Scholar] [CrossRef] [PubMed]
- Wheway, G.; Nazlamova, L.; Hancock, J.T. Signaling through the Primary Cilium. Front. Cell Dev. Biol. 2018, 6, 8. [Google Scholar] [CrossRef] [PubMed]
- Bloodgood, R.A. Sensory reception is an attribute of both primary cilia and motile cilia. J. Cell Sci. 2010, 123, 505–509. [Google Scholar] [CrossRef] [PubMed]
- Spirostomum, I.; Author, S.; Jennings, H.S. Studies on Reactions to Stimuli in Unicellular Organisms. III Reactions to Localized Stimuli. Am. Nat. 1899, 33, 373–389. [Google Scholar]
- Braun, D.A.; Hildebrandt, F. Ciliopathies. Cold Spring Harb. Perspect. Biol. 2017, 9, a028191. [Google Scholar] [CrossRef] [PubMed]
- Reiter, J.F.; Leroux, M.R. Genes and molecular pathways underpinning ciliopathies. Nat. Rev. Mol. Cell Biol. 2017, 18, 533–547. [Google Scholar] [CrossRef]
- Jana, S.C.; Mendonça, S.; Machado, P.; Werner, S.; Rocha, J.; Pereira, A.; Maiato, H.; Bettencourt-Dias, M. Differential regulation of transition zone and centriole proteins contributes to ciliary base diversity. Nat. Cell Biol. 2018, 20, 928–941. [Google Scholar] [CrossRef]
- Nechipurenko, I.V.; Berciu, C.; Sengupta, P.; Nicastro, D. Centriolar remodeling underlies basal body maturation during ciliogenesis in Caenorhabditis elegans. Elife 2017, 6. [Google Scholar] [CrossRef]
- Shahid, U.; Singh, P. Emerging Picture of Deuterosome-Dependent Centriole Amplification in MCCs. Cells 2018, 7, 152. [Google Scholar] [CrossRef]
- Nguyen, Q.; Liu, Z.; Nanjundappa, R.; Megherbi, A.; Delgehyr, N.; Ouyang, H.; Zock, L.; Coyaud, E.; Laurent, E.; Dell, S.; et al. Super-resolution Molecular Map of Basal Foot Reveals Novel Cilium in Airway Multiciliated Cells. bioRxiv 2018, 487330. [Google Scholar] [CrossRef]
- Chen, J.V.; Kao, L.-R.; Jana, S.C.; Sivan-Loukianova, E.; Mendonça, S.; Cabrera, O.A.; Singh, P.; Cabernard, C.; Eberl, D.F.; Bettencourt-Dias, M.; Megraw, T.L.; et al. Rootletin organizes the ciliary rootlet to achieve neuron sensory function in Drosophila. J. Cell Biol. 2015, 211, 435–453. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Gao, J.; Adamian, M.; Wen, X.-H.; Pawlyk, B.; Zhang, L.; Sanderson, M.J.; Zuo, J.; Makino, C.L.; Li, T. The Ciliary Rootlet Maintains Long-Term Stability of Sensory Cilia. Mol. Cell Biol. 2005, 25, 4129–4137. [Google Scholar] [CrossRef] [PubMed]
- Mohan, S.; Timbers, T.A.; Kennedy, J.; Blacque, O.E.; Leroux, M.R. Striated Rootlet and Nonfilamentous Forms of Rootletin Maintain Ciliary Function. Curr. Biol. 2013, 23, 2016–2022. [Google Scholar] [CrossRef]
- Gilliam, J.C.; Chang, J.T.; Sandoval, I.M.; Zhang, Y.; Li, T.; Pittler, S.J.; Chiu, W.; Wensel, T.G. Three-Dimensional Architecture of the Rod Sensory Cilium and Its Disruption in Retinal Neurodegeneration. Cell 2012, 151, 1029–1041. [Google Scholar] [CrossRef]
- Dentler, W.L. Cilia and Flagella. In Cytology and Cell Physiology; Elsevier: Amsterdam, The Netherlands, 1987; pp. 391–456. [Google Scholar] [CrossRef]
- Clare, D.K.; Magescas, J.; Piolot, T.; Dumoux, M.; Vesque, C.; Pichard, E.; Dang, T.; Poirier, F. Basal foot MTOC organizes pillar MTs required for coordination of beating cilia. Nat. Commun. 2014, 5, 4888. [Google Scholar] [CrossRef] [PubMed]
- Kunimoto, K.; Yamazaki, Y.; Nishida, T.; Shinohara, K.; Ishikawa, H.; Hasegawa, T.; Okanoue, T.; Hamada, H.; Noda, T.; Tamura, A.; et al. Coordinated Ciliary Beating Requires Odf2-Mediated Polarization of Basal Bodies via Basal Feet. Cell 2012, 148, 189–200. [Google Scholar] [CrossRef]
- Mazo, G.; Soplop, N.; Wang, W.-J.; Uryu, K.; Tsou, M.-F.B. Spatial Control of Primary Ciliogenesis by Subdistal Appendages Alters Sensation-Associated Properties of Cilia. Dev. Cell. 2016, 39, 424–437. [Google Scholar] [CrossRef]
- Gonçalves, J.; Pelletier, L. The Ciliary Transition Zone: Finding the Pieces and Assembling the Gate. Mol. Cell. 2017, 40, 243–253. [Google Scholar] [CrossRef]
- Gibbons, I.R. The relationship between the fine structure and direction of beat in gill cilia of a lamellibranch mollusc. J. Biophys. Biochem. Cytol. 1961, 11, 179–205. [Google Scholar] [CrossRef]
- Gibbons, I.R. Cilia and flagella of eukaryotes. J. Cell Biol. 1981, 91, 107s–124s. [Google Scholar] [CrossRef] [PubMed]
- Inaba, K. Sperm flagella: Comparative and phylogenetic perspectives of protein components. Mol. Hum. Reprod. 2011, 17, 524–538. [Google Scholar] [CrossRef] [PubMed]
- Kramer-Zucker, A.G.; Olale, F.; Haycraft, C.J.; Yoder, B.K.; Schier, A.F.; Drummond, I.A. Cilia-driven fluid flow in the zebrafish pronephros, brain and Kupffer’s vesicle is required for normal organogenesis. Development 2005, 132, 1907–1921. [Google Scholar] [CrossRef] [PubMed]
- Bird, S.D.; Legge, M.; Walker, R.J. Cultured peritoneal mesothelial cells exhibit apical primary cilia. Cell Biol. Int. 2004, 28, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Moran, D.T.; Rowley, J.C.; Jafek, B.W.; Lovell, M.A. The fine structure of the olfactory mucosa in man. J. Neurocytol. 1982, 11, 721–746. [Google Scholar] [CrossRef] [PubMed]
- Babu, D.; Roy, S. Left-right asymmetry: Cilia stir up new surprises in the node. Open Biol. 2013, 3, 130052. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, S. Motility of the 6+0 flagellum of Lecudina tuzetae. Cell Motil. Cytoskeleton 1982, 2, 369–383. [Google Scholar] [CrossRef]
- Afzelius, B. Cilia-related diseases. J. Pathol. 2004, 204, 470–477. [Google Scholar] [CrossRef]
