Tubulin βII and βIII Isoforms as the Regulators of VDAC Channel Permeability in Health and Disease

 , ,
, ,
Abstract
:1. Introduction
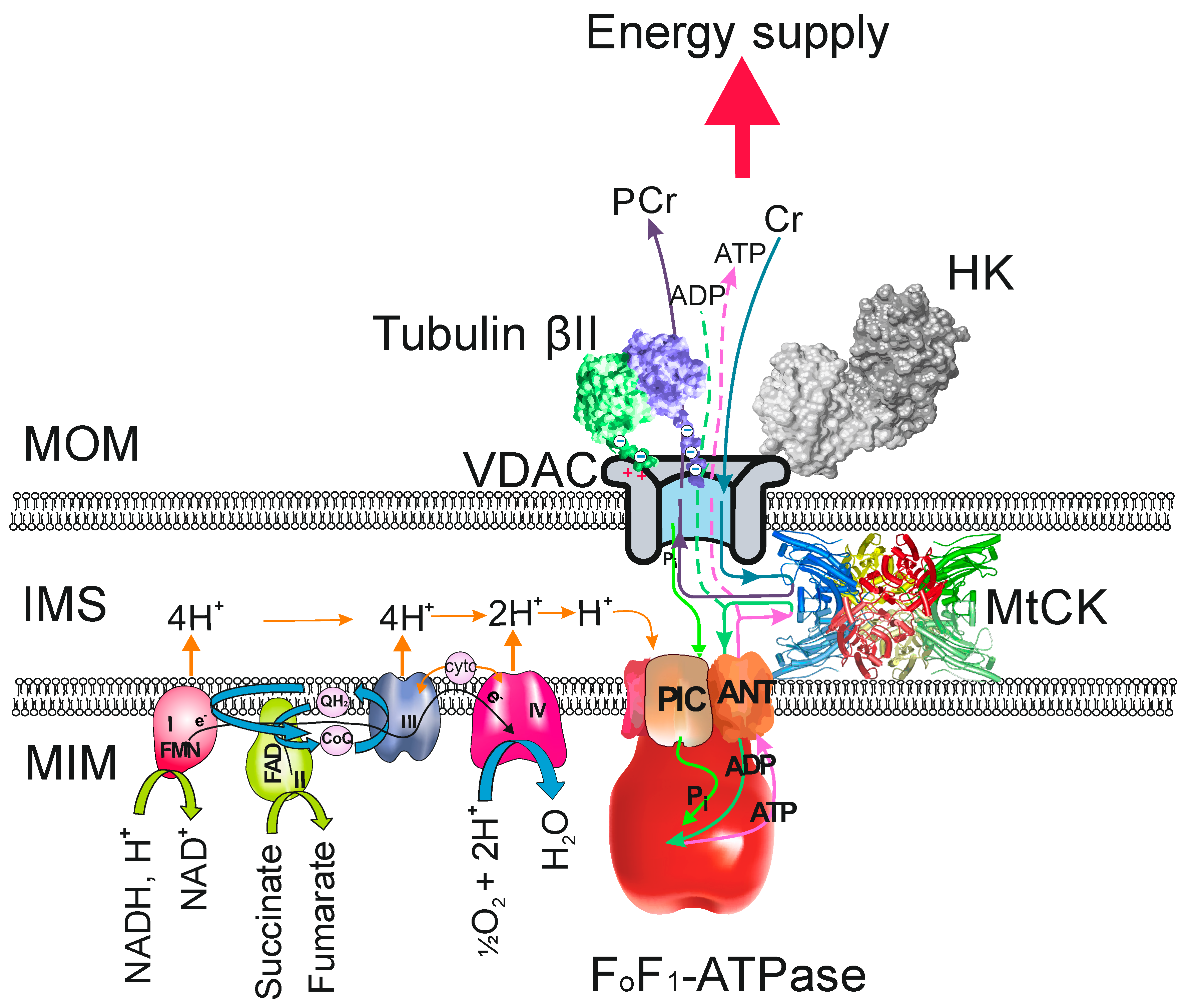
2. Tubulin as the Regulator of VDAC Permeability in Oxidative Skeletal and Heart Muscles
3. Tubulin as the Regulator of VDAC Permeability in Brain Synaptosomes
4. The Regulatory Aspects of Tubulin on VDAC Permeability in Cancer Cells
5. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ANT | Adenine nucleotide translocase |
| CTT | C-terminal tale |
| HK | Hexokinase |
| IMS | Mitochondrial intermembrane space |
| MI | Mitochondrial Interactosome |
| MT | Microtubule |
| MIM | Mitochondrial inner membrane |
| MOM | Mitochondrial outer membrane |
| MtCK | Mitochondrial creatine kinase |
| OXPHOS | Oxidative phosphorylation |
| VDAC | Voltage-dependent anion channel |
References
- Findeisen, P.; Muhlhausen, S.; Dempewolf, S.; Hertzog, J.; Zietlow, A.; Carlomagno, T.; Kollmar, M. Six subgroups and extensive recent duplications characterize the evolution of the eukaryotic tubulin protein family. Genome Biol. Evol. 2014, 6, 2274–2288. [Google Scholar] [CrossRef] [PubMed]
- Leandro-Garcia, L.J.; Leskela, S.; Landa, I.; Montero-Conde, C.; Lopez-Jimenez, E.; Leton, R.; Cascon, A.; Robledo, M.; Rodriguez-Antona, C. Tumoral and tissue-specific expression of the major human beta-tubulin isotypes. Cytoskeleton 2010, 67, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Rostovtseva, T.K.; Gurnev, P.A.; Hoogerheide, D.P.; Rovini, A.; Sirajuddin, M.; Bezrukov, S.M. Sequence diversity of tubulin isotypes in regulation of the mitochondrial voltage-dependent anion channel. J. Biol. Chem. 2018, 293, 10949–10962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sackett, D.L.; Werbovetz, K.A.; Morrissette, N.S. Isolating tubulin from nonneural sources. Methods Cell Biol. 2010, 95, 17–32. [Google Scholar] [CrossRef]
- Yu, I.; Garnham, C.P.; Roll-Mecak, A. Writing and reading the tubulin code. J. Biol. Chem. 2015, 290, 17163–17172. [Google Scholar] [CrossRef] [PubMed]
- Tepp, K.; Mado, K.; Varikmaa, M.; Klepinin, A.; Timohhina, N.; Shevchuk, I.; Chekulayev, V.; Kuznetsov, A.V.; Guzun, R.; Kaambre, T. The role of tubulin in the mitochondrial metabolism and arrangement in muscle cells. J. Bioenerg. Biomembr. 2014, 46, 421–434. [Google Scholar] [CrossRef] [PubMed]
- Shoshan-Barmatz, V.; Israelson, A.; Brdiczka, D.; Sheu, S.S. The voltage-dependent anion channel (vdac): Function in intracellular signalling, cell life and cell death. Curr. Pharm. Des. 2006, 12, 2249–2270. [Google Scholar] [CrossRef]
- Arif, T.; Krelin, Y.; Shoshan-Barmatz, V. Reducing vdac1 expression induces a non-apoptotic role for pro-apoptotic proteins in cancer cell differentiation. Biochim. Biophys. Acta 2016, 1857, 1228–1242. [Google Scholar] [CrossRef] [PubMed]
- Shoshan-Barmatz, V.; Krelin, Y.; Shteinfer-Kuzmine, A.; Arif, T. Voltage-dependent anion channel 1 as an emerging drug target for novel anti-cancer therapeutics. Front. Oncol. 2017, 7, 154. [Google Scholar] [CrossRef]
- Sheldon, K.L.; Gurnev, P.A.; Bezrukov, S.M.; Sackett, D.L. Tubulin tail sequences and post-translational modifications regulate closure of mitochondrial voltage-dependent anion channel (vdac). J. Biol. Chem. 2015, 290, 26784–26789. [Google Scholar] [CrossRef]
- Guzun, R.; Gonzalez-Granillo, M.; Karu-Varikmaa, M.; Grichine, A.; Usson, Y.; Kaambre, T.; Guerrero-Roesch, K.; Kuznetsov, A.; Schlattner, U.; Saks, V. Regulation of respiration in muscle cells in vivo by vdac through interaction with the cytoskeleton and mtck within mitochondrial interactosome. Biochim. Biophys. Acta 2012, 1818, 1545–1554. [Google Scholar] [CrossRef]
