Microtubular and Nuclear Functions of γ-Tubulin: Are They LINCed?



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Functions of γ-Tubulin with Microtubules and Its Interaction with Other Cytoskeletal Systems
3. γ-Tubulin Has an Intrinsic Ability to Assemble Filaments with a Bundling and Aggregating Capacity
4. Functions of γ-Tubulin in the Cell Cycle Regulation and in Nuclear Processes
5. γ-Tubulin Interacts with Lamin and the Interaction Is Required for Regular Nuclei Organization
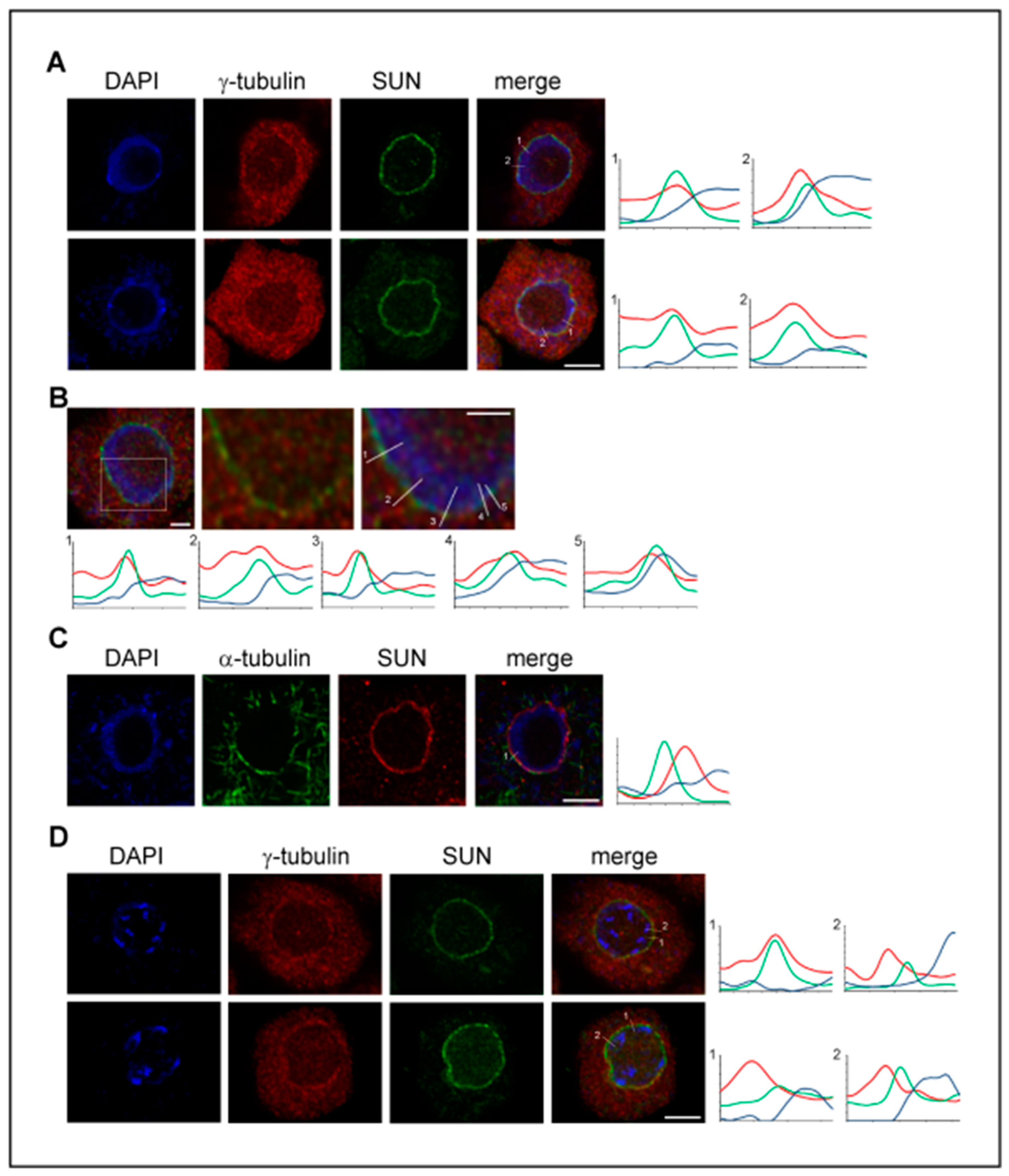
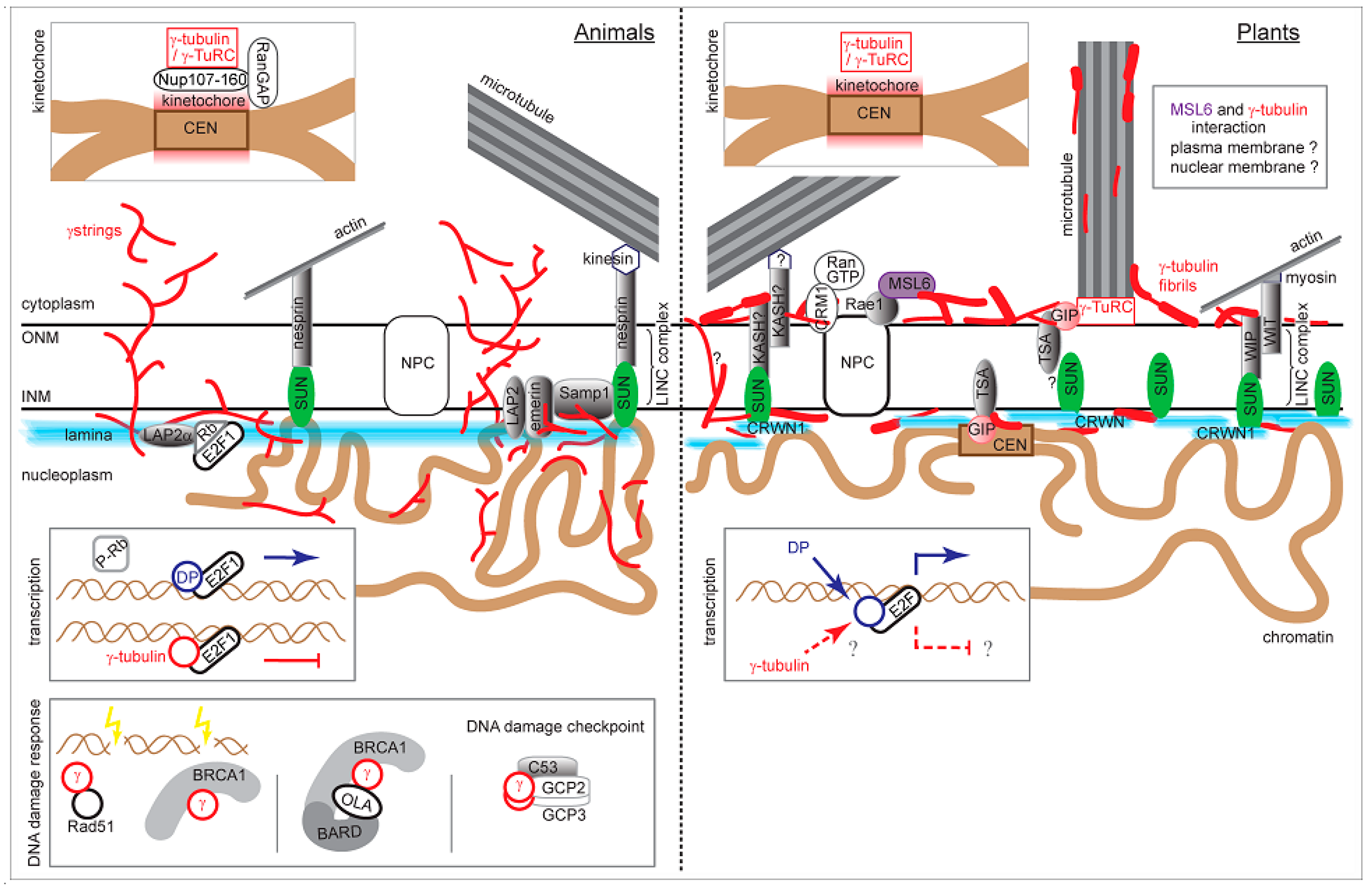
6. γ-Tubulin, Proteins of the Nuclear Envelope and LINC Complexes
7. γ-Tubulin and Mechanotransduction
8. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CM-SR | confocal laser scanning microscopy equipped with a super-resolution module |
| LINC | Linker of Nucleoskeleton and Cytoskeleton |
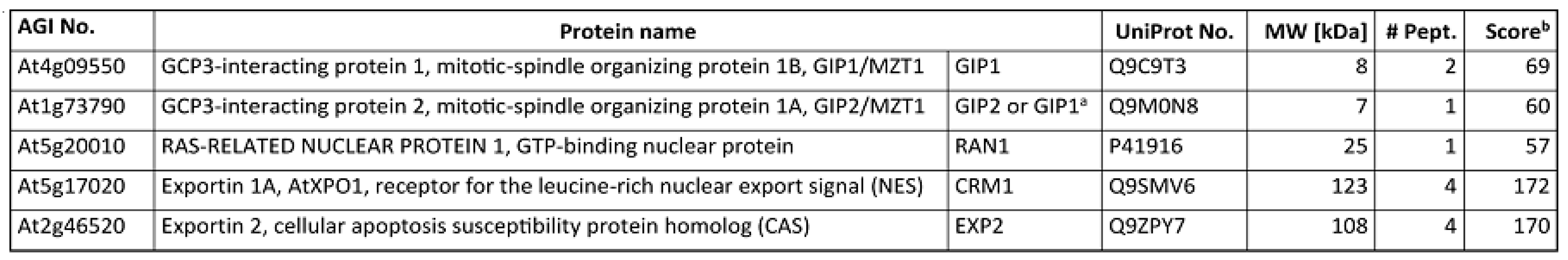
Appendix A. Co-Purification of GIP Proteins and Exportins with γ-Tubulin

Appendix B. Fluorescence Microscopy
References
- Pilhofer, M.; Ladinsky, M.S.; McDowall, A.W.; Petroni, G.; Jensen, G.J. Microtubules in bacteria: Ancient tubulins build a five-protofilament homolog of the eukaryotic cytoskeleton. PLoS Biol. 2011, 9, e1001213. [Google Scholar] [CrossRef] [PubMed]
- Chumova, J.; Trogelova, L.; Kourova, H.; Volc, J.; Sulimenko, V.; Halada, P.; Kucera, O.; Benada, O.; Kucharova, A.; Klebanovych, A.; et al. gamma-Tubulin has a conserved intrinsic property of self-polymerization into double stranded filaments and fibrillar networks. Biochim. Biophys. Acta 2018, 1865, 734–748. [Google Scholar] [CrossRef] [PubMed]
- Yutin, N.; Koonin, E.V. Archaeal origin of tubulin. Biol. Direct 2012, 7, 10. [Google Scholar] [CrossRef] [PubMed]
- Janke, C. The tubulin code: Molecular components, readout mechanisms, and functions. J. Cell Biol. 2014, 206, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Vinopal, S.; Cernohorska, M.; Sulimenko, V.; Sulimenko, T.; Vosecka, V.; Flemr, M.; Draberova, E.; Draber, P. gamma-Tubulin 2 nucleates microtubules and is downregulated in mouse early embryogenesis. PloS ONE 2012, 7, e29919. [Google Scholar] [CrossRef]
- Oegema, K.; Wiese, C.; Martin, O.C.; Milligan, R.A.; Iwamatsu, A.; Mitchison, T.J.; Zheng, Y. Characterization of two related Drosophila gamma-tubulin complexes that differ in their ability to nucleate microtubules. J. Cell Biol. 1999, 144, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Yuba-Kubo, A.; Kubo, A.; Hata, M.; Tsukita, S. Gene knockout analysis of two gamma-tubulin isoforms in mice. Dev. Biol. 2005, 282, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Draberova, E.; Sulimenko, V.; Vinopal, S.; Sulimenko, T.; Sladkova, V.; D’Agostino, L.; Sobol, M.; Hozak, P.; Kren, L.; Katsetos, C.D.; et al. Differential expression of human gamma-tubulin isotypes during neuronal development and oxidative stress points to a gamma-tubulin-2 prosurvival function. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2017, 31, 1828–1846. [Google Scholar] [CrossRef]
- Sulimenko, V.; Sulimenko, T.; Poznanovic, S.; Nechiporuk-Zloy, V.; Bohm, K.J.; Macurek, L.; Unger, E.; Draber, P. Association of brain gamma-tubulins with alpha beta-tubulin dimers. Biochem. J. 2002, 365, 889–895. [Google Scholar] [CrossRef]
- Detraves, C.; Mazarguil, H.; Lajoie-Mazenc, I.; Julian, M.; Raynaud-Messina, B.; Wright, M. Protein complexes containing gamma-tubulin are present in mammalian brain microtubule protein preparations. Cell Motil. Cytoskeleton 1997, 36, 179–189. [Google Scholar] [CrossRef]
- Moudjou, M.; Bordes, N.; Paintrand, M.; Bornens, M. gamma-Tubulin in mammalian cells: The centrosomal and the cytosolic forms. J. Cell Sci. 1996, 109, 875–887. [Google Scholar] [PubMed]
