Decidua Basalis Mesenchymal Stem Cells Favor Inflammatory M1 Macrophage Differentiation In Vitro

 , , ,
, , ,
Abstract
:1. Introduction
2. Experimental Section
2.1. Ethics, Collection of Human Placentae, and Adult Peripheral Blood
2.2. Isolation and Culture of DBMSCs
2.3. Isolation of Human Monocytes
2.4. Culture of Monocyte Derived Macrophages with DBMSCs
2.5. Phagocytic Activity of Monocyte-Derived Macrophages
2.6. T Cell Proliferation Assay
2.7. Quantification of Human Cytokines
2.8. Flow Cytometry
2.9. Statistical Analysis
3. Results and Discussion
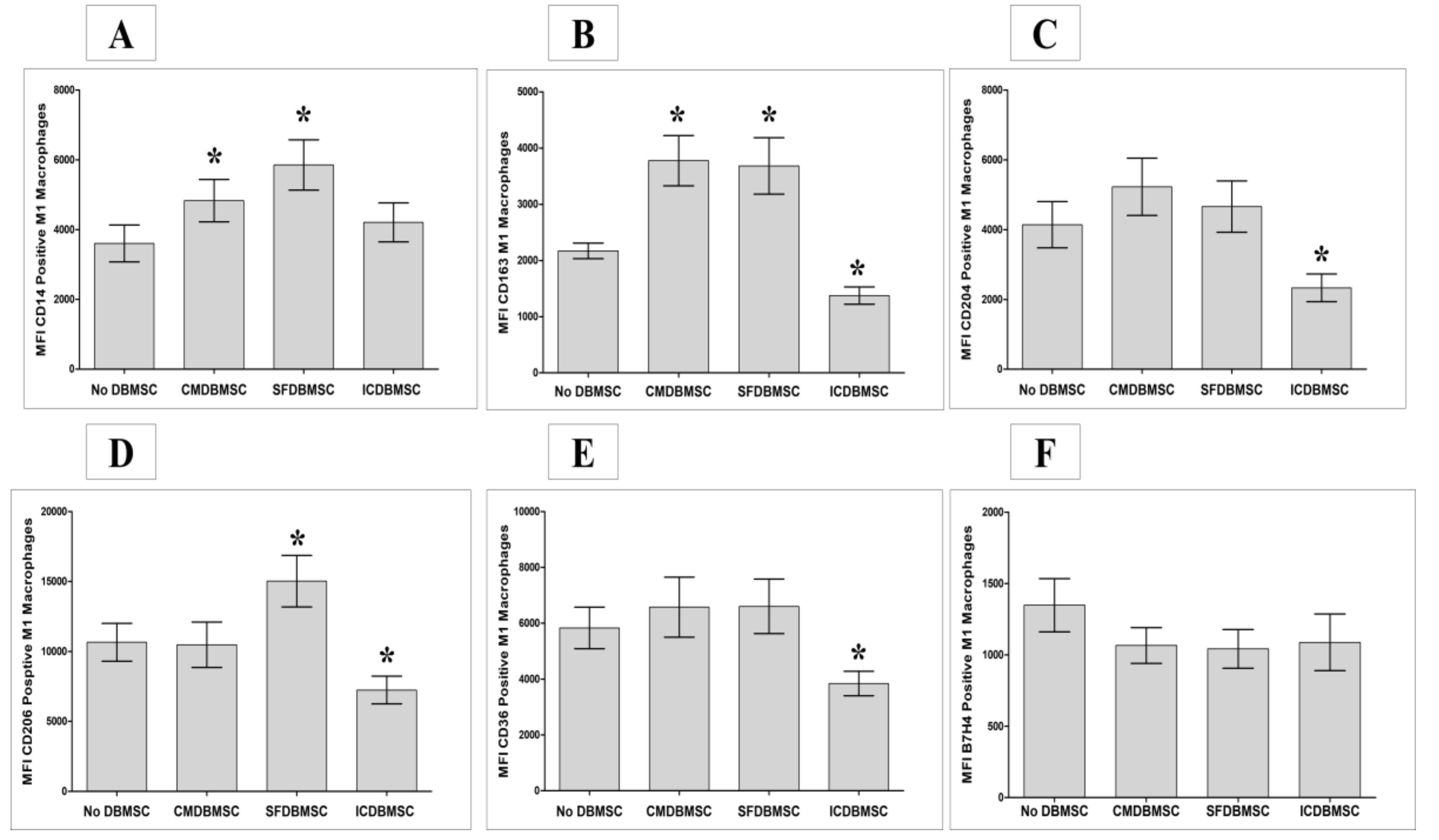
3.1. DBMSCs Effect on M1-like Macrophage Differentiation from Human Monocytes
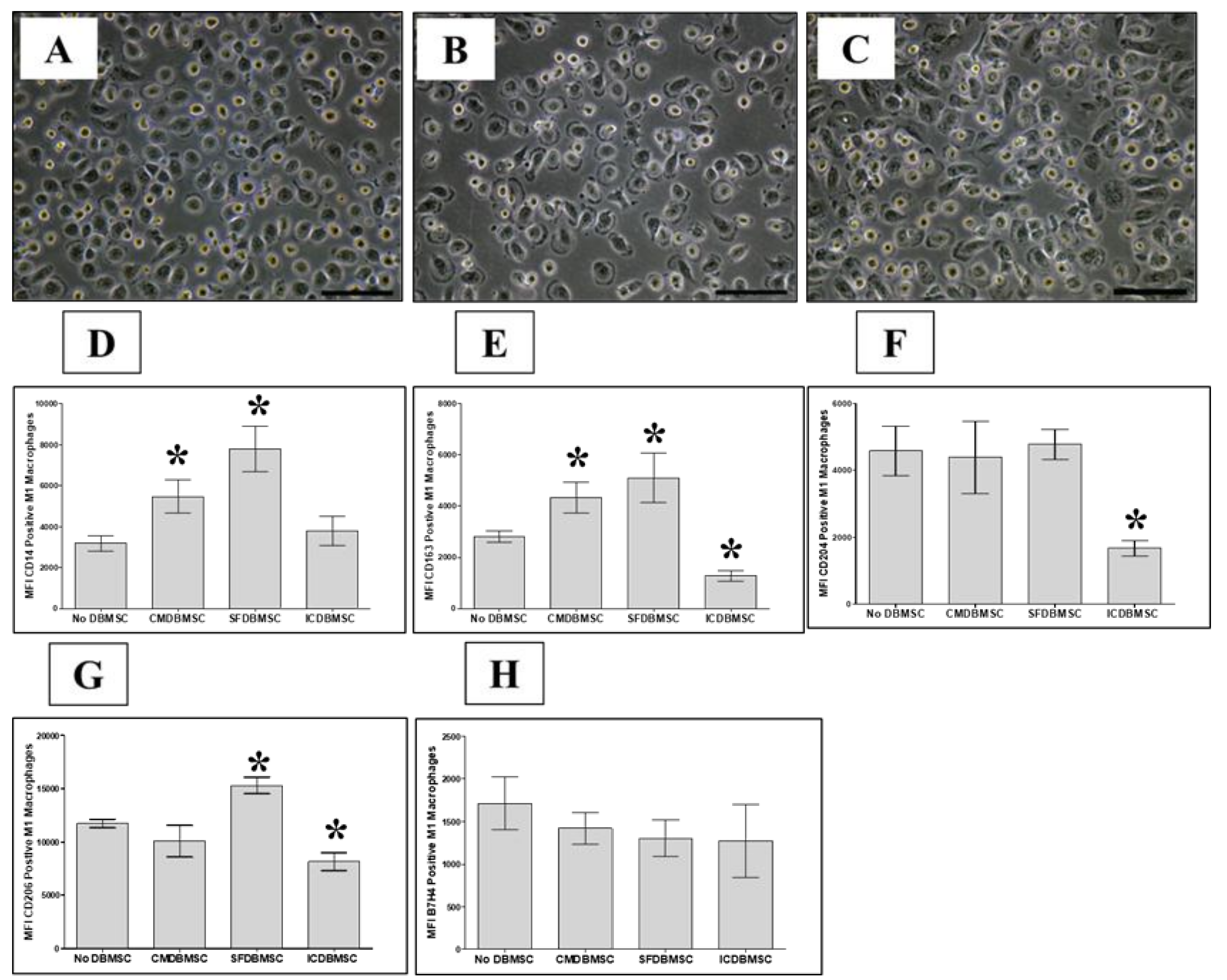
3.2. DBMSC Effects on M1-like Macrophage Differentiation Are Irreversible
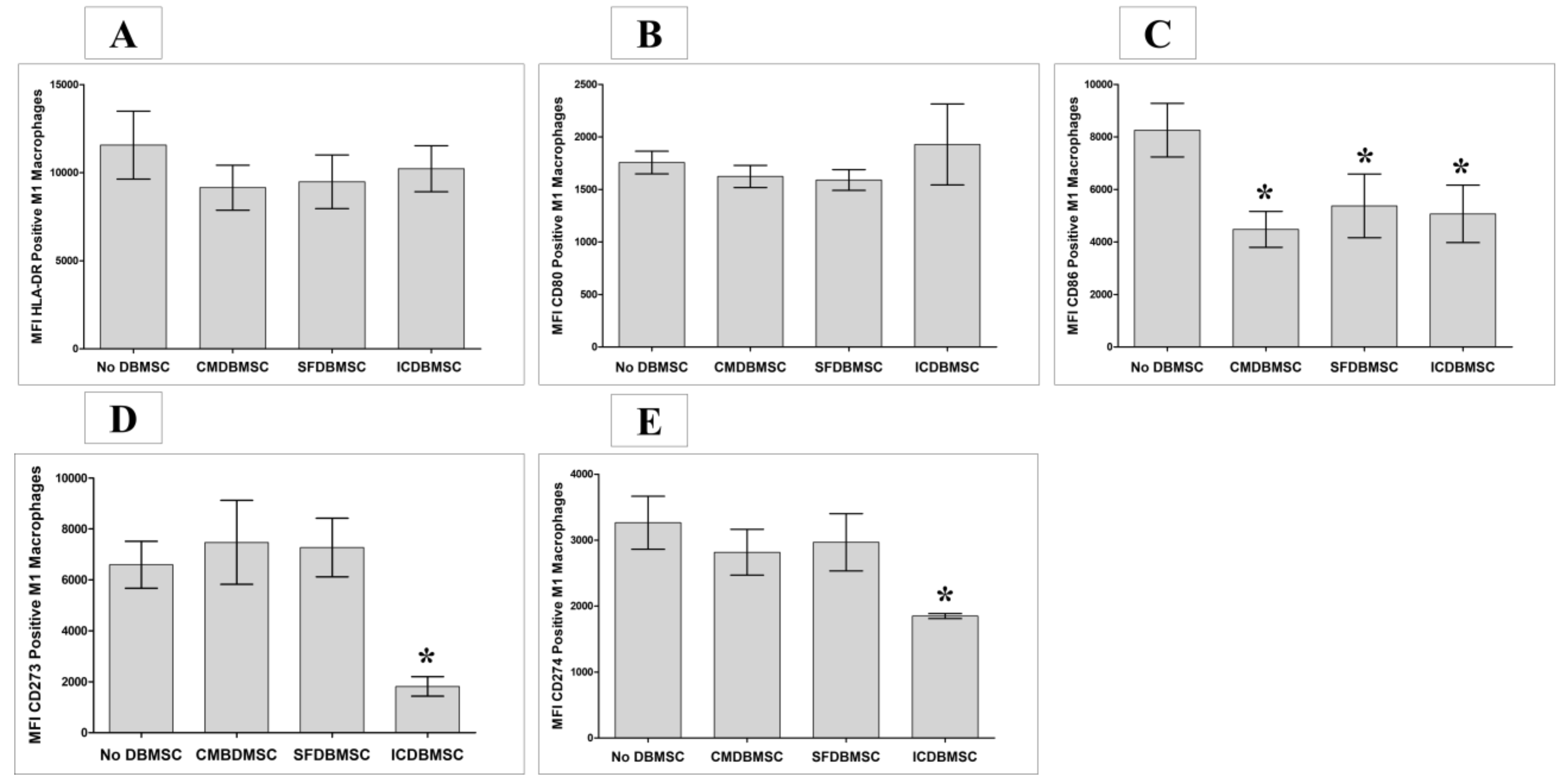
3.3. DBMSC Effects on Expression of CD80, CD86, CD273, CD274, and HLA-DR on Macrophages
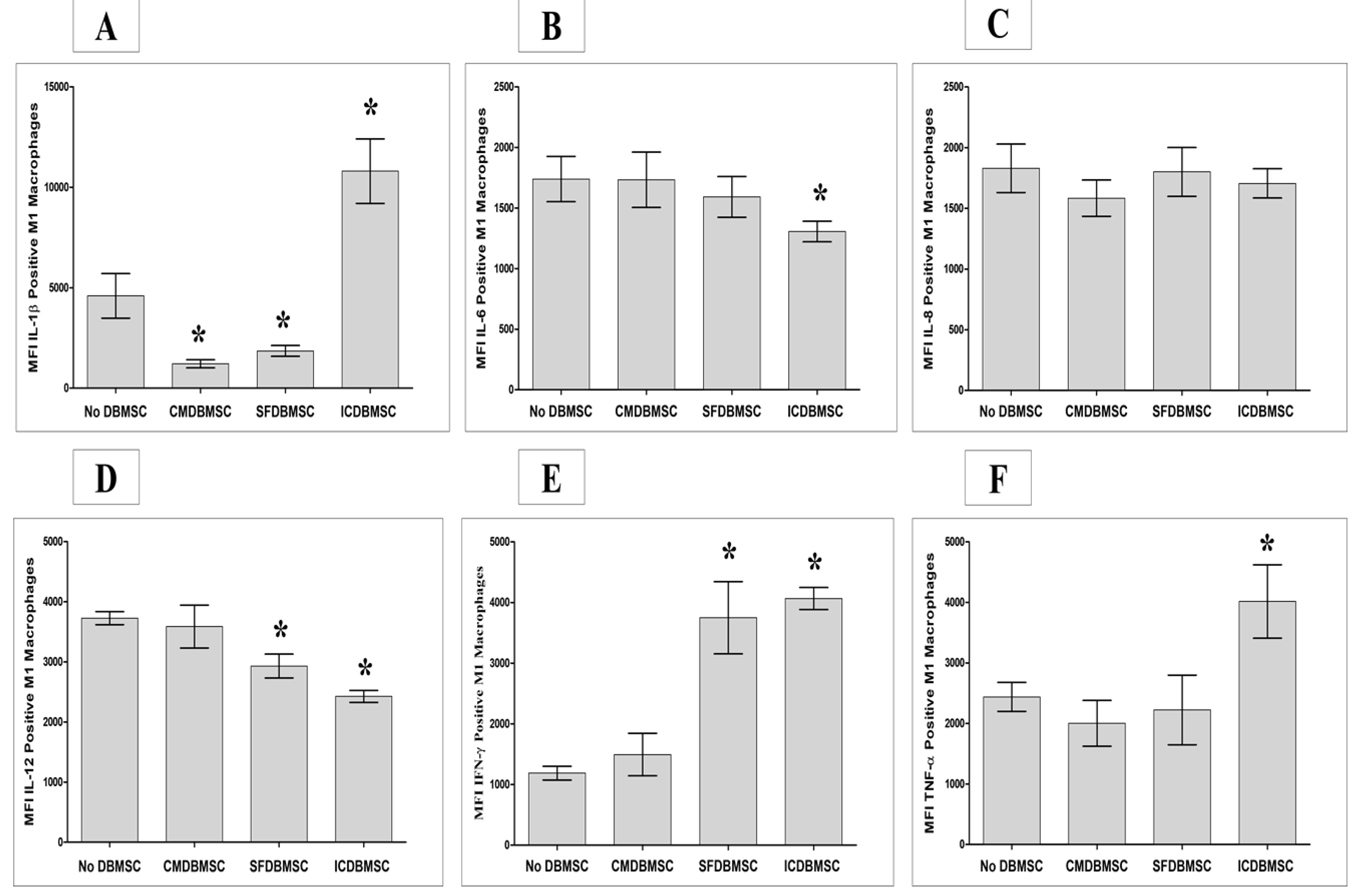
3.4. DBMSCs Modulate Expression by Macrophages of IL-1β, IL-6, IL-8, IL-12, IFN-γ, and TNF-α
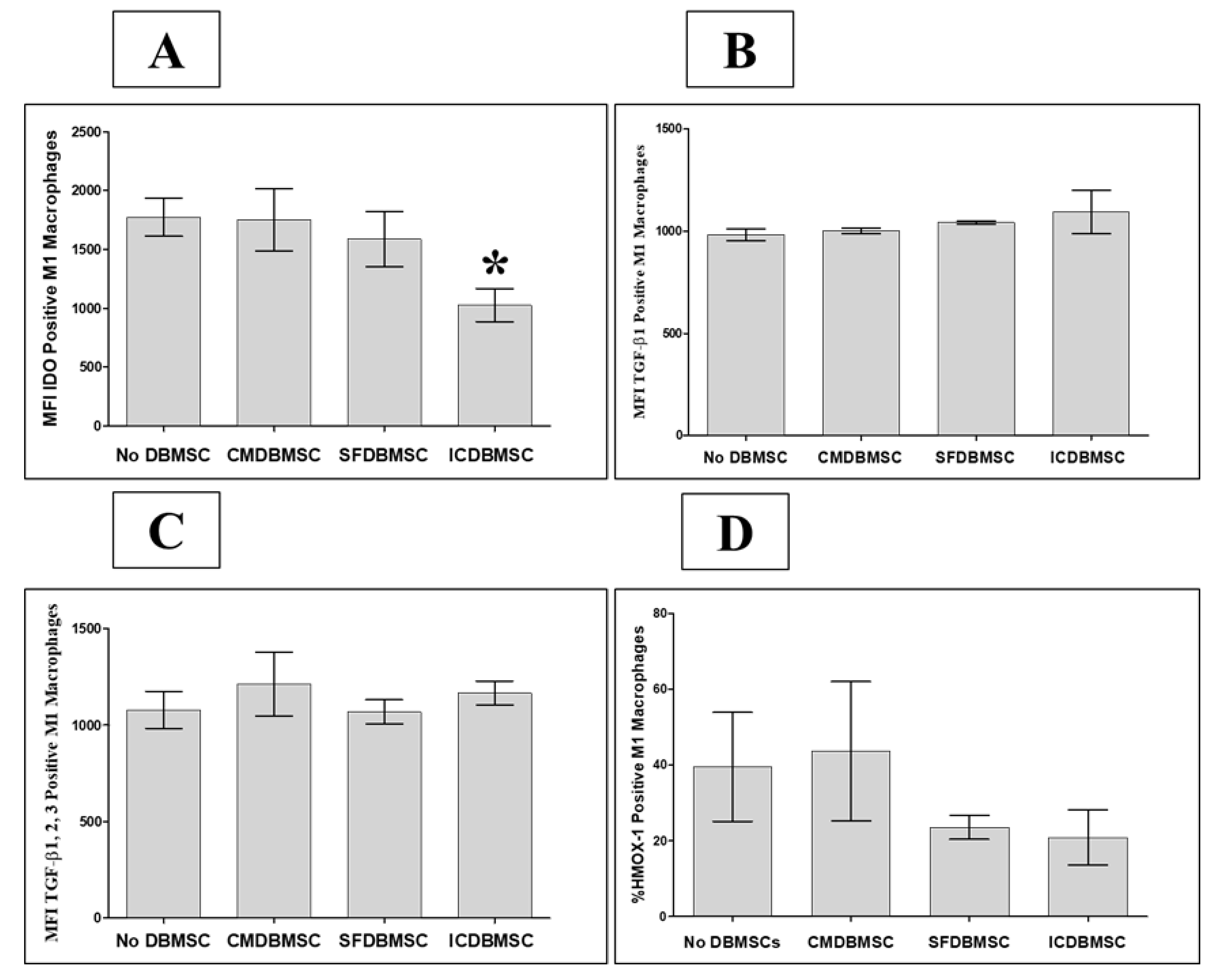
3.5. DBMSC Effects on Expression by Macrophages of IDO, TGFβ1, TGFβ1, 2, 3, and HMOX-1
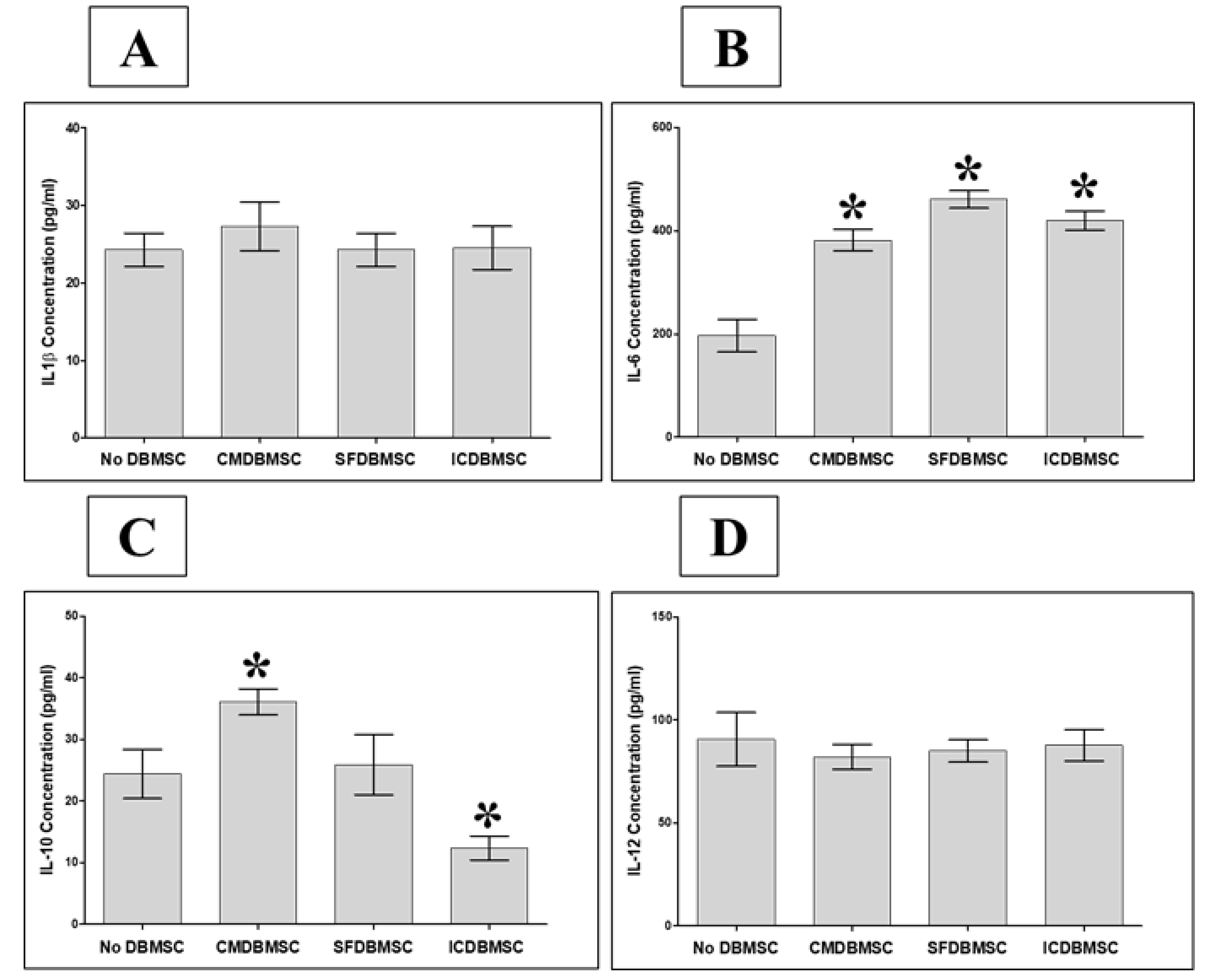
3.6. DBMSCs Modulate Secretion by Macrophages of IL-1β, IL-6, IL-10, and IL-12
3.7. DBMSCs Do Not Alter Macrophage Phagocytic Activity
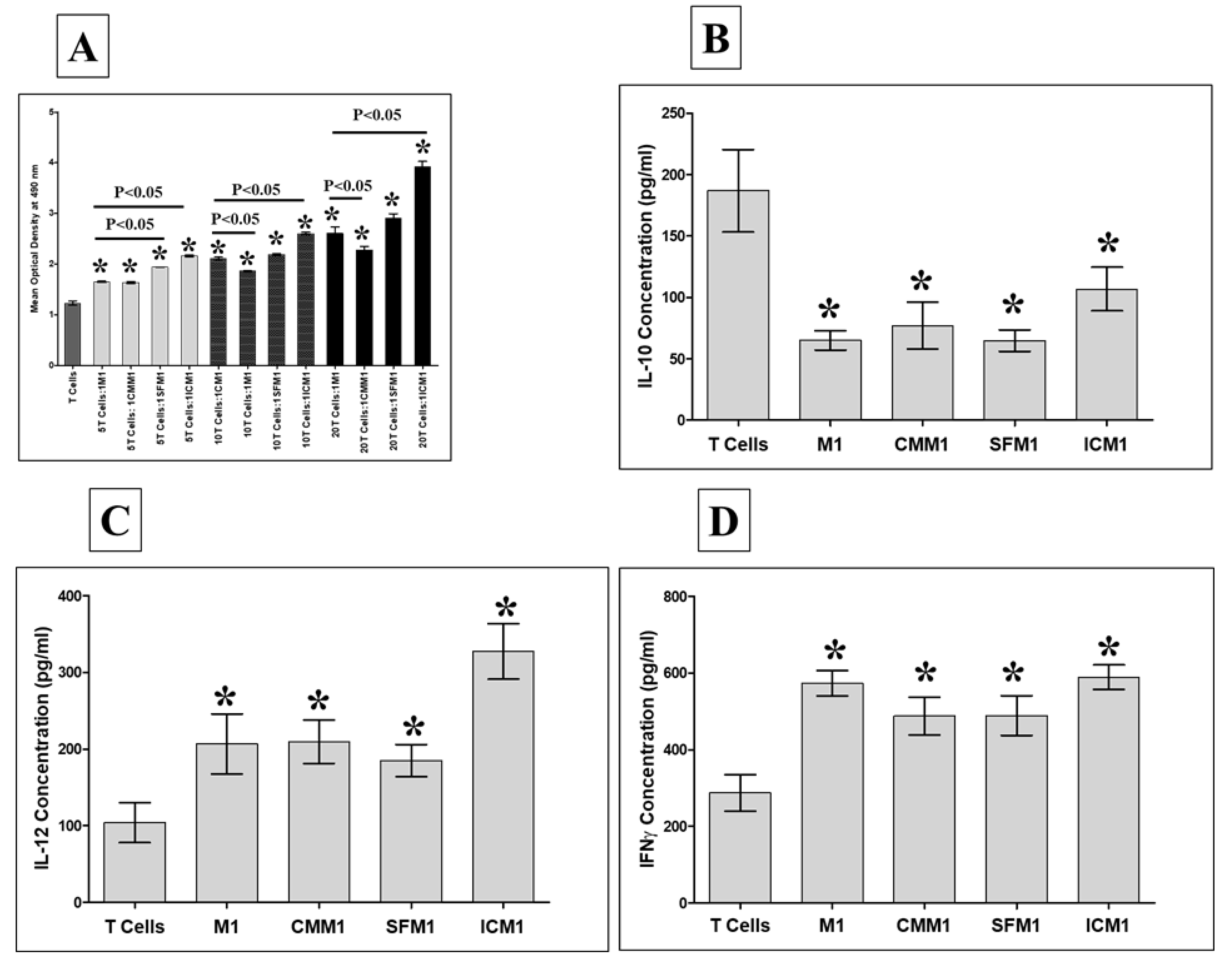
3.8. DBMSCs Induce M1-like Macrophage Effects on T Cell Function
3.9. Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability
Ethics approval and consent to participate
Consent for publication
Author Disclosure Statement
References
- Nielsen, S.R.; Schmid, M.C. Macrophages as key drivers of cancer progression and metastasis. Mediat. Inflamm. 2017, 2017, 9624760. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.J.; Wynn, T.A. Protective and pathogenic functions of macrophage subsets. Nat. Rev. Immunol. 2011, 11, 723–737. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.K.; Mantovani, A. Macrophage plasticity and interaction with lymphocyte subsets: Cancer as a paradigm. Nat. Immunol. 2010, 11, 889–896. [Google Scholar] [CrossRef]