- Timohhina, N.; Guzun, R.; Tepp, K.; Monge, C.; Varikmaa, M.; Vija, H.; Sikk, P.; Kaambre, T.; Sackett, D.; Saks, V. Direct measurement of energy fluxes from mitochondria into cytoplasm in permeabilized cardiac cells in situ: Some evidence for mitochondrial interactosome. J. Bioenerg. Biomembr. 2009, 41, 259–275. [Google Scholar] [CrossRef] [PubMed]
- Anmann, T.; Varikmaa, M.; Timohhina, N.; Tepp, K.; Shevchuk, I.; Chekulayev, V.; Saks, V.; Kaambre, T. Formation of highly organized intracellular structure and energy metabolism in cardiac muscle cells during postnatal development of rat heart. Biochim. Biophys. Acta 2014, 1837, 1350–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tepp, K.; Puurand, M.; Timohhina, N.; Adamson, J.; Klepinin, A.; Truu, L.; Shevchuk, I.; Chekulayev, V.; Kaambre, T. Changes in the mitochondrial function and in the efficiency of energy transfer pathways during cardiomyocyte aging. Mol. Cell. Biochem. 2017, 432, 141–158. [Google Scholar] [CrossRef] [PubMed]
- Dzeja, P.P.; Terzic, A. Phosphotransfer networks and cellular energetics. J. Exp. Biol. 2003, 206, 2039–2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saks, V.; Schlattner, U.; Tokarska-Schlattner, M.; Wallimann, T.; Bagur, R.; Zorman, S.; Pelosse, M.; Santos, P.; Boucher, F.; Kaambre, T.; et al. Systems level regulation of cardiac energy fluxes via metabolic cycles: Role of creatine, phosphotransfer pathways, and ampk signaling. In Systems Biology of Metabolic and Signaling Networks; Aon, M.A., Saks, V., Schlattner, U., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; Volume 16, pp. 261–320. [Google Scholar]
- Saks, V.A.; Kuznetsov, A.V.; Vendelin, M.; Guerrero, K.; Kay, L.; Seppet, E.K. Functional coupling as a basic mechanism of feedback regulation of cardiac energy metabolism. Mol. Cell. Biochem. 2004, 256, 185–199. [Google Scholar] [CrossRef]
- Wallimann, T.; Wyss, M.; Brdiczka, D.; Nicolay, K.; Eppenberger, H.M. Intracellular compartmentation, structure and function of creatine kinase isoenzymes in tissues with high and fluctuating energy demands: The ‘phosphocreatine circuit’ for cellular energy homeostasis. Biochem. J. 1992, 281, 21–40. [Google Scholar] [CrossRef]
- Maldonado, E.N.; Lemasters, J.J. Warburg revisited: Regulation of mitochondrial metabolism by voltage-dependent anion channels in cancer cells. J. Pharmacol. Exp. Ther. 2012, 342, 637–641. [Google Scholar] [CrossRef]
- Rostovtseva, T.K.; Sheldon, K.L.; Hassanzadeh, E.; Monge, C.; Saks, V.; Bezrukov, S.M.; Sackett, D.L. Tubulin binding blocks mitochondrial voltage-dependent anion channel and regulates respiration. Proc. Natl. Acad. Sci. USA 2008, 105, 18746–18751. [Google Scholar] [CrossRef] [Green Version]
- Kuznetsov, A.V.; Tiivel, T.; Sikk, P.; Kaambre, T.; Kay, L.; Daneshrad, Z.; Rossi, A.; Kadaja, L.; Peet, N.; Seppet, E.; et al. Striking differences between the kinetics of regulation of respiration by adp in slow-twitch and fast-twitch muscles in vivo. Eur. J. Biochem. 1996, 241, 909–915. [Google Scholar] [CrossRef]
- Veksler, V.I.; Kuznetsov, A.V.; Anflous, K.; Mateo, P.; van Deursen, J.; Wieringa, B.; Ventura-Clapier, R. Muscle creatine kinase-deficient mice. Ii. Cardiac and skeletal muscles exhibit tissue-specific adaptation of the mitochondrial function. J. Biol. Chem. 1995, 270, 19921–19929. [Google Scholar] [CrossRef] [PubMed]