- Stumpff, J.; Kellogg, D.R.; Krohne, K.A.; Su, T.T. Drosophila Wee1 interacts with members of the gammaTURC and is required for proper mitotic-spindle morphogenesis and positioning. Curr. Biol. 2005, 15, 1525–1534. [Google Scholar] [CrossRef] [PubMed]
- Vogel, J.; Drapkin, B.; Oomen, J.; Beach, D.; Bloom, K.; Snyder, M. Phosphorylation of gamma-tubulin regulates microtubule organization in budding yeast. Dev. Cell 2001, 1, 621–631. [Google Scholar] [CrossRef]
- Starita, L.M.; Machida, Y.; Sankaran, S.; Elias, J.E.; Griffin, K.; Schlegel, B.P.; Gygi, S.P.; Parvin, J.D. BRCA1-dependent ubiquitination of gamma-tubulin regulates centrosome number. Mol. Cell. Biol. 2004, 24, 8457–8466. [Google Scholar] [CrossRef]
- Lee, Y.J.; Liu, B. Microtubule nucleation for the assembly of acentrosomal microtubule arrays in plant cells. New Phytol. 2019. [Google Scholar] [CrossRef]
- Roostalu, J.; Surrey, T. Microtubule nucleation: Beyond the template. Nat. Rev. Mol. Cell Biol. 2017, 18, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Erickson, H.P.; Stoffler, D. Protofilaments and rings, two conformations of the tubulin family conserved from bacterial FtsZ to alpha/beta and gamma tubulin. J. Cell Biol. 1996, 135, 5–8. [Google Scholar] [CrossRef] [Green Version]
- Pastuglia, M.; Azimzadeh, J.; Goussot, M.; Camilleri, C.; Belcram, K.; Evrard, J.L.; Schmit, A.C.; Guerche, P.; Bouchez, D. Gamma-tubulin is essential for microtubule organization and development in Arabidopsis. Plant Cell 2006, 18, 1412–1425. [Google Scholar] [CrossRef]
- Binarova, P.; Cenklova, V.; Prochazkova, J.; Doskocilova, A.; Volc, J.; Vrlik, M.; Bogre, L. Gamma-tubulin is essential for acentrosomal microtubule nucleation and coordination of late mitotic events in Arabidopsis. Plant Cell 2006, 18, 1199–1212. [Google Scholar] [CrossRef] [PubMed]
- Kong, Z.; Hotta, T.; Lee, Y.R.; Horio, T.; Liu, B. The gamma-tubulin complex protein GCP4 is required for organizing functional microtubule arrays in Arabidopsis thaliana. Plant Cell 2010, 22, 191–204. [Google Scholar] [CrossRef]
- Nakamura, M.; Yagi, N.; Kato, T.; Fujita, S.; Kawashima, N.; Ehrhardt, D.W.; Hashimoto, T. Arabidopsis GCP3-interacting protein 1/MOZART 1 is an integral component of the gamma-tubulin-containing microtubule nucleating complex. Plant J. Cell Mol. Biol. 2012, 71, 216–225. [Google Scholar] [CrossRef]
- Binarova, P.; Hause, B.; Dolezel, J.; Draber, P. Association of gamma-tubulin with kinetochore/centromeric region of plant chromosomes. Plant J. 1998, 14, 751–757. [Google Scholar] [CrossRef] [Green Version]
- Mishra, R.K.; Chakraborty, P.; Arnaoutov, A.; Fontoura, B.M.A.; Dasso, M. The Nup107-160 complex and gamma-TuRC regulate microtubule polymerization at kinetochores. Nat. Cell Biol. 2010, 12, 164–169. [Google Scholar] [CrossRef]
- Meunier, S.; Vernos, I. Acentrosomal Microtubule Assembly in Mitosis: The Where, When, and How. Trends Cell Biol. 2016, 26, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.R.J.; Hiwatashi, Y.; Hotta, T.; Xie, T.T.; Doonan, J.H.; Liu, B. The Mitotic Function of Augmin Is Dependent on Its Microtubule-Associated Protein Subunit EDE1 in Arabidopsis thaliana. Curr. Biol. 2017, 27, 3891–3897. [Google Scholar] [CrossRef]
- Goshima, G.; Mayer, M.; Zhang, N.; Stuurman, N.; Vale, R.D. Augmin: A protein complex required for centrosome-independent microtubule generation within the spindle. J. Cell Biol. 2008, 181, 421–429. [Google Scholar] [CrossRef]
- Samejima, I.; Lourenco, P.C.; Snaith, H.A.; Sawin, K.E. Fission yeast mto2p regulates microtubule nucleation by the centrosomin-related protein mto1p. Mol. Biol. Cell 2005, 16, 3040–3051. [Google Scholar] [CrossRef]
- Goshima, G.; Kimura, A. New look inside the spindle: Microtubule-dependent microtubule generation within the spindle. Curr. Opin. Cell Biol. 2010, 22, 44–49. [Google Scholar] [CrossRef]