- Okada, Y.; Takeda, S.; Tanaka, Y.; Belmonte, J.-C.I.; Hirokawa, N. Mechanism of Nodal Flow: A Conserved Symmetry Breaking Event in Left-Right Axis Determination. Cell 2005, 121, 633–644. [Google Scholar] [CrossRef]
- Christensen, S.T.; Guerra, C.; Wada, Y.; Valentin, T.; Angeletti, R.H.; Satir, P.; Hamasaki, T. A Regulatory Light Chain of Ciliary Outer Arm Dynein in Tetrahymena thermophila. J. Biol. Chem. 2001, 276, 20048–20054. [Google Scholar] [CrossRef]
- Habermacher, G.; Sale, W.S. Regulation of flagellar dynein by phosphorylation of a 138-kD inner arm dynein intermediate chain. J. Cell Biol. 1997, 136, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Nicastro, D.; McIntosh, J.R.; Baumeister, W. 3D structure of eukaryotic flagella in a quiescent state revealed by cryo-electron tomography. Proc. Natl. Acad. Sci. USA 2005, 102, 15889–15894. [Google Scholar] [CrossRef] [PubMed]
- Viswanadha, R.; Sale, W.S.; Porter, M.E. Ciliary Motility: Regulation of Axonemal Dynein Motors. Cold Spring Harb. Perspect. Biol. 2017, 9, a018325. [Google Scholar] [CrossRef]
- Inaba, K. Molecular basis of sperm flagellar axonemes: Structural and evolutionary aspects. Ann. N. Y. Acad. Sci. 2007, 1101, 506–526. [Google Scholar] [CrossRef]
- Kollmar, M. Fine-Tuning Motile Cilia and Flagella: Evolution of the Dynein Motor Proteins from Plants to Humans at High Resolution. Mol. Biol. Evol. 2016, 33, 3249–3267. [Google Scholar] [CrossRef] [PubMed]
- Chasey, D. Further observations on the ultrastructure of cilia from Tetrahymena pyriformis. Exp. Cell Res. 1972, 74, 471–479. [Google Scholar] [CrossRef]
- Olson, G.E.; Linck, R.W. Observations of the structural components of flagellar axonemes and central pair microtubules from rat sperm. J. Ultrastruct. Res. 1977, 61, 21–43. [Google Scholar] [CrossRef]
- Hopkins, J.M. Subsidiary Components of the Flagella of Chlamydomonas Reinhardii. J. Cell Sci. 1970, 7. [Google Scholar]
- Warner, F.D. New observations on flagellar fine structure. The relationship between matrix structure and the microtubule component of the axoneme. J. Cell Biol. 1970, 47, 159–182. [Google Scholar] [CrossRef]
- Barber, C.F.; Heuser, T.; Carbajal-González, B.I.; Botchkarev, V.V.; Nicastro, D. Three-dimensional structure of the radial spokes reveals heterogeneity and interactions with dyneins in Chlamydomonas flagella. Mol. Biol. Cell 2012, 23, 111–120. [Google Scholar] [CrossRef]
- Pigino, G.; Bui, K.H.; Maheshwari, A.; Lupetti, P.; Diener, D.; Ishikawa, T. Cryoelectron tomography of radial spokes in cilia and flagella. J. Cell Biol. 2011, 195, 673–687. [Google Scholar] [CrossRef] [PubMed]
- Pigino, G.; Ishikawa, T. Axonemal radial spokes: 3D structure, function and assembly. Bioarchitecture 2012, 2, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Liu, Y.; Yang, P. Radial Spokes—A Snapshot of the Motility Regulation, Assembly, and Evolution of Cilia and Flagella. Cold Spring Harb. Perspect. Biol. 2017, 9, a028126. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Morsci, N.; Nguyen, K.C.Q.; Rizvi, A.; Rongo, C.; Hall, D.H.; Barr, M.M. Cell-Specific α-Tubulin Isotype Regulates Ciliary Microtubule Ultrastructure, Intraflagellar Transport, and Extracellular Vesicle Biology. Curr. Biol. 2017, 27, 968–980. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.M. Insect sperm: Their structure and morphogenesis. J. Cell Biol. 1970, 44, 243–277. [Google Scholar] [CrossRef] [PubMed]
- Dallai, R.; Lupetti, P.; Mencarelli, C. Unusual axonemes of hexapod spermatozoa. Int. Rev. Cytol. 2006, 254, 45–99. [Google Scholar] [CrossRef] [PubMed]
- Schrevel, J.; Besse, C. [A functional flagella with a 6 + 0 pattern]. J. Cell Biol. 1975, 66, 492–507. [Google Scholar] [CrossRef]
- Prensier, G.; Vivier, E.; Goldstein, S.; Schrével, J. Motile flagellum with a “3 + 0” ultrastructure. Science 1980, 207, 1493–1494. [Google Scholar] [CrossRef]
- Gluenz, E.; Höög, J.L.; Smith, A.E.; Dawe, H.R.; Shaw, M.K.; Gull, K. Beyond 9+0: Noncanonical axoneme structures characterize sensory cilia from protists to humans. FASEB J. 2010, 24, 3117–3121. [Google Scholar] [CrossRef]
- Van Deurs, B. Axonemal 12 + 0 pattern in the flagellum of the motile spermatozoon of Nymphon leptocheles. J. Ultrastruct. Res. 1973, 42, 594–598. [Google Scholar] [CrossRef]
- Baccetti, B.; Dallai, R.; Fratello, B. The spermatozoon of arthropoda. XXII. The 12+0’, 14+0’ or aflagellate sperm of protura. J. Cell Sci. 1973, 13, 321–335. [Google Scholar] [PubMed]
- Dahl, H.A. Fine structure of cilia in rat cerebral cortex. Zeitschrift für Zellforsch und Mikroskopische Anat 1963, 60, 369–386. [Google Scholar] [CrossRef]
- Fawcett, D.W.; Iton, S. The fine structure of bat spermatozoa. Am. J. Anat. 1965, 116, 567–609. [Google Scholar] [CrossRef] [PubMed]
- Fawcett, D.W. The anatomy of the mammalian spermatozoon with particular reference to the guinea pig. Z Zellforsch Mikrosk Anat. 1965, 67, 279–296. [Google Scholar] [CrossRef] [PubMed]
- Silveira, M.; Porter, K.R. The spermatozoids of Flatworms and their microtubular systems. Protoplasma 1964, 59, 240–265. [Google Scholar] [CrossRef]
- Roth, L.E.; Shigenaka, Y. The structure and formation of cilia and filaments in rumen protozoa. J. Cell Biol. 1964, 20, 249–270. [Google Scholar] [CrossRef] [PubMed]
- Roth, L.E. Aspects of ciliary fine structure in Euplotes patella. J. Biophys. Biochem. Cytol. 1956, 2, 235–240. [Google Scholar] [CrossRef]