- Saks, V.A.; Kuznetsov, A.V.; Khuchua, Z.A.; Vasilyeva, E.V.; Belikova, J.O.; Kesvatera, T.; Tiivel, T. Control of cellular respiration in vivo by mitochondrial outer membrane and by creatine kinase. A new speculative hypothesis: Possible involvement of mitochondrial-cytoskeleton interactions. J. Mol. Cell. Cardiol. 1995, 27, 625–645. [Google Scholar] [CrossRef]
- Saks, V.A.; Tiivel, T.; Kay, L.; Novel-Chate, V.; Daneshrad, Z.; Rossi, A.; Fontaine, E.; Keriel, C.; Leverve, X.; Ventura-Clapier, R.; et al. On the regulation of cellular energetics in health and disease. Mol. Cell. Biochem. 1996, 160–161, 195–208. [Google Scholar] [CrossRef]
- Saks, V.A.; Veksler, V.I.; Kuznetsov, A.V.; Kay, L.; Sikk, P.; Tiivel, T.; Tranqui, L.; Olivares, J.; Winkler, K.; Wiedemann, F.; et al. Permeabilized cell and skinned fiber techniques in studies of mitochondrial function in vivo. Mol. Cell. Biochem. 1998, 184, 81–100. [Google Scholar] [CrossRef] [PubMed]
- Tiivel, T.; Kadaya, L.; Kuznetsov, A.; Kaambre, T.; Peet, N.; Sikk, P.; Braun, U.; Ventura-Clapier, R.; Saks, V.; Seppet, E.K. Developmental changes in regulation of mitochondrial respiration by adp and creatine in rat heart in vivo. Mol. Cell. Biochem. 2000, 208, 119–128. [Google Scholar] [CrossRef]
- Saks, V.A.; Kaambre, T.; Sikk, P.; Eimre, M.; Orlova, E.; Paju, K.; Piirsoo, A.; Appaix, F.; Kay, L.; Regitz-Zagrosek, V.; et al. Intracellular energetic units in red muscle cells. Biochem. J. 2001, 356, 643–657. [Google Scholar] [CrossRef]
- Seppet, E.K.; Kaambre, T.; Sikk, P.; Tiivel, T.; Vija, H.; Tonkonogi, M.; Sahlin, K.; Kay, L.; Appaix, F.; Braun, U.; et al. Functional complexes of mitochondria with ca, mgatpases of myofibrils and sarcoplasmic reticulum in muscle cells. Biochim. Biophys. Acta 2001, 1504, 379–395. [Google Scholar] [CrossRef]
- Eimre, M.; Puhke, R.; Alev, K.; Seppet, E.; Sikkut, A.; Peet, N.; Kadaja, L.; Lenzner, A.; Haviko, T.; Seene, T.; et al. Altered mitochondrial apparent affinity for adp and impaired function of mitochondrial creatine kinase in gluteus medius of patients with hip osteoarthritis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R1271–R1275. [Google Scholar] [CrossRef]
- Varikmaa, M.; Bagur, R.; Kaambre, T.; Grichine, A.; Timohhina, N.; Tepp, K.; Shevchuk, I.; Chekulayev, V.; Metsis, M.; Boucher, F.; et al. Role of mitochondria-cytoskeleton interactions in respiration regulation and mitochondrial organization in striated muscles. Biochim. Biophys. Acta 2014, 1837, 232–245. [Google Scholar] [CrossRef] [PubMed]
- Appaix, F.; Kuznetsov, A.V.; Usson, Y.; Kay, L.; Andrienko, T.; Olivares, J.; Kaambre, T.; Sikk, P.; Margreiter, R.; Saks, V. Possible role of cytoskeleton in intracellular arrangement and regulation of mitochondria. Exp. Physiol. 2003, 88, 175–190. [Google Scholar] [CrossRef]
- Kay, L.; Li, Z.; Mericskay, M.; Olivares, J.; Tranqui, L.; Fontaine, E.; Tiivel, T.; Sikk, P.; Kaambre, T.; Samuel, J.L.; et al. Study of regulation of mitochondrial respiration in vivo. An analysis of influence of adp diffusion and possible role of cytoskeleton. Biochim. Biophys. Acta 1997, 1322, 41–59. [Google Scholar] [CrossRef]
- Bernier-Valentin, F.; Rousset, B. Interaction of tubulin with rat liver mitochondria. J. Biol. Chem. 1982, 257, 7092–7099. [Google Scholar] [PubMed]
- Carre, M.; Andre, N.; Carles, G.; Borghi, H.; Brichese, L.; Briand, C.; Braguer, D. Tubulin is an inherent component of mitochondrial membranes that interacts with the voltage-dependent anion channel. J. Biol. Chem. 2002, 277, 33664–33669. [Google Scholar] [CrossRef] [PubMed]