- Liu, T.; Tian, J.; Wang, G.D.; Yu, Y.J.; Wang, C.F.; Ma, Y.P.; Zhang, X.X.; Xia, G.X.; Liu, B.; Kong, Z.S. Augmin Triggers Microtubule-Dependent Microtubule Nucleation in Interphase Plant Cells. Curr. Biol. 2014, 24, 2708–2713. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wu, T.; Shi, L.; Zhang, L.; Zheng, W.; Qu, J.Y.; Niu, R.; Qi, R.Z. Conserved motif of CDK5RAP2 mediates its localization to centrosomes and the Golgi complex. J. Biol. Chem. 2010, 285, 22658–22665. [Google Scholar] [CrossRef]
- Chabin-Brion, K.; Marceiller, J.; Perez, F.; Settegrana, C.; Drechou, A.; Durand, G.; Pous, C. The golgi complex is a microtubule-organizing organelle. Mol. Biol. Cell 2001, 12, 2047–2060. [Google Scholar] [CrossRef]
- Drykova, D.; Cenklova, V.; Sulimenko, V.; Volc, J.; Draber, P.; Binarova, P. Plant gamma-tubulin interacts with alphabeta-tubulin dimers and forms membrane-associated complexes. Plant Cell 2003, 15, 465–480. [Google Scholar] [CrossRef]
- Bao, X.X.; Spanos, C.; Kojidani, T.; Lynch, E.M.; Rappsilber, J.; Hiraoka, Y.; Haraguchi, T.; Sawin, K.E. Exportin Crm1 is repurposed as a docking protein to generate microtubule organizing centers at the nuclear pore. Elife 2018, 7, 34. [Google Scholar] [CrossRef]
- Shen, Y.; Wang, K.; Qi, R.Z. The catalytic subunit of DNA polymerase delta is a nucleocytoplasmic shuttling protein. Exp. Cell Res. 2019. [Google Scholar] [CrossRef]
- Shen, Y.H.; Liu, P.F.; Jiang, T.L.; Hu, Y.; Au, F.K.C.; Qi, R.Z. The catalytic subunit of DNA polymerase delta inhibits gamma TuRC activity and regulates Golgi-derived microtubules. Nat. Commun. 2017, 8, 13. [Google Scholar] [CrossRef]
- Petrovska, B.; Jerabkova, H.; Kohoutova, L.; Cenklova, V.; Pochylova, Z.; Gelova, Z.; Kocarova, G.; Vachova, L.; Kurejova, M.; Tomastikova, E.; et al. Overexpressed TPX2 causes ectopic formation of microtubular arrays in the nuclei of acentrosomal plant cells. J. Exp. Bot. 2013, 64, 4575–4587. [Google Scholar] [CrossRef] [Green Version]
- Cuschieri, L.; Miller, R.; Vogel, J. Gamma-tubulin is required for proper recruitment and assembly of Kar9-Bim1 complexes in budding yeast. Mol. Biol. Cell 2006, 17, 4420–4434. [Google Scholar] [CrossRef]
- Raynaud-Messina, B.; Merdes, A. Gamma-tubulin complexes and microtubule organization. Curr. Opin. Cell Biol. 2007, 19, 24–30. [Google Scholar] [CrossRef]
- Hubert, T.; Perdu, S.; Vandekerckhove, J.; Gettemans, J. gamma-Tubulin localizes at actin-based membrane protrusions and inhibits formation of stress-fibers. Biochem. Biophys. Res. Commun. 2011, 408, 248–252. [Google Scholar] [CrossRef]
- Oriolo, A.S.; Wald, F.A.; Canessa, G.; Salas, P.J. GCP6 binds to intermediate filaments: A novel function of keratins in the organization of microtubules in epithelial cells. Mol. Biol. Cell 2007, 18, 781–794. [Google Scholar] [CrossRef]
- Rossello, C.A.; Lindstrom, L.; Glindre, J.; Eklund, G.; Alvarado-Kristensson, M. Gamma-tubulin coordinates nuclear envelope assembly around chromatin. Heliyon 2016, 2, e00166. [Google Scholar] [CrossRef]
- Rios, R.M. The centrosome-Golgi apparatus nexus. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2014, 369, 20130462. [Google Scholar] [CrossRef]
- Lindström, L.; Li, T.; Malycheva, D.; Kancharla, A.; Nilsson, H.; Vishnu, N.; Mulder, H.; Johansson, M.; Rosselló, C.A.; Alvarado-Kristensson, M. The GTPase domain of gamma-tubulin is required for normal mitochondrial function and spatial organization. Commun. Biol. 2018, 1, 37. [Google Scholar] [CrossRef]
- Hehnly, H.; Doxsey, S. Rab11 Endosomes Contribute to Mitotic Spindle Organization and Orientation. Dev. Cell 2014, 28, 497–507. [Google Scholar] [CrossRef]
- Lindstrom, L.; Alvarado-Kristensson, M. Characterization of gamma-tubulin filaments in mammalian cells. Biochim. Biophys. Acta-Mol. Cell Res. 2018, 1865, 158–171. [Google Scholar] [CrossRef]