- Koch, W.J. Studies of the Motile Cells of Chytrids. I. Electron Microscope Observations of the Flagellum, Blepharoplast and Rhizoplast. Am. J. Bot. 1956, 43, 811. [Google Scholar] [CrossRef]
- Manton, I. Electron microscopical observations on a very small flagellate: The problem of Chromulina pusilla Butcher. J. Mar. Biol. Assoc. UK 1959, 38, 319. [Google Scholar] [CrossRef]
- Tyler, S. An Adhesive Function for Modified Cilia in an Interstitial Turbellarian. Acta Zool. 1973, 54, 139–151. [Google Scholar] [CrossRef]
- Dentler, W.L. Structures linking the tips of ciliary and flagellar microtubules to the membrane. J. Cell Sci. 1980, 42, 207–220. [Google Scholar]
- Satir, P. Studies on cilia. 3. Further studies on the cilium tip and a "sliding filament" model of ciliary motility. J. Cell Biol. 1968, 39, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Dentler, W.L. Microtubule-Membrane Interactions in Cilia and Flagella. Int. Rev. Cytol. 1981, 72, 1–47. [Google Scholar] [CrossRef] [PubMed]
- Dentler, W.L.; Rosenbaum, J.L. Flagellar elongation and shortening in Chlamydomonas. III. structures attached to the tips of flagellar microtubules and their relationship to the directionality of flagellar microtubule assembly. J. Cell Biol. 1977, 74, 747–759. [Google Scholar] [CrossRef]
- Dentler, W.L. Attachment of the cap to the central microtubules of Tetrahymena cilia. J. Cell Sci. 1984, 66, 167–173. [Google Scholar] [PubMed]
- Kuhn, C.; Engleman, W. The structure of the tips of mammalian respiratory cilia. Cell Tissue Res. 1978, 186, 491–498. [Google Scholar] [CrossRef]
- Tamm, S.L.; Tamm, S. Visualization Of Changes In Ciliary Tip Configuration Caused By Sliding Displacement Of Microtubules In Macrocilia Of The Ctenophore Beroe. J. Cell Sci. 1985, 79, 161–179. [Google Scholar] [PubMed]
- Mukhopadhyay, S.; Lu, Y.; Shaham, S.; Sengupta, P. Sensory Signaling-Dependent Remodeling of Olfactory Cilia Architecture in C. elegans. Dev. Cell. 2008, 14, 762–774. [Google Scholar] [CrossRef]
- Varga, V.; Moreira-Leite, F.; Portman, N.; Gull, K. Protein diversity in discrete structures at the distal tip of the trypanosome flagellum. Proc. Natl. Acad. Sci. USA 2017, 114, E6546–E6555. [Google Scholar] [CrossRef]
- Woolley, D.; Gadelha, C.; Gull, K. Evidence for a sliding-resistance at the tip of the trypanosome flagellum. Cell Motil. Cytoskeleton 2006, 63, 741–746. [Google Scholar] [CrossRef]
- Dirksen, E.R.; Satir, P. Ciliary activity in the mouse oviduct as studied by transmission and scanning electron microscopy. Tissue Cell. 1972, 4, 389–403. [Google Scholar] [CrossRef]
- Jeffery, P.K.; Reid, L. New observations of rat airway epithelium: A quantitative and electron microscopic study. J. Anat. 1975, 120, 295–320. [Google Scholar] [PubMed]
- Cordier, A.C. Ultrastructure of the cilia of thymic cysts in “nude” mice. Anat Rec. 1975, 181, 227–249. [Google Scholar] [CrossRef] [PubMed]
- Dentler, W.L. Linkages between Microtubules and Membranes in Cilia and Flagella. In Ciliary and Flagellar Membranes; Springer US: Boston, MA, USA, 1990; pp. 31–64. [Google Scholar]
- Chailley, B.; N’Diaye, A.; Boisvieux-Ulrich, E.; Sandoz, D. Comparative study of the distribution of fuzzy coat, lectin receptors, and intramembrane particles of the ciliary membrane. Eur. J. Cell Biol. 1981, 25, 300–307. [Google Scholar] [PubMed]
- Tyler, S. Distinctive features of cilia in metazoans and their significance for systematics. Tissue Cell. 1979, 11, 385–400. [Google Scholar] [CrossRef]
- Dentler, W.L.; LeCluyse, E.L. Microtubule capping structures at the tips of tracheal cilia: Evidence for their firm attachment during ciliary bend formation and the restriction of microtubule sliding. Cell Motil. 1982, 2, 549–572. [Google Scholar] [CrossRef] [PubMed]
- LeCluyse, E.L.; Dentler, W.L. Asymmetrical microtubule capping structures in frog palate cilia. J. Ultrastruct. Res. 1984, 86, 75–85. [Google Scholar] [CrossRef]
- Owen, G.; McCrae, J.M. Sensory Cell/Gland Cell Complexes Associated with the Pallial Tentacles of the Bivalve Lima hians (Gmelin), with a Note on Specialized Cilia on the Pallial Curtains. Phil. Trans. R. Soc. B 1979, 287, 45–62. [Google Scholar] [CrossRef]
- Toh, Y.; Yokohari, F. Structure of the antennal chordotonal sensilla of the American cockroach. J. Ultrastruct. Res. 1985, 90, 124–134. [Google Scholar] [CrossRef]
- Pedersen, L.B.; Schrøder, J.M.; Satir, P.; Christensen, S.T. The Ciliary Cytoskeleton. In Comprehensive Physiology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012; pp. 779–803. [Google Scholar]
- Goodenough, U.W. Tipping of flagellar agglutinins by gametes of Chlamydomonas reinhardtii. Cell Motil. Cytoskeleton 1993, 25, 179–189. [Google Scholar] [CrossRef]
- Tuomanen, E. Adjuncts to the therapy of bacterial meningitis. Pediatr. Infect. Dis. J. 1990, 9. [Google Scholar] [CrossRef]
- Anderson, R.G.; Hein, C.E. Distribution of anionic sites on the oviduct ciliary membrane. J. Cell Biol. 1977, 72, 482–492. [Google Scholar] [CrossRef] [PubMed]
- Aiello, E.; Sleigh, M. Ciliary function of the frog oro-pharyngeal epithelium. Cell Tissue Res. 1977, 178, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.A.; Rosenbaum, J.L. Polarity of flagellar assembly in Chlamydomonas. J. Cell Biol. 1992, 119, 1605–1611. [Google Scholar] [CrossRef] [PubMed]