- Mantovani, A.; Sozzani, S.; Locati, M.; Allavena, P.; Sica, A. Macrophage polarization: Tumor-associated macrophages as a paradigm for polarized m2 mononuclear phagocytes. Trends Immunol. 2002, 23, 549–555. [Google Scholar] [CrossRef]
- Ong, S.M.; Tan, Y.C.; Beretta, O.; Jiang, D.; Yeap, W.H.; Tai, J.J.; Wong, W.C.; Yang, H.; Schwarz, H.; Lim, K.H.; et al. Macrophages in human colorectal cancer are pro-inflammatory and prime t cells towards an anti-tumor type-1 inflammatory response. Eur. J. Immunol. 2012, 42, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Abumaree, M.H.; Al Jumah, M.A.; Kalionis, B.; Jawdat, D.; Al Khaldi, A.; Abomaray, F.M.; Fatani, A.S.; Chamley, L.W.; Knawy, B.A. Human placental mesenchymal stem cells (pmscs) play a role as immune suppressive cells by shifting macrophage differentiation from inflammatory m1 to anti-inflammatory m2 macrophages. Stem Cell Rev. 2013, 9, 620–641. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.; Martinez, F.O. Alternative activation of macrophages: Mechanism and functions. Immunity 2010, 32, 593–604. [Google Scholar] [CrossRef]
- Hao, N.B.; Lu, M.H.; Fan, Y.H.; Cao, Y.L.; Zhang, Z.R.; Yang, S.M. Macrophages in tumor microenvironments and the progression of tumors. Clin. Dev. Immunol. 2012, 2012, 948098. [Google Scholar] [CrossRef]
- Kigerl, K.A.; Gensel, J.C.; Ankeny, D.P.; Alexander, J.K.; Donnelly, D.J.; Popovich, P.G. Identification of two distinct macrophage subsets with divergent effects causing either neurotoxicity or regeneration in the injured mouse spinal cord. J. Neurosci. 2009, 29, 13435–13444. [Google Scholar] [CrossRef]
- Abumaree, M.H.; Abomaray, F.M.; Alshabibi, M.A.; AlAskar, A.S.; Kalionis, B. Immunomodulatory properties of human placental mesenchymal stem/stromal cells. Placenta 2017, 59, 87–95. [Google Scholar] [CrossRef]
- Abomaray, F.M.; Al Jumah, M.A.; Kalionis, B.; AlAskar, A.S.; Al Harthy, S.; Jawdat, D.; Al Khaldi, A.; Alkushi, A.; Knawy, B.A.; Abumaree, M.H. Human chorionic villous mesenchymal stem cells modify the functions of human dendritic cells, and induce an anti-inflammatory phenotype in cd1+ dendritic cells. Stem Cell Rev. 2015, 11, 423–441. [Google Scholar] [CrossRef] [PubMed]