- Melki, R.; Vainberg, I.E.; Chow, R.L.; Cowan, N.J. Chaperonin-mediated folding of vertebrate actin-related protein and gamma-tubulin. J. Cell Biol. 1993, 122, 1301–1310. [Google Scholar] [CrossRef] [Green Version]
- Inclan, Y.F.; Nogales, E. Structural models for the self-assembly and microtubule interactions of gamma-, delta- and epsilon-tubulin. J. Cell Sci. 2001, 114, 413–422. [Google Scholar]
- Vassilev, A.; Kimble, M.; Silflow, C.D.; LaVoie, M.; Kuriyama, R. Identification of intrinsic dimer and overexpressed monomeric forms of gamma-tubulin in Sf9 cells infected with baculovirus containing the Chlamydomonas gamma-tubulin sequence. J. Cell Sci. 1995, 108, 1083–1092. [Google Scholar]
- Aldaz, H.; Rice, L.M.; Stearns, T.; Agard, D.A. Insights into microtubule nucleation from the crystal structure of human gamma-tubulin. Nature 2005, 435, 523–527. [Google Scholar] [CrossRef]
- Leguy, R.; Melki, R.; Pantaloni, D.; Carlier, M.F. Monomeric gamma-tubulin nucleates microtubules. J. Biol. Chem. 2000, 275, 21975–21980. [Google Scholar] [CrossRef]
- Pouchucq, L.; Lobos-Ruiz, P.; Araya, G.; Valpuesta, J.M.; Monasterio, O. The chaperonin CCT promotes the formation of fibrillar aggregates of gamma-tubulin. BBA-Proteins Proteom. 2018, 1866, 519–526. [Google Scholar] [CrossRef]
- Pilhofer, M.; Jensen, G.J. The bacterial cytoskeleton: More than twisted filaments. Curr. Opin. Cell Biol. 2013, 25, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Ghosal, D.; Lowe, J. Collaborative protein filaments. EMBO J. 2015, 34, 2312–2320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strauss, M.P.; Liew, A.T.; Turnbull, L.; Whitchurch, C.B.; Monahan, L.G.; Harry, E.J. 3D-SIM super resolution microscopy reveals a bead-like arrangement for FtsZ and the division machinery: Implications for triggering cytokinesis. PLoS Biol. 2012, 10, e1001389. [Google Scholar] [CrossRef] [PubMed]
- Kollman, J.M.; Polka, J.K.; Zelter, A.; Davis, T.N.; Agard, D.A. Microtubule nucleating gamma-TuSC assembles structures with 13-fold microtubule-like symmetry. Nature 2010, 466, 879–882. [Google Scholar] [CrossRef] [PubMed]
- Noble, J.W.; Hunter, D.V.; Roskelley, C.D.; Chan, E.K.L.; Mills, J. Loukoumasomes Are Distinct Subcellular Structures from Rods and Rings and Are Structurally Associated with MAP2 and the Nuclear Envelope in Retinal Cells. PLoS ONE 2016, 11, e0165162. [Google Scholar] [CrossRef]
- Hendrickson, T.W.; Yao, J.; Bhadury, S.; Corbett, A.H.; Joshi, H.C. Conditional Mutations in gamma-Tubulin Reveal Its Involvement in Chromosome Segregation and Cytokinesis. Mol. Biol. Cell 2001, 12, 2469–2481. [Google Scholar] [CrossRef]
- Prigozhina, N.L.; Oakley, C.E.; Lewis, A.M.; Nayak, T.; Osmani, S.A.; Oakley, B.R. Gamma-tubulin plays an essential role in the coordination of mitotic events. Mol. Biol. Cell 2004, 15, 1374–1386. [Google Scholar] [CrossRef] [PubMed]
- Nayak, T.; Edgerton-Morgan, H.; Horio, T.; Xiong, Y.; De Souza, C.P.; Osmani, S.A.; Oakley, B.R. Gamma-tubulin regulates the anaphase-promoting complex/cyclosome during interphase. J. Cell Biol. 2010, 190, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Binarova, P.; Cenklova, V.; Hause, B.; Kubatova, E.; Lysak, M.; Dolezel, J.; Bogre, L.; Draber, P. Nuclear gamma-tubulin during acentriolar plant mitosis. Plant Cell 2000, 12, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Lesca, C.; Germanier, M.; Raynaud-Messina, B.; Pichereaux, C.; Etievant, C.; Emond, S.; Burlet-Schiltz, O.; Monsarrat, B.; Wright, M.; Defais, M. DNA damage induce gamma-tubulin-RAD51 nuclear complexes in mammalian cells. Oncogene 2005, 24, 5165–5172. [Google Scholar] [CrossRef] [PubMed]