- Witman, G.B. The site of in vivo assembly of flagellar microtubules. Ann. N. Y. Acad. Sci. 1975, 253, 178–191. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, J.L.; Carlson, K. Cilia regeneration in Tetrahymena and its inhibition by colchicine. J. Cell Biol. 1969, 40, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, J.L.; Moulder, J.E.; Ringo, D.L. Flagellar elongation and shortening in Chlamydomonas. The use of cycloheximide and colchicine to study the synthesis and assembly of flagellar proteins. J. Cell Biol. 1969, 41, 600–619. [Google Scholar] [CrossRef]
- Johnson, U.G.; Porter, K.R. Fine structure of cell division in Chlamydomonas reinhardi. Basal bodies and microtubules. J. Cell Biol. 1968, 38, 403–425. [Google Scholar] [CrossRef]
- Marshall, W.F.; Rosenbaum, J.L. Intraflagellar transport balances continuous turnover of outer doublet microtubules. J. Cell Biol. 2001, 155, 405–414. [Google Scholar] [CrossRef]
- Hao, L.; Thein, M.; Brust-Mascher, I.; Civelekoglu-Scholey, G.; Lu, Y.; Acar, S.; Prevo, B.; Shaham, S.; Scholey, J.M. Intraflagellar transport delivers tubulin isotypes to sensory cilium middle and distal segments. Nat. Cell Biol. 2011, 13, 790–798. [Google Scholar] [CrossRef]
- Seixas, C.; Cruto, T.; Tavares, A.; Gaertig, J.; Soares, H. CCTα and CCTδ Chaperonin Subunits Are Essential and Required for Cilia Assembly and Maintenance in Tetrahymena. PLoS ONE 2010, 5, e10704. [Google Scholar] [CrossRef] [PubMed]
- Rich, D.R.; Clark, A.L. Chondrocyte primary cilia shorten in response to osmotic challenge and are sites for endocytosis. Osteoarthr. Cartil. 2012, 20, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Craft, J.M.; Harris, J.A.; Hyman, S.; Kner, P.; Lechtreck, K.F. Tubulin transport by IFT is upregulated during ciliary growth by a cilium-autonomous mechanism. J. Cell Biol. 2015, 208, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Prevo, B.; Mangeol, P.; Oswald, F.; Scholey, J.M.; Peterman, E.J.G. Functional differentiation of cooperating kinesin-2 motors orchestrates cargo import and transport in C. elegans cilia. Nat. Cell Biol. 2015, 17, 1536–1545. [Google Scholar] [CrossRef] [PubMed]
- Kozminski, K.G.; Beech, P.L.; Rosenbaum, J.L. The Chlamydomonas kinesin-like protein FLA10 is involved in motility associated with the flagellar membrane. J. Cell Biol. 1995, 131, 1517–1527. [Google Scholar] [CrossRef] [PubMed]
- Snow, J.J.; Ou, G.; Gunnarson, A.L.; Walker, M.R.S.; Zhou, H.M.; Brust-Mascher, I.; Scholey, J.M. Two anterograde intraflagellar transport motors cooperate to build sensory cilia on C. elegans neurons. Nat. Cell Biol. 2004, 6, 1109–1113. [Google Scholar] [CrossRef]
- Pazour, G.J.; Dickert, B.L.; Witman, G.B. The DHC1b (DHC2) isoform of cytoplasmic dynein is required for flagellar assembly. J. Cell Biol. 1999, 144, 473–481. [Google Scholar] [CrossRef]
- Porter, M.E.; Bower, R.; Knott, J.A.; Byrd, P.; Dentler, W. Cytoplasmic Dynein Heavy Chain 1b Is Required for Flagellar Assembly in Chlamydomonas. Mol. Biol. Cell 1999, 10, 693–712. [Google Scholar] [CrossRef]
- Signor, D.; Wedaman, K.P.; Orozco, J.T.; Dwyer, N.D.; Bargmann, C.I.; Rose, L.S.; Signor, D.; Scholey, J.M. Role of a class DHC1b dynein in retrograde transport of IFT motors and IFT raft particles along cilia, but not dendrites, in chemosensory neurons of living Caenorhabditis elegans. J. Cell Biol. 1999, 147, 519–530. [Google Scholar] [CrossRef]
- Chien, A.; Shih, S.M.; Bower, R.; Tritschler, D.; Porter, M.E.; Yildiz, A. Dynamics of the IFT machinery at the ciliary tip. Elife 2017, 6. [Google Scholar] [CrossRef]
- Pedersen, L.B.; Rosenbaum, J.L. Chapter Two Intraflagellar Transport (IFT). Curr. Top. Dev. Biol. 2008, 85, 23–61. [Google Scholar] [CrossRef] [PubMed]
- Portman, R.W.; LeCluyse, E.L.; Dentler, W.L. Development of microtubule capping structures in ciliated epithelial cells. J. Cell Sci. 1987, 87, 85–94. [Google Scholar] [PubMed]
- Fok, A.K.; Wang, H.; Katayama, A.; Aihara, M.S.; Allen, R.D. 22S axonemal dynein is preassembled and functional prior to being transported to and attached on the axonemes. Cell Motil. Cytoskeleton 1994, 29, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Diener, D.R.; Geimer, S.; Cole, D.G.; Rosenbaum, J.L. Intraflagellar transport (IFT) cargo. J. Cell Biol. 2004, 164, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Seixas, C.; Gonçalves, J.; Melo, L.V.; Soares, H. Tetrahymena Cilia Cap is Built in a Multi-step Process: A Study by Atomic Force Microscopy. Protist 2017, 168, 697–717. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, M.J.; Phetruen, T.; Fisher, R.L.; Chen, K.; Pentecost, B.T.; Gomez, G.; Ounjai, P.; Sui, H. The Developmental Process of the Growing Motile Ciliary Tip Region. Sci. Rep. 2018, 8, 7977. [Google Scholar] [CrossRef] [PubMed]
- Dentler, W.L.; LeCluyse, E.L. The effects of structures attached to the tips of tracheal ciliary microtubules on the nucleation of microtubule assembly in vitro. Prog. Clin. Biol. Res. 1982, 80, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Péchart, I.; Kann, M.L.; Levilliers, N.; Bré, M.H.; Fouquet, J.P. Composition and organization of tubulin isoforms reveals a variety of axonemal models. Biol. cell. 1999, 91, 685–697. [Google Scholar] [CrossRef]
- Jensen-Smith, H.C.; Ludueña, R.F.; Hallworth, R. Requirement for the βI and βIV tubulin isotypes in mammalian cilia. Cell Motil. Cytoskeleton 2003, 55, 213–220. [Google Scholar] [CrossRef]
- Bosch Grau, M.; Gonzalez Curto, G.; Rocha, C.; Magiera, M.M.; Marques Sousa, P.; Giordano, T.; Spassky, N.; Janke, C. Tubulin glycylases and glutamylases have distinct functions in stabilization and motility of ependymal cilia. J. Cell Biol. 2013, 202, 441–451. [Google Scholar] [CrossRef]