- Puurand, U.; Kadaja, L.; Seppet, E.K. Kindred DNA amplification from two distinct populations of cdna fragments. Biotechniques 2003, 34, 994–1000. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, K.; Monge, C.; Bruckner, A.; Puurand, U.; Kadaja, L.; Kaambre, T.; Seppet, E.; Saks, V. Study of possible interactions of tubulin, microtubular network, and stop protein with mitochondria in muscle cells. Mol. Cell. Biochem. 2010, 337, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Guzun, R.; Karu-Varikmaa, M.; Gonzalez-Granillo, M.; Kuznetsov, A.V.; Michel, L.; Cottet-Rousselle, C.; Saaremae, M.; Kaambre, T.; Metsis, M.; Grimm, M.; et al. Mitochondria-cytoskeleton interaction: Distribution of beta-tubulins in cardiomyocytes and hl-1 cells. Biochim. Biophys. Acta 2011, 1807, 458–469. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Granillo, M.; Grichine, A.; Guzun, R.; Usson, Y.; Tepp, K.; Chekulayev, V.; Shevchuk, I.; Karu-Varikmaa, M.; Kuznetsov, A.V.; Grimm, M.; et al. Studies of the role of tubulin beta ii isotype in regulation of mitochondrial respiration in intracellular energetic units in cardiac cells. J. Mol. Cell. Cardiol. 2012, 52, 437–447. [Google Scholar] [CrossRef]
- Kuznetsov, A.V.; Javadov, S.; Guzun, R.; Grimm, M.; Saks, V. Cytoskeleton and regulation of mitochondrial function: The role of beta-tubulin ii. Front. Physiol. 2013, 4, 82. [Google Scholar] [CrossRef]
- Schlattner, U.; Tokarska-Schlattner, M.; Wallimann, T. Mitochondrial creatine kinase in human health and disease. Biochim. Biophys. Acta 2006, 1762, 164–180. [Google Scholar] [CrossRef] [Green Version]
- Colombini, M. Vdac: The channel at the interface between mitochondria and the cytosol. Mol. Cell. Biochem. 2004, 256–257, 107–115. [Google Scholar] [CrossRef]
- Guzun, R.; Timohhina, N.; Tepp, K.; Gonzalez-Granillo, M.; Shevchuk, I.; Chekulayev, V.; Kuznetsov, A.V.; Kaambre, T.; Saks, V.A. Systems bioenergetics of creatine kinase networks: Physiological roles of creatine and phosphocreatine in regulation of cardiac cell function. Amino Acids 2011, 40, 1333–1348. [Google Scholar] [CrossRef]
- Burelle, Y.; Hochachka, P.W. Endurance training induces muscle-specific changes in mitochondrial function in skinned muscle fibers. J. Appl. Physiol. 2002, 92, 2429–2438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagur, R.; Tanguy, S.; Foriel, S.; Grichine, A.; Sanchez, C.; Pernet-Gallay, K.; Kaambre, T.; Kuznetsov, A.V.; Usson, Y.; Boucher, F.; et al. The impact of cardiac ischemia/reperfusion on the mitochondria-cytoskeleton interactions. Biochim. Biophys. Acta 2016, 1862, 1159–1171. [Google Scholar] [CrossRef] [PubMed]
- Saks, V.; Guzun, R.; Timohhina, N.; Tepp, K.; Varikmaa, M.; Monge, C.; Beraud, N.; Kaambre, T.; Kuznetsov, A.; Kadaja, L.; et al. Structure-function relationships in feedback regulation of energy fluxes in vivo in health and disease: Mitochondrial interactosome. Biochim. Biophys. Acta 2010, 1797, 678–697. [Google Scholar] [CrossRef] [PubMed]
- Anmann, T.; Guzun, R.; Beraud, N.; Pelloux, S.; Kuznetsov, A.V.; Kogerman, L.; Kaambre, T.; Sikk, P.; Paju, K.; Peet, N.; et al. Different kinetics of the regulation of respiration in permeabilized cardiomyocytes and in hl-1 cardiac cells. Importance of cell structure/organization for respiration regulation. Biochim. Biophys. Acta 2006, 1757, 1597–1606. [Google Scholar] [CrossRef] [PubMed]