- Arquint, C.; Gabryjonczyk, A.M.; Nigg, E.A. Centrosomes as signalling centres. Philos. Trans. R. Soc. B-Biol. Sci. 2014, 369, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Hemmerich, P.; Grosse, F. Centrosomal localization of DNA damage checkpoint proteins. J. Cell. Biochem. 2007, 101, 451–465. [Google Scholar] [CrossRef]
- Matsuzawa, A.; Kanno, S.; Nakayama, M.; Mochiduki, H.; Wei, L.; Shimaoka, T.; Furukawa, Y.; Kato, K.; Shibata, S.; Yasui, A.; et al. The BRCA1/BARD1-interacting protein OLA1 functions in centrosome regulation. Mol. Cell 2014, 53, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Hubert, T.; Vandekerckhove, J.; Gettemans, J. Cdk1 and BRCA1 target gamma-tubulin to microtubule domains. Biochem. Biophys. Res. Commun. 2011, 414, 240–245. [Google Scholar] [CrossRef]
- Horejsi, B.; Vinopal, S.; Sladkova, V.; Draberova, E.; Sulimenko, V.; Sulimenko, T.; Vosecka, V.; Philimonenko, A.; Hozak, P.; Katsetos, C.D.; et al. Nuclear gamma-tubulin associates with nucleoli and interacts with tumor suppressor protein C53. J. Cell Physiol. 2012, 227, 367–382. [Google Scholar] [CrossRef]
- Mullee, L.I.; Morrison, C.G. Centrosomes in the DNA damage response--the hub outside the centre. Chromosome Res. 2016, 24, 35–51. [Google Scholar] [CrossRef]
- Vazquez, M.; Cooper, M.T.; Zurita, M.E.; Kennison, J.A. gammaTub23C interacts genetically with Brahma chromatin-remodeling complexes in Drosophila melanogaster. Genetics 2008, 180, 835–843. [Google Scholar] [CrossRef]
- Hoog, G.; Zarrizi, R.; von Stedingk, K.; Jonsson, K.; Alvarado-Kristensson, M. Nuclear localization of gamma-tubulin affects E2F transcriptional activity and S-phase progression. FASEB J. 2011, 25, 3815–3827. [Google Scholar] [CrossRef]
- Kohoutova, L.; Kourova, H.; Nagy, S.K.; Volc, J.; Halada, P.; Meszaros, T.; Meskiene, I.; Bogre, L.; Binarova, P. The Arabidopsis mitogen-activated protein kinase 6 is associated with gamma-tubulin on microtubules, phosphorylates EB1c and maintains spindle orientation under nitrosative stress. New Phytol. 2015, 207, 1061–1074. [Google Scholar] [CrossRef]
- Katsetos, C.D.; Draberova, E.; Legido, A.; Draber, P. Tubulin Targets in the Pathobiology and Therapy of Glioblastoma Multiforme. II. gamma-Tubulin. J. Cell. Physiol. 2009, 221, 514–520. [Google Scholar] [CrossRef] [PubMed]
- Katsetos, C.D.; Draberova, E.; Smejkalova, B.; Reddy, G.; Bertrand, L.; de Chadarevian, J.P.; Legido, A.; Nissanov, J.; Baas, P.W.; Draber, P. Class III beta-tubulin and gamma-tubulin are co-expressed and form complexes in human glioblastoma cells. Neurochem. Res. 2007, 32, 1387–1398. [Google Scholar] [CrossRef]
- Ehlen, A.; Rossello, C.A.; von Stedingk, K.; Hoog, G.; Nilsson, E.; Pettersson, H.M.; Jirstrom, K.; Alvarado-Kristensson, M. Tumors with nonfunctional retinoblastoma protein are killed by reduced gamma-tubulin levels. J. Biol. Chem. 2012, 287, 17241–17247. [Google Scholar] [CrossRef]
- Alvarado-Kristensson, M. gamma-tubulin as a signal-transducing molecule and meshwork with therapeutic potential. Signal Transduct. Target. Ther. 2018, 3, 6. [Google Scholar] [CrossRef]
- Woodruff, J.B.; Wueseke, O.; Viscardi, V.; Mahamid, J.; Ochoa, S.D.; Bunkenborg, J.; Widlund, P.O.; Pozniakovsky, A.; Zanin, E.; Bahmanyar, S.; et al. Centrosomes. Regulated assembly of a supramolecular centrosome scaffold in vitro. Science 2015, 348, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Ramer, M.S.; Cruz Cabrera, M.A.; Alan, N.; Scott, A.L.; Inskip, J.A. A new organellar complex in rat sympathetic neurons. PLoS ONE 2010, 5, e10872. [Google Scholar] [CrossRef]
- Serebryannyy, L.; Misteli, T. Protein sequestration at the nuclear periphery as a potential regulatory mechanism in premature aging. J. Cell Biol. 2018, 217, 21–37. [Google Scholar] [CrossRef]