- Abomaray, F.M.; Al Jumah, M.A.; Alsaad, K.O.; Jawdat, D.; Al Khaldi, A.; AlAskar, A.S.; Al Harthy, S.; Al Subayyil, A.M.; Khatlani, T.; Alawad, A.O.; et al. Phenotypic and functional characterization of mesenchymal stem/multipotent stromal cells from decidua basalis of human term placenta. Stem Cells Int. 2016, 2016, 5184601. [Google Scholar] [CrossRef] [PubMed]
- Alshabibi, M.A.; Al Huqail, A.J.; Khatlani, T.; Abomaray, F.M.; Alaskar, A.S.; Alawad, A.O.; Kalionis, B.; Abumaree, M.H. Mesenchymal stem/multipotent stromal cells from human decidua basalis reduce endothelial cell activation. Stem Cells Dev. 2017, 26, 1355–1373. [Google Scholar] [CrossRef] [PubMed]
- Abumaree, M.H.; Chamley, L.W.; Badri, M.; El-Muzaini, M.F. Trophoblast debris modulates the expression of immune proteins in macrophages: A key to maternal tolerance of the fetal allograft? J. Reprod. Immunol. 2012, 94, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Hematti, P. Mesenchymal stem cell-educated macrophages: A novel type of alternatively activated macrophages. Exp. Hematol. 2009, 37, 1445–1453. [Google Scholar] [CrossRef] [PubMed]
- Maggini, J.; Mirkin, G.; Bognanni, I.; Holmberg, J.; Piazzon, I.M.; Nepomnaschy, I.; Costa, H.; Canones, C.; Raiden, S.; Vermeulen, M.; et al. Mouse bone marrow-derived mesenchymal stromal cells turn activated macrophages into a regulatory-like profile. PLoS ONE 2010, 5, e9252. [Google Scholar] [CrossRef]
- Zhang, Q.Z.; Su, W.R.; Shi, S.H.; Wilder-Smith, P.; Xiang, A.P.; Wong, A.; Nguyen, A.L.; Kwon, C.W.; Le, A.D. Human gingiva-derived mesenchymal stem cells elicit polarization of m2 macrophages and enhance cutaneous wound healing. Stem Cells 2010, 28, 1856–1868. [Google Scholar] [CrossRef]
- Dayan, V.; Yannarelli, G.; Billia, F.; Filomeno, P.; Wang, X.H.; Davies, J.E.; Keating, A. Mesenchymal stromal cells mediate a switch to alternatively activated monocytes/macrophages after acute myocardial infarction. Basic Res. Cardiol. 2011, 106, 1299–1310. [Google Scholar] [CrossRef]