- Monge, C.; Beraud, N.; Kuznetsov, A.V.; Rostovtseva, T.; Sackett, D.; Schlattner, U.; Vendelin, M.; Saks, V.A. Regulation of respiration in brain mitochondria and synaptosomes: Restrictions of adp diffusion in situ, roles of tubulin, and mitochondrial creatine kinase. Mol. Cell. Biochem. 2008, 318, 147–165. [Google Scholar] [CrossRef] [PubMed]
- Kaambre, T.; Chekulayev, V.; Shevchuk, I.; Tepp, K.; Timohhina, N.; Varikmaa, M.; Bagur, R.; Klepinin, A.; Anmann, T.; Koit, A.; et al. Metabolic control analysis of respiration in human cancer tissue. Front. Physiol. 2013, 4, 151. [Google Scholar] [CrossRef] [PubMed]
- Chekulayev, V.; Mado, K.; Shevchuk, I.; Koit, A.; Kaldma, A.; Klepinin, A.; Timohhina, N.; Tepp, K.; Kandashvili, M.; Ounpuu, L.; et al. Metabolic remodeling in human colorectal cancer and surrounding tissues: Alterations in regulation of mitochondrial respiration and metabolic fluxes. Biochem. Biophys. Rep. 2015, 4, 111–125. [Google Scholar] [CrossRef] [PubMed]
- Kaldma, A.; Klepinin, A.; Chekulayev, V.; Mado, K.; Shevchuk, I.; Timohhina, N.; Tepp, K.; Kandashvili, M.; Varikmaa, M.; Koit, A.; et al. An in situ study of bioenergetic properties of human colorectal cancer: The regulation of mitochondrial respiration and distribution of flux control among the components of atp synthasome. Int. J. Biochem. Cell Biol. 2014, 55, 171–186. [Google Scholar] [CrossRef]
- Koit, A.; Shevchuk, I.; Ounpuu, L.; Klepinin, A.; Chekulayev, V.; Timohhina, N.; Tepp, K.; Puurand, M.; Truu, L.; Heck, K.; et al. Mitochondrial respiration in human colorectal and breast cancer clinical material is regulated differently. Oxid. Med. Cell. Longev. 2017, 2017, 1372640. [Google Scholar] [CrossRef]
- Klepinin, A.; Ounpuu, L.; Mado, K.; Truu, L.; Chekulayev, V.; Puurand, M.; Shevchuk, I.; Tepp, K.; Planken, A.; Kaambre, T. The complexity of mitochondrial outer membrane permeability and vdac regulation by associated proteins. J. Bioenerg. Biomembr. 2018, 50, 339–354. [Google Scholar] [CrossRef]
- Klepinin, A.; Chekulayev, V.; Timohhina, N.; Shevchuk, I.; Tepp, K.; Kaldma, A.; Koit, A.; Saks, V.; Kaambre, T. Comparative analysis of some aspects of mitochondrial metabolism in differentiated and undifferentiated neuroblastoma cells. J. Bioenerg. Biomembr. 2014, 46, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Ounpuu, L.; Truu, L.; Shevchuk, I.; Chekulayev, V.; Klepinin, A.; Koit, A.; Tepp, K.; Puurand, M.; Rebane-Klemm, E.; Kaambre, T. Comparative analysis of the bioenergetics of human adenocarcinoma caco-2 cell line and postoperative tissue samples from colorectal cancer patients. Biochem. Cell Biol. 2018, 96, 808–817. [Google Scholar] [CrossRef] [PubMed]
- Blitz, A.L.; Fine, R.E. Muscle-like contractile proteins and tubulin in synaptosomes. Proc. Natl. Acad. Sci. USA 1974, 71, 4472–4476. [Google Scholar] [CrossRef] [PubMed]
- Cumming, R.; Burgoyne, R.D.; Lytton, N.A.; Gray, E.G. Immunocytochemical evidence for tubulin in the presynaptic terminal of synaptosomes. Neurosci. Lett. 1983, 37, 215–220. [Google Scholar] [CrossRef]
- Zisapel, N.; Levi, M.; Gozes, I. Tubulin: An integral protein of mammalian synaptic vesicle membranes. J. Neurochem. 1980, 34, 26–32. [Google Scholar] [CrossRef]
- Breuss, M.W.; Leca, I.; Gstrein, T.; Hansen, A.H.; Keays, D.A. Tubulins and brain development-the origins of functional specification. Mol. Cell. Neurosci. 2017, 84, 58–67. [Google Scholar] [CrossRef]
- Tischfield, M.A.; Engle, E.C. Distinct alpha- and beta-tubulin isotypes are required for the positioning, differentiation and survival of neurons: New support for the ‘multi-tubulin’ hypothesis. Biosci. Rep. 2010, 30, 319–330. [Google Scholar] [CrossRef]