- Fisch, C.; Dupuis-Williams, P. Ultrastructure of Cilia and Flagella - Back to the Future! Biol. Cell. 2011, 103, 249–270. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.M.; Wang, W.; Balczon, R.; Dentler, W.L. Ciliary Microtubule Capping Structures Contain a Mammalian Kinetochore Antigen. J. Cell Biol. 1990, 110, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Bloch, M.A.; Johnson, K.A. Identification of a molecular chaperone in the eukaryotic flagellum and its localization to the site of microtubule assembly. J. Cell Sci. 1995, 108, 3541–3545. [Google Scholar] [PubMed]
- Piperno, G.; Mead, K. Transport of a novel complex in the cytoplasmic matrix of Chlamydomonas flagella. Proc. Natl. Acad. Sci. USA 1997, 94, 4457–4462. [Google Scholar] [CrossRef] [PubMed]
- Seixas, C.; Casalou, C.; Melo, L.V.; Nolasco, S.; Brogueira, P.; Soares, H. Subunits of the chaperonin CCT are associated with Tetrahymena microtubule structures and are involved in cilia biogenesis. Exp. Cell Res. 2003, 290, 303–321. [Google Scholar] [CrossRef]
- Stephens, R.E.; Lemieux, N.A. Molecular chaperones in cilia and flagella: Implications for protein turnover. Cell Motil. Cytoskeleton 1999, 44, 274–283. [Google Scholar] [CrossRef]
- Luo, W.; Ruba, A.; Takao, D.; Zweifel, L.P.; Lim, R.Y.H.; Verhey, K.J.; Yang, W. Axonemal Lumen Dominates Cytosolic Protein Diffusion inside the Primary Cilium. Sci. Rep. 2017, 7, 15793. [Google Scholar] [CrossRef] [PubMed]
- Mirvis, M.; Stearns, T.; James Nelson, W. Cilium structure, assembly, and disassembly regulated by the cytoskeleton. Biochem. J. 2018, 475, 2329–2353. [Google Scholar] [CrossRef]
- Dawson, S.C.; Sagolla, M.S.; Mancuso, J.J.; Woessner, D.J.; House, S.A.; Fritz-Laylin, L.; Cande, W.Z. Kinesin-13 Regulates Flagellar, Interphase, and Mitotic Microtubule Dynamics in Giardia intestinalis. Eukaryot. Cell 2007, 6, 2354–2364. [Google Scholar] [CrossRef]
- Pedersen, L.B.; Geimer, S.; Sloboda, R.D.; Rosenbaum, J.L. The Microtubule plus end-tracking protein EB1 is localized to the flagellar tip and basal bodies in Chlamydomonas reinhardtii. Curr. Biol. 2003, 13, 1969–1974. [Google Scholar] [CrossRef]
- Pedersen, L.B.; Miller, M.S.; Geimer, S.; Leitch, J.M.; Rosenbaum, J.L.; Cole, D.G. Chlamydomonas IFT172 Is Encoded by FLA11, Interacts with CrEB1, and Regulates IFT at the Flagellar Tip. Curr. Biol. 2005, 15, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Brooks, E.R.; Wallingford, J.B. Control of vertebrate intraflagellar transport by the planar cell polarity effector Fuz. J. Cell Biol. 2012, 198, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Schrøder, J.M.; Larsen, J.; Komarova, Y.; Akhmanova, A.; Thorsteinsson, R.I.; Grigoriev, I.; Manguso, R.; Christensen, S.T.; Pedersen, S.F.; Geimer, S.; et al. EB1 and EB3 promote cilia biogenesis by several centrosome-related mechanisms. J. Cell Sci. 2011, 124, 2539–2551. [Google Scholar] [CrossRef]
- Larsen, J.; Grigoriev, I.; Akhmanova, A.; Pedersen, L.B. Analysis of Microtubule Plus-End-Tracking Proteins in Cilia. Methods Rnzymol. 2013, 524, 105–122. [Google Scholar] [CrossRef]
- Liu, Q.; Tan, G.; Levenkova, N.; Li, T.; Pugh, E.N.; Rux, J.J.; Speicher, D.W.; Pierce, E.A. The Proteome of the Mouse Photoreceptor Sensory Cilium Complex. Mol. Cell Proteom. 2007, 6, 1299–1317. [Google Scholar] [CrossRef] [PubMed]
- Dougherty, G.W.; Adler, H.J.; Rzadzinska, A.; Gimona, M.; Tomita, Y.; Lattig, M.C.; Merritt, R.C.; Kachar, B. CLAMP, a novel microtubule-associated protein with EB-type calponin homology. Cell Motil. Cytoskeleton 2005, 62, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Gray, R.S.; Abitua, P.B.; Wlodarczyk, B.J.; Szabo-Rogers, H.L.; Blanchard, O.; Lee, I.; Weiss, G.S.; Liu, K.J.; Marcotte, E.M.; Wallingford, J.B.; et al. The planar cell polarity effector Fuz is essential for targeted membrane trafficking, ciliogenesis and mouse embryonic development. Nat. Cell Biol. 2009, 11, 1225–1232. [Google Scholar] [CrossRef] [PubMed]
- Slep, K.C. The role of TOG domains in microtubule plus end dynamics. Biochem. Soc. Trans. 2009, 37, 1002–1006. [Google Scholar] [CrossRef]
- Das, A.; Dickinson, D.J.; Wood, C.C.; Goldstein, B.; Slep, K.C. Crescerin uses a TOG domain array to regulate microtubules in the primary cilium. Mol. Biol. Cell 2015, 26, 4248–4264. [Google Scholar] [CrossRef]
- Louka, P.; Vasudevan, K.K.; Guha, M.; Joachimiak, E.; Wloga, D.; Tomasi, F.-X.; Baroud, C.N.; Dupuis-Williams, P.; Galati, D.F.; Pearson, C.G.; et al. Proteins that control the geometry of microtubules at the ends of cilia. J. Cell Biol. 2018, 217, 4298–4313. [Google Scholar] [CrossRef]
- Van De Weghe, J.C.; Rusterholz, T.D.S.; Latour, B.; Grout, M.E.; Aldinger, K.A.; Shaheen, R.; Dempsey, J.C.; Maddirevula, S.; Cheng, Y.-H.H.; Phelps, I.G.; et al. Mutations in ARMC9, which Encodes a Basal Body Protein, Cause Joubert Syndrome in Humans and Ciliopathy Phenotypes in Zebrafish. Am. J. Hum. Genet. 2017, 101, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Kar, A.; Phadke, S.R.; Das Bhowmik, A.; Dalal, A. Whole exome sequencing reveals a mutation in ARMC9 as a cause of mental retardation, ptosis, and polydactyly. Am. J. Med. Genet. Part A 2018, 176, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Breslow, D.K.; Hoogendoorn, S.; Kopp, A.R.; Morgens, D.W.; Vu, B.K.; Kennedy, M.C.; Han, K.; Li, A.; Hess, G.T.; Bassik, M.C.; et al. A CRISPR-based screen for Hedgehog signaling provides insights into ciliary function and ciliopathies. Nat. Genet. 2018, 50, 460–471. [Google Scholar] [CrossRef]