- Ciska, M.; Moreno Diaz de la Espina, S. The intriguing plant nuclear lamina. Front. Plant Sci. 2014, 5, 166. [Google Scholar] [CrossRef]
- Fiserova, J.; Kiseleva, E.; Goldberg, M.W. Nuclear envelope and nuclear pore complex structure and organization in tobacco BY-2 cells. Plant J. 2009, 59, 243–255. [Google Scholar] [CrossRef] [Green Version]
- Crisp, M.; Liu, Q.; Roux, K.; Rattner, J.B.; Shanahan, C.; Burke, B.; Stahl, P.D.; Hodzic, D. Coupling of the nucleus and cytoplasm: Role of the LINC complex. J. Cell Biol. 2006, 172, 41–53. [Google Scholar] [CrossRef]
- Gundersen, G.G.; Worman, H.J. Nuclear positioning. Cell 2013, 152, 1376–1389. [Google Scholar] [CrossRef] [PubMed]
- Haque, F.; Lloyd, D.J.; Smallwood, D.T.; Dent, C.L.; Shanahan, C.M.; Fry, A.M.; Trembath, R.C.; Shackleton, S. SUN1 interacts with nuclear lamin A and cytoplasmic nesprins to provide a physical connection between the nuclear lamina and the cytoskeleton. Mol. Cell. Biol. 2006, 26, 3738–3751. [Google Scholar] [CrossRef]
- Patel, J.T.; Bottrill, A.; Prosser, S.L.; Jayaraman, S.; Straatman, K.; Fry, A.M.; Shackleton, S. Mitotic phosphorylation of SUN1 loosens its connection with the nuclear lamina while the LINC complex remains intact. Nucleus 2014, 5, 462–473. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.C.; Lei, K.; Yuan, X.B.; Wu, X.H.; Zhuang, Y.; Xu, T.; Xu, R.; Han, M. SUN1/2 and Syne/Nesprin-1/2 Complexes Connect Centrosome to the Nucleus during Neurogenesis and Neuronal Migration in Mice. Neuron 2009, 64, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.J.; Antoku, S.; Gundersen, G.G. Centrifugal Displacement of Nuclei Reveals Multiple LINC Complex Mechanisms for Homeostatic Nuclear Positioning. Curr. Biol. 2017, 27, 3097–3110. [Google Scholar] [CrossRef] [PubMed]
- Calvi, A.; Wong, A.S.W.; Wright, G.; Wong, E.S.M.; Loo, T.H.; Stewart, C.L.; Burke, B. SUN4 is essential for nuclear remodeling during mammalian spermiogenesis. Dev. Biol. 2015, 407, 321–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, K.; Zhu, X.Q.; Xu, R.; Shao, C.L.; Xu, T.; Zhuang, Y.; Han, M. Inner Nuclear Envelope Proteins SUN1 and SUN2 Play a Prominent Role in the DNA Damage Response. Curr. Biol. 2012, 22, 1609–1615. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Katsuyama, S.; Tateho, K.; Nakamura, H.; Miyoshi, J.; Ohba, T.; Matsuhara, H.; Miki, F.; Okazaki, K.; Haraguchi, T.; et al. Microtubule-organizing center formation at telomeres induces meiotic telomere clustering. J. Cell Biol. 2013, 200, 385–395. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.; Xu, R.; Yu, J.H.; Xu, T.; Zhuang, Y.; Han, M. SUN1 is required for telomere attachment to nuclear envelope and gametogenesis in mice. Dev. Cell 2007, 12, 863–872. [Google Scholar] [CrossRef]
- Varas, J.; Graumann, K.; Osman, K.; Pradillo, M.; Evans, D.E.; Santos, J.L.; Armstrong, S.J. Absence of SUN1 and SUN2 proteins in Arabidopsis thaliana leads to a delay in meiotic progression and defects in synapsis and recombination. Plant J. 2015, 81, 329–346. [Google Scholar] [CrossRef]
- Murphy, S.P.; Gumber, H.K.; Mao, Y.; Bass, H.W. A dynamic meiotic SUN belt includes the zygotene-stage telomere bouquet and is disrupted in chromosome segregation mutants of maize (Zea mays L.). Front. Plant Sci. 2014, 5, 11. [Google Scholar] [CrossRef] [PubMed]
- Oda, Y.; Fukuda, H. Dynamics of Arabidopsis SUN proteins during mitosis and their involvement in nuclear shaping. Plant J. 2011, 66, 629–641. [Google Scholar] [CrossRef]
- Graumann, K.; Runions, J.; Evans, D.E. Characterization of SUN-domain proteins at the higher plant nuclear envelope. Plant J. 2010, 61, 134–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Groves, N.R.; Meier, I. Plant nuclear shape is independently determined by the SUN-WIP-WIT2-myosin XI-i complex and CRWN1. Nucleus 2015, 6, 144–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groves, N.R.; Biel, A.M.; Newman-Griffis, A.H.; Meier, I. Dynamic Changes in Plant Nuclear Organization in Response to Environmental and Developmental Signals. Plant Physiol. 2018, 176, 230–241. [Google Scholar] [CrossRef]