- Wallimann, T.; Hemmer, W. Creatine kinase in non-muscle tissues and cells. Mol. Cell. Biochem. 1994, 133–134, 193–220. [Google Scholar] [CrossRef]
- Friedman, D.L.; Roberts, R. Compartmentation of brain-type creatine kinase and ubiquitous mitochondrial creatine kinase in neurons: Evidence for a creatine phosphate energy shuttle in adult rat brain. J. Comp. Neurol. 1994, 343, 500–511. [Google Scholar] [CrossRef]
- Warburg, O.; Wind, F.; Negelein, E. The metabolism of tumors in the body. J. Gen. Physiol. 1927, 8, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Warburg, O. On respiratory impairment in cancer cells. Science 1956, 124, 269–270. [Google Scholar] [PubMed]
- Lemasters, J.J.; Holmuhamedov, E. Voltage-dependent anion channel (vdac) as mitochondrial governator--thinking outside the box. Biochim. Biophys. Acta 2006, 1762, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Sanchez, R.; Saavedra, E.; Gallardo-Perez, J.C.; Rumjanek, F.D.; Rodriguez-Enriquez, S. Understanding the cancer cell phenotype beyond the limitations of current omics analyses. FEBS J. 2016, 283, 54–73. [Google Scholar] [CrossRef]
- Parker, A.L.; Teo, W.S.; McCarroll, J.A.; Kavallaris, M. An emerging role for tubulin isotypes in modulating cancer biology and chemotherapy resistance. Int. J. Mol. Sci. 2017, 18, 1434. [Google Scholar] [CrossRef] [PubMed]
- Kaambre, T.; Chekulayev, V.; Shevchuk, I.; Karu-Varikmaa, M.; Timohhina, N.; Tepp, K.; Bogovskaja, J.; Kutner, R.; Valvere, V.; Saks, V. Metabolic control analysis of cellular respiration in situ in intraoperational samples of human breast cancer. J. Bioenerg. Biomembr. 2012, 44, 539–558. [Google Scholar] [CrossRef]
- Mathupala, S.P.; Ko, Y.H.; Pedersen, P.L. Hexokinase-2 bound to mitochondria: Cancer’s stygian link to the “warburg effect” and a pivotal target for effective therapy. Semin. Cancer Biol. 2009, 19, 17–24. [Google Scholar] [CrossRef]
- Maldonado, E.N.; Sheldon, K.L.; DeHart, D.N.; Patnaik, J.; Manevich, Y.; Townsend, D.M.; Bezrukov, S.M.; Rostovtseva, T.K.; Lemasters, J.J. Voltage-dependent anion channels modulate mitochondrial metabolism in cancer cells: Regulation by free tubulin and erastin. J. Biol. Chem. 2013, 288, 11920–11929. [Google Scholar] [CrossRef]
- Pedersen, P.L. Warburg, me and hexokinase 2: Multiple discoveries of key molecular events underlying one of cancers’ most common phenotypes, the “warburg effect”, i.E., elevated glycolysis in the presence of oxygen. J. Bioenerg. Biomembr. 2007, 39, 211–222. [Google Scholar] [CrossRef]
- Maldonado, E.N.; Patnaik, J.; Mullins, M.R.; Lemasters, J.J. Free tubulin modulates mitochondrial membrane potential in cancer cells. Cancer Res. 2010, 70, 10192–10201. [Google Scholar] [CrossRef]
- Maldonado, E.N. Vdac-tubulin, an anti-warburg pro-oxidant switch. Front. Oncol. 2017, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Shen, H.; Zhang, L.G.; Liu, J.; Cao, X.G.; Yao, A.L.; Kang, S.S.; Gao, W.X.; Han, H.; Cao, F.H.; et al. Construction and analysis of protein-protein interaction networks based on proteomics data of prostate cancer. Int. J. Mol. Med. 2016, 37, 1576–1586. [Google Scholar] [CrossRef] [Green Version]
- Katsetos, C.D.; Legido, A.; Perentes, E.; Mork, S.J. Class iii beta-tubulin isotype: A key cytoskeletal protein at the crossroads of developmental neurobiology and tumor neuropathology. J. Child Neurol. 2003, 18, 851–866. [Google Scholar] [CrossRef] [PubMed]