- Satish Tammana, T.V.; Tammana, D.; Diener, D.R.; Rosenbaum, J. Centrosomal protein CEP104 (Chlamydomonas FAP256) moves to the ciliary tip during ciliary assembly. J. Cell Sci. 2013, 126, 5018–5029. [Google Scholar] [CrossRef] [PubMed]
- Rezabkova, L.; Kraatz, S.H.W.; Akhmanova, A.; Steinmetz, M.O.; Kammerer, R.A. Biophysical and Structural Characterization of the Centriolar Protein Cep104 Interaction Network. J. Biol. Chem. 2016, 291, 18496–18504. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Toedt, G.; Montenegro Gouveia, S.; Davey, N.E.; Hua, S.; van der Vaart, B.; Grigoriev, I.; Larsen, J.; Pedersen, L.B.; Bezstarosti, K.; et al. A Proteome-wide Screen for Mammalian SxIP Motif-Containing Microtubule Plus-End Tracking Proteins. Curr. Biol. 2012, 22, 1800–1807. [Google Scholar] [CrossRef] [PubMed]
- Westermann, S.; Avila-Sakar, A.; Wang, H.-W.; Niederstrasser, H.; Wong, J.; Drubin, D.G.; Nogales, E.; Barnes, G. Formation of a Dynamic Kinetochore- Microtubule Interface through Assembly of the Dam1 Ring Complex. Mol. Cell 2005, 17, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Alushin, G.M.; Ramey, V.H.; Pasqualato, S.; Ball, D.A.; Grigorieff, N.; Musacchio, A.; Nogales, E. The Ndc80 kinetochore complex forms oligomeric arrays along microtubules. Nature 2010, 467, 805–810. [Google Scholar] [CrossRef]
- Wang, W.; Jiang, Q.; Argentini, M.; Cornu, D.; Gigant, B.; Knossow, M.; Wang, C. Kif2C Minimal Functional Domain Has Unusual Nucleotide Binding Properties That Are Adapted to Microtubule Depolymerization. J. Biol. Chem. 2012, 287, 15143–15153. [Google Scholar] [CrossRef] [PubMed]
- Helenius, J.; Brouhard, G.; Kalaidzidis, Y.; Diez, S.; Howard, J. The depolymerizing kinesin MCAK uses lattice diffusion to rapidly target microtubule ends. Nature 2006, 441, 115–119. [Google Scholar] [CrossRef]
- Desai, A.; Verma, S.; Mitchison, T.J.; Walczak, C.E. Kin I kinesins are microtubule-destabilizing enzymes. Cell 1999, 96, 69–78. [Google Scholar] [CrossRef]
- Walczak, C.E.; Gayek, S.; Ohi, R. Microtubule-Depolymerizing Kinesins. Annu. Rev. Cell Dev. Biol. 2013, 29, 417–441. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.; Langford, K.J.; Askham, J.M.; Brüning-Richardson, A.; Morrison, E.E. MCAK associates with EB1. Oncogene 2008, 27, 2494–2500. [Google Scholar] [CrossRef] [PubMed]
- Montenegro Gouveia, S.; Leslie, K.; Kapitein, L.C.; Buey, R.M.; Grigoriev, I.; Wagenbach, M.; Smal, I.; Meijering, E.; Hoogenraad, C.C.; Wordeman, L.; et al. In Vitro Reconstitution of the Functional Interplay between MCAK and EB3 at Microtubule Plus Ends. Curr. Biol. 2010, 20, 1717–1722. [Google Scholar] [CrossRef] [PubMed]
- Blaineau, C.; Tessier, M.; Dubessay, P.; Tasse, L.; Crobu, L.; Pagès, M.; Bastien, P. A Novel Microtubule-Depolymerizing Kinesin Involved in Length Control of a Eukaryotic Flagellum. Curr. Biol. 2007, 17, 778–782. [Google Scholar] [CrossRef] [PubMed]
- Piao, T.; Luo, M.; Wang, L.; Guo, Y.; Li, D.; Li, P.; Pan, J.; Snell, W.J. A microtubule depolymerizing kinesin functions during both flagellar disassembly and flagellar assembly in Chlamydomonas. Proc. Natl. Acad. Sci. USA 2009, 106, 4713–4718. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.Y.; Ersfeld, K. The role of the Kinesin-13 family protein TbKif13-2 in flagellar length control of Trypanosoma brucei. Mol. Biochem. Parasitol. 2010, 174, 137–140. [Google Scholar] [CrossRef]
- Moore, A.T.; Rankin, K.E.; von Dassow, G.; Peris, L.; Wagenbach, M.; Ovechkina, Y.; Andrieux, A.; Job, D.; Wordeman, L. MCAK associates with the tips of polymerizing microtubules. J. Cell Biol. 2005, 169, 391–397. [Google Scholar] [CrossRef]
- Gupta, M.L.; Carvalho, P.; Roof, D.M.; Pellman, D. Plus end-specific depolymerase activity of Kip3, a kinesin-8 protein, explains its role in positioning the yeast mitotic spindle. Nat. Cell Biol. 2006, 8, 913–923. [Google Scholar] [CrossRef]
- Varga, V.; Helenius, J.; Tanaka, K.; Hyman, A.A.; Tanaka, T.U.; Howard, J. Yeast kinesin-8 depolymerizes microtubules in a length-dependent manner. Nat. Cell Biol. 2006, 8, 957–962. [Google Scholar] [CrossRef]
- Niwa, S.; Nakajima, K.; Miki, H.; Minato, Y.; Wang, D.; Hirokawa, N. KIF19A Is a Microtubule-Depolymerizing Kinesin for Ciliary Length Control. Dev. Cell. 2012, 23, 1167–1175. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, L.B.; Akhmanova, A. Kif7 keeps cilia tips in shape. Nat. Cell Biol. 2014, 16, 623–625. [Google Scholar] [CrossRef] [PubMed]
- Liem, K.F.; He, M.; Ocbina, P.J.R.; Anderson, K.V. Mouse Kif7/Costal2 is a cilia-associated protein that regulates Sonic hedgehog signaling. Proc. Natl. Acad. Sci. USA 2009, 106, 13377–13382. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Subramanian, R.; Bangs, F.; Omelchenko, T.; Liem Jr, K.F.; Kapoor, T.M.; Anderson, K.V. The kinesin-4 protein Kif7 regulates mammalian Hedgehog signalling by organizing the cilium tip compartment. Nat. Cell Biol. 2014, 16, 663–672. [Google Scholar] [CrossRef]