- Nemeth, K.; Leelahavanichkul, A.; Yuen, P.S.; Mayer, B.; Parmelee, A.; Doi, K.; Robey, P.G.; Leelahavanichkul, K.; Koller, B.H.; Brown, J.M.; et al. Bone marrow stromal cells attenuate sepsis via prostaglandin e(2)-dependent reprogramming of host macrophages to increase their interleukin-10 production. Nat. Med. 2009, 15, 42–49. [Google Scholar] [CrossRef]
- Gupta, N.; Su, X.; Popov, B.; Lee, J.W.; Serikov, V.; Matthay, M.A. Intrapulmonary delivery of bone marrow-derived mesenchymal stem cells improves survival and attenuates endotoxin-induced acute lung injury in mice. J. Immunol. 2007, 179, 1855–1863. [Google Scholar] [CrossRef]
- Adutler-Lieber, S.; Ben-Mordechai, T.; Naftali-Shani, N.; Asher, E.; Loberman, D.; Raanani, E.; Leor, J. Human macrophage regulation via interaction with cardiac adipose tissue-derived mesenchymal stromal cells. J. Cardiovasc. Pharmacol. Ther. 2013, 18, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Raijmakers, M.T.; Roes, E.M.; Poston, L.; Steegers, E.A.; Peters, W.H. The transient increase of oxidative stress during normal pregnancy is higher and persists after delivery in women with pre-eclampsia. Eur. J. Obstet. Gynecol. Reprod. Biol. 2008, 138, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E. Oxidative stress. Best Pract. Res. Clin. Obstet. Gynaecol. 2011, 25, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Braekke, K.; Harsem, N.K.; Staff, A.C. Oxidative stress and antioxidant status in fetal circulation in preeclampsia. Pediatric Res. 2006, 60, 560–564. [Google Scholar] [CrossRef] [PubMed]
- Kusuma, G.D.; Abumaree, M.H.; Pertile, M.D.; Perkins, A.V.; Brennecke, S.P.; Kalionis, B. Mesenchymal stem/stromal cells derived from a reproductive tissue niche under oxidative stress have high aldehyde dehydrogenase activity. Stem Cell Rev. 2016, 12, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Urao, N.; Ushio-Fukai, M. Redox regulation of stem/progenitor cells and bone marrow niche. Free Radic. Biol. Med. 2013, 54, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Waterman, R.S.; Tomchuck, S.L.; Henkle, S.L.; Betancourt, A.M. A new mesenchymal stem cell (msc) paradigm: Polarization into a pro-inflammatory msc1 or an immunosuppressive msc2 phenotype. PLoS ONE 2010, 5, e10088. [Google Scholar] [CrossRef]
