The Centriolar Adjunct–Appearance and Disassembly in Spermiogenesis and the Potential Impact on Fertility



Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Sperm Analysis
2.2. Porcine Testicles and Sperm Analysis
2.3. Transmission Electron Microscopy (TEM)
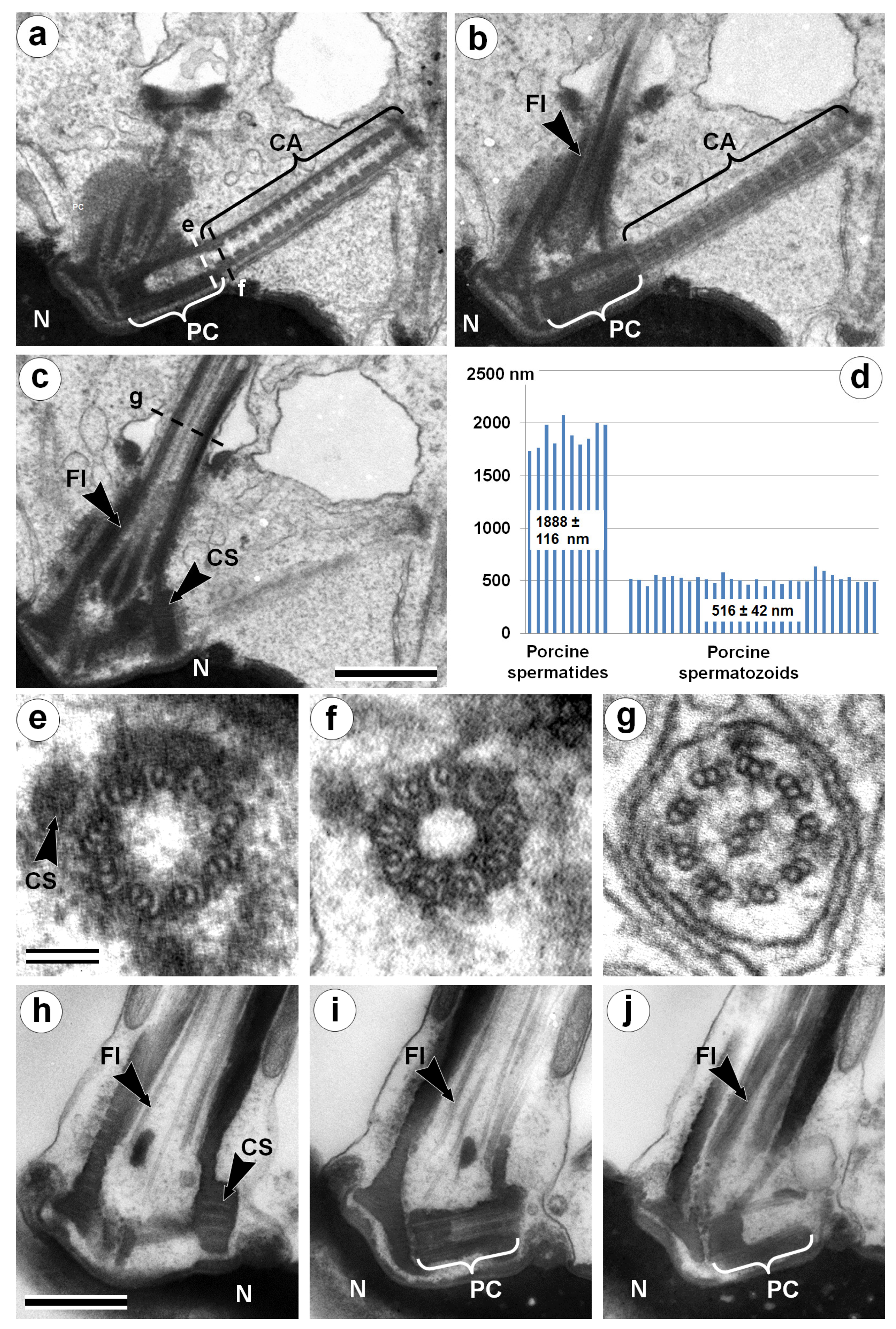
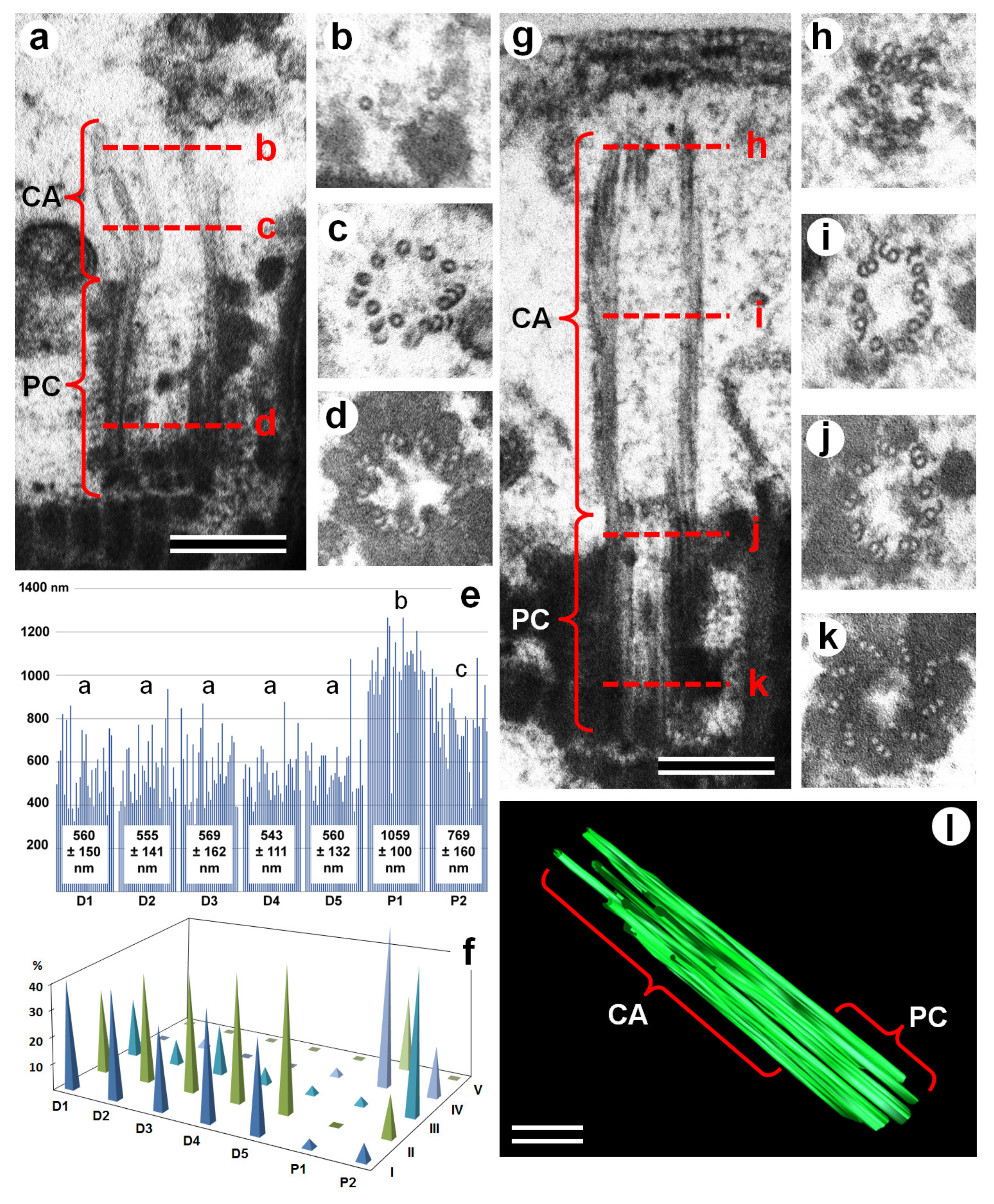
2.4. Measurements and Statistical Analysis
2.5. Three-Dimensional Reconstruction
3. Results and Discussion
3.1. Centriolar Adjunct Structure in Pig Spermatids
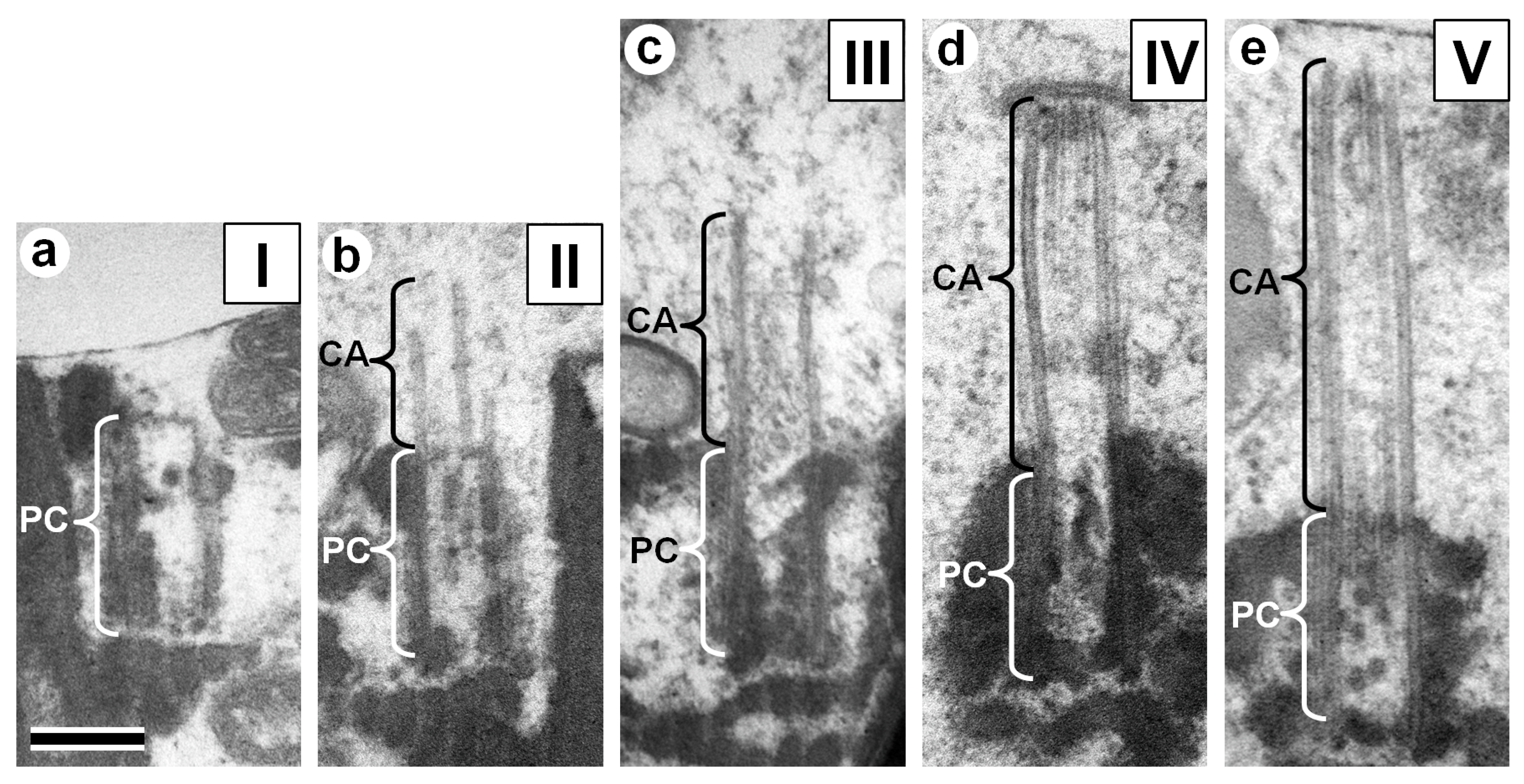
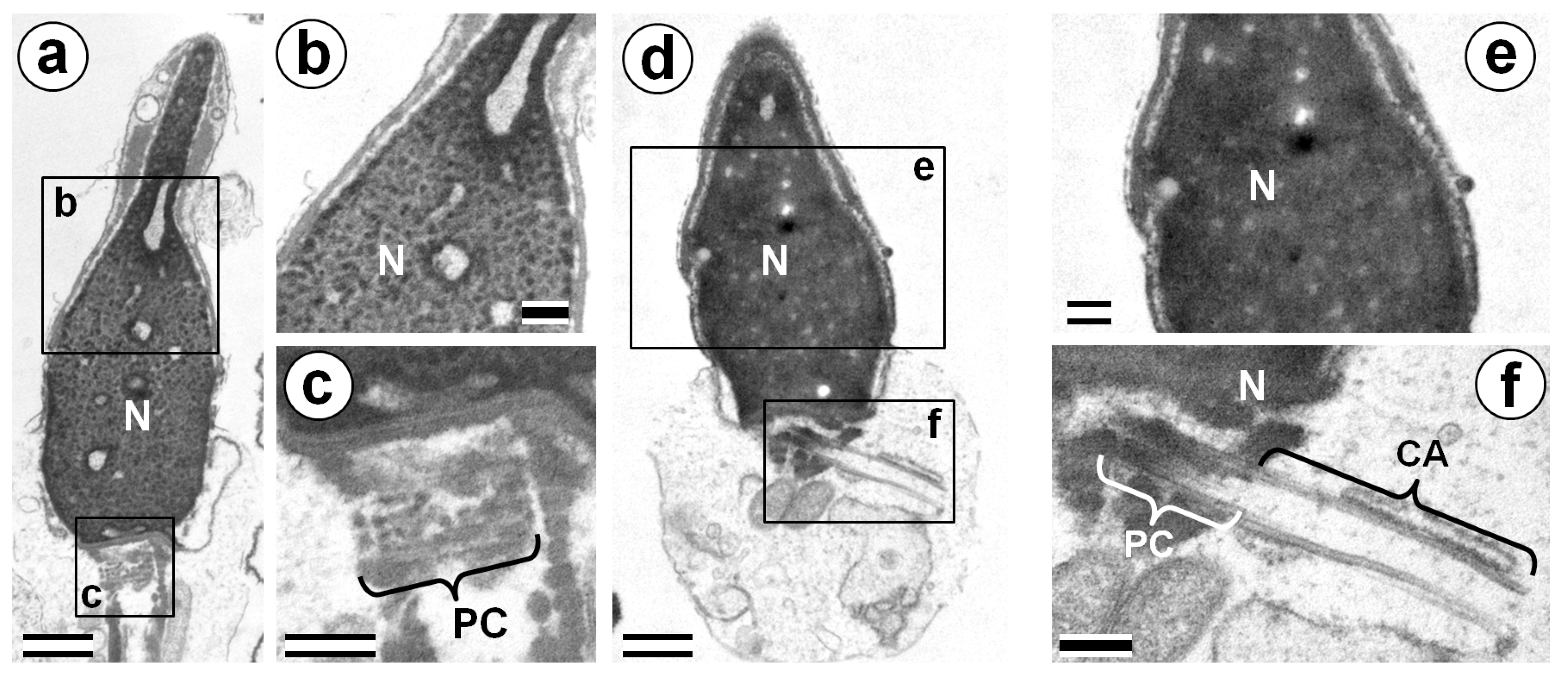
3.2. Centriolar Adjunct Structure in Human Spermatozoa
3.3. Chromatin State in Nuclei of Human IMS Patients and Donors’ Spermatozoa
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Holstein, A.F. Ultrastructural observations on the differentiation of spermatids in man. Andrologia 1976, 8, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Zamboni, L. The ultrastructural pathology of the spermatozoon as a cause of infertility, the role of electron microscopy in the evaluation of semen quality. Fertil. Steril. 1987, 48, 711–734. [Google Scholar] [CrossRef]
- Bragina, E.E.; Abdumalikov, R.A. Manual in Spermatology; Sorek-Poligrafiya Press: Moscow, Russia, 2002; pp. 1–108. ISBN 5-86492-042-8. [Google Scholar]