- Briscoe, J.; Thérond, P.P. The mechanisms of Hedgehog signalling and its roles in development and disease. Nat. Rev. Mol. Cell Biol. 2013, 14, 416–429. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.C.; Couzens, A.L.; Deshwar, A.R.; McBroom-Cerajewski, L.D.B.; Zhang, X.; Puviindran, V.; Scott, I.C.; Gingras, A.-C.; Hui, C.-C.; Angers, S. The PPFIA1-PP2A protein complex promotes trafficking of Kif7 to the ciliary tip and Hedgehog signaling. Sci. Signal. 2014, 7, ra117. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, N.; Lane, A.; Jovanovic, K.; Parfitt, D.A.; Aguila, M.; Thompson, C.L.; Da Cruz, L.; Coffey, P.J.; Chapple, J.P.; Hardcastle, A.J.; et al. Arl3 and RP2 regulate the trafficking of ciliary tip kinesins. Hum. Mol. Genet. 2017, 26, 2480–2492. [Google Scholar] [CrossRef] [PubMed]
- Jaulin, F.; Kreitzer, G. KIF17 stabilizes microtubules and contributes to epithelial morphogenesis by acting at MT plus ends with EB1 and APC. J. Cell Biol. 2010, 190, 443–460. [Google Scholar] [CrossRef]
- Evans, J.E.; Snow, J.J.; Gunnarson, A.L.; Ou, G.; Stahlberg, H.; McDonald, K.L.; Scholey, J.M. Functional modulation of IFT kinesins extends the sensory repertoire of ciliated neurons in Caenorhabditis elegans. J. Cell Biol. 2006, 172, 663–669. [Google Scholar] [CrossRef]
- Mesland, D.A.; Hoffman, J.L.; Caligor, E.; Goodenough, U.W. Flagellar tip activation stimulated by membrane adhesions in Chlamydomonas gametes. J. Cell Biol. 1980, 84, 599–617. [Google Scholar] [CrossRef]
- Stepanek, L.; Pigino, G. Microtubule doublets are double-track railways for intraflagellar transport trains. Science 2016, 352, 721–724. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Pang, Y.; Wu, Q.; Hu, Z.; Han, X.; Xu, Y.; Deng, H.; Pan, J. FLA8/KIF3B Phosphorylation Regulates Kinesin-II Interaction with IFT-B to Control IFT Entry and Turnaround. Dev. Cell. 2014, 30, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Liang, Y.; He, W.; Pan, J. Cilia disassembly with two distinct phases of regulation. Cell Rep. 2015, 10, 1803–1810. [Google Scholar] [CrossRef] [PubMed]
- Canning, P.; Park, K.; Gonçalves, J.; Li, C.; Howard, C.J.; Sharpe, T.D.; Holt, L.J.; Pelletier, L.; Bullock, A.N.; Leroux, M.R. CDKL Family Kinases Have Evolved Distinct Structural Features and Ciliary Function. Cell Rep. 2018, 22, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Apagyi, K.; McLeavy, L.; Ersfeld, K. Expression and cellular localisation of calpain-like proteins in Trypanosoma brucei. Mol. Biochem. Parasitol. 2010, 169, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Ono, Y.; Saido, T.C.; Sorimachi, H. Calpain research for drug discovery: Challenges and potential. Nat. Rev. Drug Discov. 2016, 15, 854–876. [Google Scholar] [CrossRef]
- Tonami, K.; Kurihara, Y.; Aburatani, H.; Uchijima, Y.; Asano, T.; Kurihara, H. Calpain 6 Is Involved in Microtubule Stabilization and Cytoskeletal Organization. Mol. Cell Biol. 2007, 27, 2548–2561. [Google Scholar] [CrossRef]
- Olego-Fernandez, S.; Vaughan, S.; Shaw, M.K.; Gull, K.; Ginger, M.L. Cell Morphogenesis of Trypanosoma brucei Requires the Paralogous, Differentially Expressed Calpain-related Proteins CAP5.5 and CAP5.5V. Protist 2009, 160, 576–590. [Google Scholar] [CrossRef]
- Padovano, V.; Podrini, C.; Boletta, A.; Caplan, M.J. Metabolism and mitochondria in polycystic kidney disease research and therapy. Nat. Rev. Nephrol. 2018, 14, 678–687. [Google Scholar] [CrossRef]
- González-Perrett, S.; Kim, K.; Ibarra, C.; Damiano, A.E.; Zotta, E.; Batelli, M.; Harris, P.C.; Reisin, L.L.; Arnaout, M.A.; Cantiello, H.F. Polycystin-2, the protein mutated in autosomal dominant polycystic kidney disease (ADPKD), is a Ca2+-permeable nonselective cation channel. Proc. Natl. Acad. Sci. USA 2001, 98, 1182–1187. [Google Scholar]
- Köttgen, M.; Hofherr, A.; Li, W.; Chu, K.; Cook, S.; Montell, C. Drosophila Sperm Swim Backwards in the Female Reproductive Tract and Are Activated via TRPP2 Ion Channels. PLoS ONE 2011, 6, e20031. [Google Scholar] [CrossRef]
- Lee, J.; Moon, S.; Cha, Y.; Chung, Y.D. Drosophila TRPN( = NOMPC) Channel Localizes to the Distal End of Mechanosensory Cilia. PLoS ONE 2010, 5, e11012. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.-B.; Adams, D.; Paukert, M.; Siba, M.; Sidi, S.; Levin, M.; Gillespie, P.G.; Gründer, S. Xenopus TRPN1 (NOMPC) localizes to microtubule-based cilia in epithelial cells, including inner-ear hair cells. Proc. Natl. Acad. Sci. USA 2005, 102, 12572–12577. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, K.; Sloboda, R.D. Protein arginine methyltransferases interact with intraflagellar transport particles and change location during flagellar growth and resorption. Mol. Biol. Cell 2017, 28, 1208–1222. [Google Scholar] [CrossRef] [PubMed]
- Kubo, A.; Yuba-Kubo, A.; Tsukita, S.; Tsukita, S.; Amagai, M. Sentan: A Novel Specific Component of the Apical Structure of Vertebrate Motile Cilia. Mol. Biol. Cell 2008, 19, 5338–5346. [Google Scholar] [CrossRef] [PubMed]
- Subota, I.; Julkowska, D.; Vincensini, L.; Reeg, N.; Buisson, J.; Blisnick, T.; Huet, D.; Perrot, S.; Santi-Rocca, J.; Duchateau, M.; et al. Proteomic Analysis of Intact Flagella of Procyclic Trypanosoma brucei Cells Identifies Novel Flagellar Proteins with Unique Sub-localization and Dynamics. Mol. Cell Proteom. 2014, 13, 1769–1786. [Google Scholar] [CrossRef] [PubMed]
- Dossou, S.J.Y.; Bré, M.-H.; Hallworth, R. Mammalian cilia function is independent of the polymeric state of tubulin glycylation. Cell Motil. Cytoskeleton 2007, 64, 847–855. [Google Scholar] [CrossRef]