- Schaer, D.J.; Alayash, A.I.; Buehler, P.W. Gating the radical hemoglobin to macrophages: The anti-inflammatory role of cd163, a scavenger receptor. Antioxid. Redox Signal. 2007, 9, 991–999. [Google Scholar] [CrossRef]
- Philippidis, P.; Mason, J.C.; Evans, B.J.; Nadra, I.; Taylor, K.M.; Haskard, D.O.; Landis, R.C. Hemoglobin scavenger receptor cd163 mediates interleukin-10 release and heme oxygenase-1 synthesis: Antiinflammatory monocyte-macrophage responses in vitro, in resolving skin blisters in vivo, and after cardiopulmonary bypass surgery. Circ. Res. 2004, 94, 119–126. [Google Scholar] [CrossRef]
- Hamann, W.; Floter, A.; Schmutzler, W.; Zwadlo-Klarwasser, G. Characterization of a novel anti-inflammatory factor produced by rm3/1 macrophages derived from glucocorticoid treated human monocytes. Inflamm. Res. 1995, 44, 535–540. [Google Scholar] [CrossRef]
- Goerdt, S.; Orfanos, C.E. Other functions, other genes: Alternative activation of antigen-presenting cells. Immunity 1999, 10, 137–142. [Google Scholar] [CrossRef]
- Poole, J.A.; Anderson, L.; Gleason, A.M.; West, W.W.; Romberger, D.J.; Wyatt, T.A. Pattern recognition scavenger receptor a/cd204 regulates airway inflammatory homeostasis following organic dust extract exposures. J. Immunotoxicol. 2015, 12, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Woo, M.S.; Yang, J.; Beltran, C.; Cho, S. Cell surface cd36 protein in monocyte/macrophage contributes to phagocytosis during the resolution phase of ischemic stroke in mice. J. Biol. Chem. 2016, 291, 23654–23661. [Google Scholar] [CrossRef] [PubMed]
- Riella, L.V.; Paterson, A.M.; Sharpe, A.H.; Chandraker, A. Role of the pd-1 pathway in the immune response. Am. J. Transplant. 2012, 12, 2575–2587. [Google Scholar] [CrossRef] [PubMed]