- Vreeburg, J.; van Roijen, J.; Ooms, M.P.; van der Eijnden, M.; Weber, R.F.; Grootegoed, J. Testis-specific histone 2B in human spermatozoa. Int. J. Androl. 1997, 20 (Suppl. 1), 92–104. [Google Scholar]
- Evenson, D.P.; Darzynkiewicz, Z.; Melamed, M.R. Relation of mammalian sperm chromatin heterogeneity to fertility. Science 1980, 210, 1131–1133. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, P.G.; Manicardi, G.C.; Urner, F.; Campana, A.; Sakkas, D. Chromatin packaging and morphology in ejaculated human spermatozoa: Evidence of hidden anomalies in normal spermatozoa. Mol. Hum. Reprod. 1996, 2, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.Y.; Baker, H.W. Sperm nuclear chromatin normality: Relationship with sperm morphology, sperm-zona pellucida binding, and fertilization rates in vitro. Fertil. Steril. 1992, 58, 1178–1184. [Google Scholar] [CrossRef]
- Sathananthan, A.H.; Ratner, S.S.; Ng, S.C.; Thrin, J.J.; Gianaroli, L.; Trounson, A. The sperm centriole: Its inheritance, replication and perpetuation in early human embryos. Hum. Reprod. 1996, 11, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Schatten, H.; Sun, Q.Y. The role of centrosomes in mammalian fertilization and its significance for ICSI. Mol. Hum. Reprod. 2009, 15, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Avidor-Reiss, T.; Khire, A.; Fishman, E.L.; Jo, K.H. Atypical centrioles during sexual reproduction. Front. Cell Dev. Biol. 2015, 3, 21. [Google Scholar] [CrossRef] [PubMed]