- Jirasek, T.; Mandys, V.; Viklicky, V. Expression of class iii beta-tubulin in neuroendocrine tumours of gastrointestinal tract. Folia Histochem. Cytobiol. 2002, 40, 305–310. [Google Scholar] [PubMed]
- Urano, N.; Fujiwara, Y.; Doki, Y.; Kim, S.J.; Miyoshi, Y.; Noguchi, S.; Miyata, H.; Takiguchi, S.; Yasuda, T.; Yano, M.; et al. Clinical significance of class iii beta-tubulin expression and its predictive value for resistance to docetaxel-based chemotherapy in gastric cancer. Int. J. Oncol. 2006, 28, 375–381. [Google Scholar]
- Mariani, M.; Shahabi, S.; Sieber, S.; Scambia, G.; Ferlini, C. Class iii beta-tubulin (tubb3): More than a biomarker in solid tumors? Curr. Mol. Med. 2011, 11, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.O.; Kim, W.Y.; Go, H.; Paik, J.H.; Kim, J.E.; Kim, Y.A.; Huh, J.R.; Jeon, Y.K.; Kim, C.W. Class iii beta-tubulin shows unique expression patterns in a variety of neoplastic and non-neoplastic lymphoproliferative disorders. Am. J. Surg. Phatol. 2010, 34, 645–655. [Google Scholar] [CrossRef]
- Mozzetti, S.; Ferlini, C.; Concolino, P.; Filippetti, F.; Raspaglio, G.; Prislei, S.; Gallo, D.; Martinelli, E.; Ranelletti, F.O.; Ferrandina, G.; et al. Class iii beta-tubulin overexpression is a prominent mechanism of paclitaxel resistance in ovarian cancer patients. Clin. Cancer Res. 2005, 11, 298–305. [Google Scholar]
- Cucchiarelli, V.; Hiser, L.; Smith, H.; Frankfurter, A.; Spano, A.; Correia, J.J.; Lobert, S. Beta-tubulin isotype classes ii and v expression patterns in nonsmall cell lung carcinomas. Cell Motil. Cytoskelet. 2008, 65, 675–685. [Google Scholar] [CrossRef]
- Dumontet, C.; Isaac, S.; Souquet, P.J.; Bejui-Thivolet, F.; Pacheco, Y.; Peloux, N.; Frankfurter, A.; Luduena, R.; Perol, M. Expression of class iii beta tubulin in non-small cell lung cancer is correlated with resistance to taxane chemotherapy. Bull. Cancer 2005, 92, E25–E30. [Google Scholar]
- Cicchillitti, L.; Penci, R.; Di Michele, M.; Filippetti, F.; Rotilio, D.; Donati, M.B.; Scambia, G.; Ferlini, C. Proteomic characterization of cytoskeletal and mitochondrial class iii beta-tubulin. Mol. Cancer Ther. 2008, 7, 2070–2079. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Outschoorn, U.E.; Pestell, R.G.; Howell, A.; Tykocinski, M.L.; Nagajyothi, F.; Machado, F.S.; Tanowitz, H.B.; Sotgia, F.; Lisanti, M.P. Energy transfer in “parasitic” cancer metabolism: Mitochondria are the powerhouse and achilles’ heel of tumor cells. Cell Cycle 2011, 10, 4208–4216. [Google Scholar] [CrossRef] [PubMed]
- Monge, C.; Beraud, N.; Tepp, K.; Pelloux, S.; Chahboun, S.; Kaambre, T.; Kadaja, L.; Roosimaa, M.; Piirsoo, A.; Tourneur, Y.; et al. Comparative analysis of the bioenergetics of adult cardiomyocytes and nonbeating hl-1 cells: Respiratory chain activities, glycolytic enzyme profiles, and metabolic fluxes. Can. J. Physiol. Pharmacol. 2009, 87, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Klepinin, A.; Ounpuu, L.; Guzun, R.; Chekulayev, V.; Timohhina, N.; Tepp, K.; Shevchuk, I.; Schlattner, U.; Kaambre, T. Simple oxygraphic analysis for the presence of adenylate kinase 1 and 2 in normal and tumor cells. J. Bioenerg. Biomembr. 2016, 48, 531–548. [Google Scholar] [CrossRef] [PubMed]
- Noskov, S.Y.; Rostovtseva, T.K.; Bezrukov, S.M. Atp transport through vdac and the vdac-tubulin complex probed by equilibrium and nonequilibrium md simulations. Biochemistry 2013, 52, 9246–9256. [Google Scholar] [CrossRef] [PubMed]
- Rostovtseva, T.K.; Gurnev, P.A.; Chen, M.Y.; Bezrukov, S.M. Membrane lipid composition regulates tubulin interaction with mitochondrial voltage-dependent anion channel. J. Biol. Chem. 2012, 287, 29589–29598. [Google Scholar] [CrossRef] [PubMed]