- Soliman, H.; Mediavilla-Varela, M.; Antonia, S. Indoleamine 2,3-dioxygenase: Is it an immune suppressor? Cancer J. 2010, 16, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Terai, M.; Tamura, Y.; Alexeev, V.; Mastrangelo, M.J.; Selvan, S.R. Interleukin 10 in the tumor microenvironment: A target for anticancer immunotherapy. Immunol. Res. 2011, 51, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Fisher, D.T.; Appenheimer, M.M.; Evans, S.S. The two faces of il-6 in the tumor microenvironment. Semin. Immunol. 2014, 26, 38–47. [Google Scholar] [CrossRef]
- Di Nicola, M.; Carlo-Stella, C.; Magni, M.; Milanesi, M.; Longoni, P.D.; Matteucci, P.; Grisanti, S.; Gianni, A.M. Human bone marrow stromal cells suppress t-lymphocyte proliferation induced by cellular or nonspecific mitogenic stimuli. Blood 2002, 99, 3838–3843. [Google Scholar] [CrossRef]
- Tse, W.T.; Pendleton, J.D.; Beyer, W.M.; Egalka, M.C.; Guinan, E.C. Suppression of allogeneic t-cell proliferation by human marrow stromal cells: Implications in transplantation. Transplantation 2003, 75, 389–397. [Google Scholar] [CrossRef]
- Rasmusson, I.; Ringden, O.; Sundberg, B.; Le Blanc, K. Mesenchymal stem cells inhibit the formation of cytotoxic t lymphocytes, but not activated cytotoxic t lymphocytes or natural killer cells. Transplantation 2003, 76, 1208–1213. [Google Scholar] [CrossRef]
- Groh, M.E.; Maitra, B.; Szekely, E.; Koc, O.N. Human mesenchymal stem cells require monocyte-mediated activation to suppress alloreactive t cells. Exp. Hematol. 2005, 33, 928–934. [Google Scholar] [CrossRef] [PubMed]
- Corcione, A.; Benvenuto, F.; Ferretti, E.; Giunti, D.; Cappiello, V.; Cazzanti, F.; Risso, M.; Gualandi, F.; Mancardi, G.L.; Pistoia, V.; et al. Human mesenchymal stem cells modulate b-cell functions. Blood 2006, 107, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Quatromoni, J.G.; Eruslanov, E. Tumor-associated macrophages: Function, phenotype, and link to prognosis in human lung cancer. Am. J. Transl. Res. 2012, 4, 376–389. [Google Scholar] [PubMed]