- Fishman, E.L.; Jo, K.; Nguyen, Q.P.H.; Kong, D.; Royfman, R.; Cekic, A.R.; Khanal, S.; Miller, A.L.; Simerly, C.; Schatten, G.; et al. A novel atypical sperm centriole is functional during human fertilization. Nat Commun. 2018, 9, 2210, Erratum in Nat. Commun. 2018, 9, 2800. [Google Scholar] [CrossRef] [PubMed]
- Fawcett, D.W.; Phillips, D.M. The fine structure and development of the neck region of the mammalian spermatozoon. Anat. Rec. 1969, 165, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Fawcett, D.W. The Cell. Second edition. Chapter 12: Centrioles; W.B. Saunders Company Press: Philadelphia, PA, USA; London, UK; Toronto, ON, Canada; Mexico City, Mexico; Rio de Janeiro, Brazil; Sydney, Australia; Tokyo, Japan, 1981; pp. 551–574. ISBN 0-7216-3584-9. [Google Scholar]
- Manandhar, G.; Sutovsky, P.; Joshi, H.C.; Stearns, T.; Schatten, G. Centrosome reduction during mouse spermiogenesis. Dev. Biol. 1998, 203, 424–434. [Google Scholar] [CrossRef] [PubMed]
- Fouquet, J.P.; Kann, M.L.; Combeau, C.; Melki, R. Gamma-tubulin during the differentiation of spermatozoa in various mammals and man. Mol. Hum. Reprod. 1998, 4, 1122–1129. [Google Scholar] [CrossRef] [PubMed]
- Manandhar, G.; Simerly, C.; Schatten, G. Highly degenerated distal centrioles in rhesus and human spermatozoa. Hum. Reprod. 2000, 15, 256–263. [Google Scholar] [CrossRef] [PubMed]