- Oh, S.A.; Allen, T.; Kim, G.J.; Sidorova, A.; Borg, M.; Park, S.K.; Twell, D. Arabidopsis Fused kinase and the Kinesin-12 subfamily constitute a signalling module required for phragmoplast expansion. Plant. J. 2012, 72, 308–319. [Google Scholar] [CrossRef]
- Lee, Y.R.; Liu, B. Identification of a phragmoplast-associated kinesin-related protein in higher plants. Curr. Biol. 2000, 10, 797–800. [Google Scholar] [CrossRef]
- Byron, A.; Askari, J.A.; Humphries, J.D.; Jacquemet, G.; Koper, E.J.; Warwood, S.; Choi, C.K.; Stroud, M.J.; Chen, C.S.; Knight, D.; et al. A proteomic approach reveals integrin activation state-dependent control of microtubule cortical targeting. Nat. Commun. 2015, 6, 6135. [Google Scholar] [CrossRef]
- Goodman, L.; Zallocchi, M. Integrin α8 and Pcdh15 act as a complex to regulate cilia biogenesis in sensory cells. J. Cell Sci. 2017, 130, 3698–3712. [Google Scholar] [CrossRef] [PubMed]
- Nager, A.R.; Goldstein, J.S.; Herranz-Pérez, V.; Portran, D.; Ye, F.; Garcia-Verdugo, J.M.; Nachury, M.V. An Actin Network Dispatches Ciliary GPCRs into Extracellular Vesicles to Modulate Signaling. Cell 2017, 168, 252–263. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Axoneme Pattern | Function | Reference |
|---|---|---|---|
| Lecudina tuzetae (Miozoa; parasitic protozoa) | (6+0) | Motile flagellum | [70,90] |
| Diplauxis hatti (Miozoa; parasitic protozoa) | (3+0) | Motile flagellum | [91] |
| Leishmania mexicana amastigotes (Euglenozoa; parasitic protozoa) | (8+1) or (7+2) | Immotile Flagellum | [92] |
| Caenorhabditis Elegans (Nematoda) | (9+0) changing transiently to 18–22 singlet microtubules at middle region, consequence of doublet splaying | Sensory cilia (Cephalic male (CEM) neurons) | [87] |
| Several species of Arthropoda | (12+0); (14+0); (16+0); (9+9+2) | Motile and non-motile spermatozoa | [89,93,94] |
| Rattus (Chordata) | (8+1) | Immotile cilia (Cerebral cortex) | [95] |
| Organism | Plugs | Central MT 1 Cap | Ciliary Crown | Other | References |
|---|---|---|---|---|---|
| Chlamydomonas (green alga) Tetrahymena (body cilia, ciliate protozoa) Aequipecten (bay scallop gill) | Plug structures are inserted into the tip of the A-tubules of the outer doublets and attached to the membrane by distal filaments | Linked to the membranes | No | [15,104,107,108] | |
| Tetrahymena (oral apparatus cilia, ciliate protozoa) Aequipecten (certain cilia, bay scallop gill) | Plug structures are inserted into the tip of the A-tubules of the outer doublets and attached to the central microtubule cap by distal filaments | Linked to the membranes | No | [104] | |
| Crithidia, Herpetomonas, Trypanosoma and Leishmania (Promastigote; parasitic protozoa) | No | No | No | Blunt end with two dense material regions, one associated with the MT central pair and the other with the doublets | [113] |
| Leishmania (Amastigote, parasitic protozoa) | No | No | No | No dense material or organized structure | [92] |
| Beroe (macrocilia, ctenophore) | No | No | No | Giant capping structure at the tip, formed by extensions of the A and central-pair MTs, bound together by amorphous material | [110] |
| Paratomella (haptocilia, flat worm) | No | No | Structure resemble crown | Asymmetrical cap structure. Some of the doublets end as a single A-microtubule, whereas for others, the A- and B-microtubules terminate in the cap. MT central pair and doublets 1, 2, 3, 8 and 9 terminate in a distal cap, whereas doublets 4 to 7 end at a dense material of a proximal secondary cap | [103,119] |
| Caenorhabditis (sensory amphid channel, worm) | No | No | No | Tip presents only MT singlets. No cap or dense material | [111] |
| Caenorhabditis (sensory amphid wing cilia C, worm) | No | No | No | The middle region of the axoneme both contain doublets and singlets that “splay apart” laterally and end together. The tip presents a membranous fan-like structure. No cap or dense material | [111] |
| Lima (bivalve) | No | No | No | Dense material at the distal ends of the axonemal MTs | [122] |
| Periplaneta (antenna pedicel, cockroach) | No | No | No | Dilated tip, doublets become singlets and terminate in an electron-dense spheroid | [123] |
| Bombina (frog) | Plugs are inserted into the lumen of the A-tubules, attaching them to the caps | No | No | Asymmetrical cap structure. One large cap, linked to the membrane and to doublet MTs number 4 to 7, and a smaller cap linked to the doublets number 1, 2, 3, 8 and 9, as well as to the two central MTs | [121] |
| Primary cilia (vertebrates) | No | No | No | In distal segment doublets are converted into singlets through the loss of the B-tubule. No cap or dense material | [124] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soares, H.; Carmona, B.; Nolasco, S.; Viseu Melo, L.; Gonçalves, J. Cilia Distal Domain: Diversity in Evolutionarily Conserved Structures. Cells 2019, 8, 160. https://doi.org/10.3390/cells8020160
Soares H, Carmona B, Nolasco S, Viseu Melo L, Gonçalves J. Cilia Distal Domain: Diversity in Evolutionarily Conserved Structures. Cells. 2019; 8(2):160. https://doi.org/10.3390/cells8020160
Chicago/Turabian StyleSoares, Helena, Bruno Carmona, Sofia Nolasco, Luís Viseu Melo, and João Gonçalves. 2019. "Cilia Distal Domain: Diversity in Evolutionarily Conserved Structures" Cells 8, no. 2: 160. https://doi.org/10.3390/cells8020160
APA StyleSoares, H., Carmona, B., Nolasco, S., Viseu Melo, L., & Gonçalves, J. (2019). Cilia Distal Domain: Diversity in Evolutionarily Conserved Structures. Cells, 8(2), 160. https://doi.org/10.3390/cells8020160