- De Palma, M.; Lewis, C.E. Macrophage regulation of tumor responses to anticancer therapies. Cancer Cell 2013, 23, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Toes, R.E.; Ossendorp, F.; Offringa, R.; Melief, C.J. Cd4 t cells and their role in antitumor immune responses. J. Exp. Med. 1999, 189, 753–756. [Google Scholar] [CrossRef] [PubMed]
- Buechler, C.; Ritter, M.; Orso, E.; Langmann, T.; Klucken, J.; Schmitz, G. Regulation of scavenger receptor cd163 expression in human monocytes and macrophages by pro- and antiinflammatory stimuli. J. Leukoc. Biol. 2000, 67, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Heusinkveld, M.; de Vos van Steenwijk, P.J.; Goedemans, R.; Ramwadhdoebe, T.H.; Gorter, A.; Welters, M.J.; van Hall, T.; van der Burg, S.H. M2 macrophages induced by prostaglandin e2 and il-6 from cervical carcinoma are switched to activated m1 macrophages by cd4+ th1 cells. J. Immunol. 2011, 187, 1157–1165. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Macrophage Markers | Costimulatory and HLA Molecules | Inflammatory Molecules | Anti-inflammatory Molecules |
|---|---|---|---|
| CD14 | CD40 | IL-1β | IDO |
| CD11b | CD80 | IL-6 | TGFβ1 |
| CD36 | CD86 | IL-8 | TGFβ1, 2, 3 |
| CD163 | CD273 | IL-12 | HMOX-1 |
| CD204 | CD274 | IFN-γ | |
| CD206 | HLA-DR | TNF-α | |
| B7-H4 |
| Markers | Types | CMDBMSC | SFDBMSC | ICDBMSC | Markers | Types | CMDBMSC | SFDBMSC | ICDBMSC | |
|---|---|---|---|---|---|---|---|---|---|---|
| CD14 | Functional Markers | ↑ | ↑ | No Change | IDO | Anti-inflammatory Markers | No Change | No Change | ↓ | |
| CD163 | ↑ | ↑ | ↓ | TGFβ1 | No Change | No Change | No Change | |||
| CD204 | No Change | No Change | ↓ | TGFβ1, 2, 3 | No Change | No Change | No Change | |||
| CD206 | No Change | ↑ | ↓ | HMOX-1 | No Change | No Change | No Change | |||
| CD36 | No Change | No Change | ↓ | |||||||
| B7H4 | No Change | No Change | No Change | |||||||
| CD80 | Costimulatory Molecules | No Change | No Change | No Change | ||||||
| CD86 | ↓ | ↓ | ↓ | |||||||
| CD273 | Co-Inhibitory Molecules | No Change | No Change | ↓ | ||||||
| CD274 | No Change | No Change | ↓ | |||||||
| HLA-DR | Antigen Presenting Molecule | No Change | No Change | No Change | ||||||
| IL-1β | Inflammatory Cytokines | ↓ | ↓ | ↑ | ||||||