- De Kretser, D.M. Ultrastructural features of human spermiogenesis. Z Zellforsch 1969, 98, 477–505. [Google Scholar] [CrossRef] [PubMed]
- Zamboni, L.; Stefanini, M. The fine structure of the neck of mammalian spermatozoa. Anat. Res. 1971, 169, 155–172. [Google Scholar] [CrossRef] [PubMed]
- Kojima, Y. Ultrastructure of goat testes: Centriolar adjunct in spermiogenesis. J. Vet. Med. Sci. 1994, 56, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Depla, M.; Uzbekov, R.; Hourioux, C.; Blanchard, E.; Le Gouge, A.; Gillet, L.; Roingeard, P. Ultrastructural and quantitative analysis of the lipid droplet clustering induced by hepatitis C virus core protein. Cell Mol. Life Sci. 2010, 67, 3151–3161. [Google Scholar] [CrossRef] [PubMed]
- Uzbekov, R.; Garanina, A.; Bressac, C. Centrioles without microtubules: A new morphological type of centriole. Biol. Open 2018, 7, bio036012. [Google Scholar] [CrossRef] [PubMed]
- Avidor-Reiss, T.; Leroux, M.R. Shared and Distinct Mechanisms of Compartmentalized and Cytosolic Ciliogenesis. Curr. Biol. 2015, 25, R1143–R1150. [Google Scholar] [CrossRef] [PubMed]
- Alieva, I.B.; Staub, C.; Uzbekova, S.; Uzbekov, R. A question of flagella origin for spermatids – mother or daughter centriole? In Flagella and Cilia. Types Structure and Functions; Uzbekov, R.E., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2018; Chapter 5; pp. 109–126. [Google Scholar]