- Meier, I. LINCing the eukaryotic tree of life—Towards a broad evolutionary comparison of nucleocytoplasmic bridging complexes. J. Cell Sci. 2016, 129, 3523–3531. [Google Scholar] [CrossRef]
- Graumann, K. Evidence for LINC1-SUN associations at the plant nuclear periphery. PLoS ONE 2014, 9, e93406. [Google Scholar] [CrossRef] [PubMed]
- Pawar, V.; Poulet, A.; Detourne, G.; Tatout, C.; Vanrobays, E.; Evans, D.E.; Graumann, K. A novel family of plant nuclear envelope-associated proteins. J. Exp. Bot. 2016, 67, 5699–5710. [Google Scholar] [CrossRef]
- Gimpel, P.; Lee, Y.L.; Sobota, R.M.; Calvi, A.; Koullourou, V.; Patel, R.; Mamchaoui, K.; Nedelec, F.; Shackleton, S.; Schmoranzer, J.; et al. Nesprin-1alpha-Dependent Microtubule Nucleation from the Nuclear Envelope via Akap450 Is Necessary for Nuclear Positioning in Muscle Cells. Curr. Biol. 2017, 27, 2999.e9–3009.e9. [Google Scholar] [CrossRef]
- Larsson, V.J. Characterization of the Inner Nuclear Membrane Protein Samp1, during Interphase and Mitosis. Ph.D. Thesis, Stockholm University, Stockholm, Sweden, 2018. [Google Scholar]
- Larsson, V.J.; Jafferali, M.H.; Vijayaraghavan, B.; Figueroa, R.A.; Hallberg, E. Mitotic spindle assembly and gamma-tubulin localisation depend on the integral nuclear membrane protein Samp1. J. Cell Sci. 2018, 131. [Google Scholar] [CrossRef]
- Teixido-Travesa, N.; Villen, J.; Lacasa, C.; Bertran, M.T.; Archinti, M.; Gygi, S.P.; Caelles, C.; Roig, J.; Luders, J. The gammaTuRC revisited: A comparative analysis of interphase and mitotic human gammaTuRC redefines the set of core components and identifies the novel subunit GCP8. Mol. Biol. Cell 2010, 21, 3963–3972. [Google Scholar] [CrossRef] [PubMed]
- Batzenschlager, M.; Masoud, K.; Janski, N.; Houlne, G.; Herzog, E.; Evrard, J.L.; Baumberger, N.; Erhardt, M.; Nomine, Y.; Kieffer, B.; et al. The GIP gamma-tubulin complex-associated proteins are involved in nuclear architecture in Arabidopsis thaliana. Front. Plant Sci. 2013, 4, 480. [Google Scholar] [CrossRef]
- Batzenschlager, M.; Lermontova, I.; Schubert, V.; Fuchs, J.; Berr, A.; Koini, M.A.; Houlne, G.; Herzog, E.; Rutten, T.; Alioua, A.; et al. Arabidopsis MZT1 homologs GIP1 and GIP2 are essential for centromere architecture. Proc. Natl. Acad. Sci. USA 2015, 112, 8656–8660. [Google Scholar] [CrossRef] [PubMed]
- Chaboute, M.E.; Berr, A. GIP Contributions to the Regulation of Centromere at the Interface Between the Nuclear Envelope and the Nucleoplasm. Front. Plant Sci. 2016, 7, 118. [Google Scholar] [CrossRef] [PubMed]
- Fal, K.; Asnacios, A.; Chaboute, M.E.; Hamant, O. Nuclear envelope: A new frontier in plant mechanosensing? Biophys. Rev. 2017, 9, 389–403. [Google Scholar] [CrossRef] [PubMed]
- Le, H.Q.; Ghatak, S.; Yeung, C.Y.; Tellkamp, F.; Gunschmann, C.; Dieterich, C.; Yeroslaviz, A.; Habermann, B.; Pombo, A.; Niessen, C.M.; et al. Mechanical regulation of transcription controls Polycomb-mediated gene silencing during lineage commitment. Nat. Cell Biol. 2016, 18, 864–875. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhou, Z. Lamin A/C, laminopathies and premature ageing. Histol. Histopathol. 2008, 23, 747–763. [Google Scholar] [CrossRef]
- Hamilton, E.S.; Jensen, G.S.; Maksaev, G.; Katims, A.; Sherp, A.M.; Haswell, E.S. Mechanosensitive channel MSL8 regulates osmotic forces during pollen hydration and germination. Science 2015, 350, 438–441. [Google Scholar] [CrossRef] [Green Version]