| IL-6 | No Change | No Change | ↓ | |||||||
| IL-8 | No Change | No Change | No Change | |||||||
| IL-12 | No Change | ↓ | ↓ | |||||||
| IFN-γ | No Change | ↑ | ↑ | |||||||
| TNF-α | No Change | No Change | ↑ | |||||||
| Markers | Type | CMDBMSC | SFDBMSC | ICDBMSC |
|---|---|---|---|---|
| IL-1β | Inflammatory Cytokines | No Change | No Change | No Change |
| IL-6 | ↑ | ↑ | ↑ | |
| IL-10 | ↑ | No Change | ↓ | |
| IL-12 | Anti-inflammatory Cytokine | No Change | No Change | No Change |
| Markers | Type | CM1 | CMM1 | SFM1 | ICM1 |
|---|---|---|---|---|---|
| IL-12 | Inflammatory Cytokine | ↑ | ↑ | ↑ | ↑ |
| IFN-γ | |||||
| IL-10 | Anti-inflammatory Cytokine | ↓ | ↓ | ↓ | ↓ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abumaree, M.H.; Al Harthy, S.; Al Subayyil, A.M.; Alshabibi, M.A.; Abomaray, F.M.; Khatlani, T.; Kalionis, B.; El- Muzaini, M.F.; Al Jumah, M.A.; Jawdat, D.; et al. Decidua Basalis Mesenchymal Stem Cells Favor Inflammatory M1 Macrophage Differentiation In Vitro. Cells 2019, 8, 173. https://doi.org/10.3390/cells8020173
Abumaree MH, Al Harthy S, Al Subayyil AM, Alshabibi MA, Abomaray FM, Khatlani T, Kalionis B, El- Muzaini MF, Al Jumah MA, Jawdat D, et al. Decidua Basalis Mesenchymal Stem Cells Favor Inflammatory M1 Macrophage Differentiation In Vitro. Cells. 2019; 8(2):173. https://doi.org/10.3390/cells8020173
Chicago/Turabian StyleAbumaree, Mohamed H., Seham Al Harthy, Abdullah M. Al Subayyil, Manal A. Alshabibi, Fawaz M. Abomaray, Tanvier Khatlani, Bill Kalionis, Mohammed F. El- Muzaini, Mohammed A. Al Jumah, Dunia Jawdat, and et al. 2019. "Decidua Basalis Mesenchymal Stem Cells Favor Inflammatory M1 Macrophage Differentiation In Vitro" Cells 8, no. 2: 173. https://doi.org/10.3390/cells8020173
APA StyleAbumaree, M. H., Al Harthy, S., Al Subayyil, A. M., Alshabibi, M. A., Abomaray, F. M., Khatlani, T., Kalionis, B., El- Muzaini, M. F., Al Jumah, M. A., Jawdat, D., Alawad, A. O., & AlAskar, A. S. (2019). Decidua Basalis Mesenchymal Stem Cells Favor Inflammatory M1 Macrophage Differentiation In Vitro. Cells, 8(2), 173. https://doi.org/10.3390/cells8020173