- Izawa, I.; Goto, H.; Kasahara, K.; Inagaki, M. Current topics of functional links between primary cilia and cell cycle. Cilia 2015, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- Phua, S.C.; Chiba, S.; Suzuki, M.; Su, E.; Roberson, E.C.; Pusapati, G.V.; Setou, M.; Rohatgi, R.; Reiter, J.F.; Ikegami, K.; et al. Dynamic Remodeling of Membrane Composition Drives Cell Cycle through Primary Cilia Excision. Cell 2017, 168, 264–279. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Singh, A.K. Trends of male factor infertility, an important cause of infertility: A review of literature. J. Hum. Reprod. Sci. 2015, 8, 191–196. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Donors (D1–D5) and Patients (P1, P2) | D1 | D2 | D3 | D4 | D5 | P1 | P2 |
|---|---|---|---|---|---|---|---|
| Length of complex PC+CA (nm) | 560 ± 150 | 555 ± 141 | 569 ± 162 | 543 ± 111 | 560 ± 132 | 1059 ± 100 | 769 ± 160 |
| % of spermatozoa without centriolar adjunct (I class) | 43.3 | 43.3 | 33.3 | 43.3 | 36.7 | 3.3 | 6.7 |
| % of spermatozoa population with complete condensation of chromatin | 83.6 (n = 535) | 81.2 (n = 521) | 72.6 (n = 594) | 85.1 (n = 510) | 88.8 (n = 500) | 39.9 (n = 501) | 88.8 (n = 525) |