- Puurand, M.; Tepp, K.; Klepinin, A.; Klepinina, L.; Shevchuk, I.; Kaambre, T. Intracellular energy-transfer networks and high-resolution respirometry: A convenient approach for studying their function. Int. J. Mol. Sci. 2018, 19, 2933. [Google Scholar] [CrossRef]
- Reina, S.; De Pinto, V. Anti-cancer compounds targeted to vdac: Potential and perspectives. Curr. Med. Chem. 2017, 24, 4447–4469. [Google Scholar] [CrossRef]
- Hoogerheide, D.P.; Noskov, S.Y.; Jacobs, D.; Bergdoll, L.; Silin, V.; Worcester, D.L.; Abramson, J.; Nanda, H.; Rostovtseva, T.K.; Bezrukov, S.M. Structural features and lipid binding domain of tubulin on biomimetic mitochondrial membranes. Proc. Natl. Acad. Sci. USA 2017, 114, E3622–E3631. [Google Scholar] [CrossRef]
- Draberova, E.; Sulimenko, V.; Vinopal, S.; Sulimenko, T.; Sladkova, V.; D’Agostino, L.; Sobol, M.; Hozak, P.; Kren, L.; Katsetos, C.D.; et al. Differential expression of human gamma-tubulin isotypes during neuronal development and oxidative stress points to a gamma-tubulin-2 prosurvival function. FASEB J. 2017, 31, 1828–1846. [Google Scholar] [CrossRef]
- Lindstrom, L.; Li, T.; Malycheva, D.; Kancharla, A.; Nilsson, H.; Vishnu, N.; Mulder, H.; Johansson, M.; Rossello, C.A.; Alvarado-Kristensson, M. The gtpase domain of gamma-tubulin is required for normal mitochondrial function and spatial organization. Commun. Biol. 2018, 1, 37. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Tissue | Apparent KmADP 1, µM | References |
|---|---|---|
| Rat heart mitochondria | 10–20 | [12,21,23,26,31] |
| Rat heart fibers | 300–500 | [21,23,25,26,27,31] |
| Rat heart isolated cardiomyocytes | 300–500 | [13,14,38,46] |
| Trypsin-treated rat heart fibers | 100–110 | [21,23] |
| Rat heart fibers in the presence of creatine | 80–100 | [21,25,26,31] |
| Rat m. soleus | 300-400 | [21,30] |
| Rat m. gastrocnemius white | 5–15 | [21,30] |
| Rat brain mitochondria | 10 | [47] |
| Rat brain synaptosomes | 110 | [47] |
| Rat brain synaptosomes in the presence of creatine | 25 | [47] |
| Human colorectal cancer | 90–130 | [48,49,50] |
| Human breast cancer | ||
| Mitochondrial population I | 45 | [51] |
| Mitochondrial population II | 300 | [51] |
| Cell lines | ||
| Mouse neuroblastoma N2a | 20–40 | [52,53] |
| Mouse sarcoma HL-1 | 25–50 | [46] |
| Human colorectal cancer Caco-2 | 40 | [54] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puurand, M.; Tepp, K.; Timohhina, N.; Aid, J.; Shevchuk, I.; Chekulayev, V.; Kaambre, T. Tubulin βII and βIII Isoforms as the Regulators of VDAC Channel Permeability in Health and Disease. Cells 2019, 8, 239. https://doi.org/10.3390/cells8030239
Puurand M, Tepp K, Timohhina N, Aid J, Shevchuk I, Chekulayev V, Kaambre T. Tubulin βII and βIII Isoforms as the Regulators of VDAC Channel Permeability in Health and Disease. Cells. 2019; 8(3):239. https://doi.org/10.3390/cells8030239
Chicago/Turabian StylePuurand, Marju, Kersti Tepp, Natalja Timohhina, Jekaterina Aid, Igor Shevchuk, Vladimir Chekulayev, and Tuuli Kaambre. 2019. "Tubulin βII and βIII Isoforms as the Regulators of VDAC Channel Permeability in Health and Disease" Cells 8, no. 3: 239. https://doi.org/10.3390/cells8030239
APA StylePuurand, M., Tepp, K., Timohhina, N., Aid, J., Shevchuk, I., Chekulayev, V., & Kaambre, T. (2019). Tubulin βII and βIII Isoforms as the Regulators of VDAC Channel Permeability in Health and Disease. Cells, 8(3), 239. https://doi.org/10.3390/cells8030239