| % of “normal spermatozoa” by two parameters: centriolar adjunct disassembly and chromatin condensation | 36.2 | 35.2 | 24.1 | 36.8 | 32.6 | 1.3 | 5.9 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garanina, A.S.; Alieva, I.B.; Bragina, E.E.; Blanchard, E.; Arbeille, B.; Guerif, F.; Uzbekova, S.; Uzbekov, R.E. The Centriolar Adjunct–Appearance and Disassembly in Spermiogenesis and the Potential Impact on Fertility. Cells 2019, 8, 180. https://doi.org/10.3390/cells8020180
Garanina AS, Alieva IB, Bragina EE, Blanchard E, Arbeille B, Guerif F, Uzbekova S, Uzbekov RE. The Centriolar Adjunct–Appearance and Disassembly in Spermiogenesis and the Potential Impact on Fertility. Cells. 2019; 8(2):180. https://doi.org/10.3390/cells8020180
Chicago/Turabian StyleGaranina, Anastasiia S., Irina B. Alieva, Elizaveta E. Bragina, Emmanuelle Blanchard, Brigitte Arbeille, Fabrice Guerif, Svetlana Uzbekova, and Rustem E. Uzbekov. 2019. "The Centriolar Adjunct–Appearance and Disassembly in Spermiogenesis and the Potential Impact on Fertility" Cells 8, no. 2: 180. https://doi.org/10.3390/cells8020180
APA StyleGaranina, A. S., Alieva, I. B., Bragina, E. E., Blanchard, E., Arbeille, B., Guerif, F., Uzbekova, S., & Uzbekov, R. E. (2019). The Centriolar Adjunct–Appearance and Disassembly in Spermiogenesis and the Potential Impact on Fertility. Cells, 8(2), 180. https://doi.org/10.3390/cells8020180