- Doskocilova, A.; Plihal, O.; Volc, J.; Chumova, J.; Kourova, H.; Halada, P.; Petrovska, B.; Binarova, P. A nodulin/glutamine synthetase-like fusion protein is implicated in the regulation of root morphogenesis and in signalling triggered by flagellin. Planta 2011, 234, 459–476. [Google Scholar] [CrossRef]
- Nemeth, K.; Salchert, K.; Putnoky, P.; Bhalerao, R.; Koncz-Kalman, Z.; Stankovic-Stangeland, B.; Bako, L.; Mathur, J.; Okresz, L.; Stabel, S.; et al. Pleiotropic control of glucose and hormone responses by PRL1, a nuclear WD protein, in Arabidopsis. Genes Dev. 1998, 12, 3059–3073. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Curran, A.; Chang, I.F.; Chang, C.L.; Garg, S.; Miguel, R.M.; Barron, Y.D.; Li, Y.; Romanowsky, S.; Cushman, J.C.; Gribskov, M.; et al. Calcium-dependent protein kinases from Arabidopsis show substrate specificity differences in an analysis of 103 substrates. Front. Plant Sci. 2011, 2, 36. [Google Scholar] [CrossRef] [PubMed]
- Benschop, J.J.; Mohammed, S.; O’Flaherty, M.; Heck, A.J.; Slijper, M.; Menke, F.L. Quantitative phosphoproteomics of early elicitor signaling in Arabidopsis. Mol. Cell Proteom. 2007, 6, 1198–1214. [Google Scholar] [CrossRef]
- Oda, Y. Cortical microtubule rearrangements and cell wall patterning. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Fukao, Y.; Iwamoto, M.; Haraguchi, T.; Hara-Nishimura, I. Identification and Characterization of Nuclear Pore Complex Components in Arabidopsis thaliana. Plant Cell 2010, 22, 4084–4097. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, A.; Palancade, B. Regulation of mRNA Trafficking by Nuclear Pore Complexes. Genes 2014, 5, 767–791. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-Y.; Lee, H.-S.; Wi, S.-J.; Park, K.Y.; Schmit, A.-C.; Pai, H.-S. Dual functions of Nicotiana benthamiana Rae1 in interphase and mitosis. Plant J. 2009, 59, 278–291. [Google Scholar] [CrossRef]
- Wagstaff, J.; Lowe, J. Prokaryotic cytoskeletons: Protein filaments organizing small cells. Nat. Rev. Microbiol. 2018, 16, 187–201. [Google Scholar] [CrossRef]
- Izore, T.; Kureisaite-Ciziene, D.; McLaughlin, S.H.; Lowe, J. Crenactin forms actin-like double helical filaments regulated by arcadin-2. Elife 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, Y.; Takagi, S. LITTLE NUCLEI 1 and 4 Regulate Nuclear Morphology in Arabidopsis thaliana. Plant Cell Physiol. 2013, 54, 622–633. [Google Scholar] [CrossRef]
- Wang, H.Y.; Dittmer, T.A.; Richards, E.J. Arabidopsis CROWDED NUCLEI (CRWN) proteins are required for nuclear size control and heterochromatin organization. BMC Plant Biol. 2013, 13. [Google Scholar] [CrossRef] [PubMed]
- Sundararajan, K.; Miguel, A.; Desmarais, S.M.; Meier, E.L.; Casey Huang, K.; Goley, E.D. The bacterial tubulin FtsZ requires its intrinsically disordered linker to direct robust cell wall construction. Nat. Commun. 2015, 6, 7281. [Google Scholar] [CrossRef] [PubMed]
- Magyar, Z.; Atanassova, A.; De Veylder, L.; Rombauts, S.; Inze, D. Characterization of two distinct DP-related genes from Arabidopsis thaliana. FEBS Lett. 2000, 486, 79–87. [Google Scholar] [CrossRef]
- Draberova, E.; D’Agostino, L.; Caracciolo, V.; Sladkova, V.; Sulimenko, T.; Sulimenko, V.; Sobol, M.; Maounis, N.F.; Tzelepis, E.; Mahera, E.; et al. Overexpression and Nucleolar Localization of gamma-Tubulin Small Complex Proteins GCP2 and GCP3 in Glioblastoma. J. Neuropathol. Exp. Neurol. 2015, 74, 723–742. [Google Scholar] [CrossRef] [PubMed]
- Horvath, B.M.; Kourova, H.; Nagy, S.; Nemeth, E.; Magyar, Z.; Papdi, C.; Ahmad, Z.; Sanchez-Perez, G.F.; Perilli, S.; Blilou, I.; et al. Arabidopsis RETINOBLASTOMA RELATED directly regulates DNA damage responses through functions beyond cell cycle control. EMBO J. 2017, 36, 1261–1278. [Google Scholar] [CrossRef] [PubMed]
- Novakova, M.; Draberova, E.; Schurmann, W.; Czihak, G.; Viklicky, V.; Dr-aber, P. gamma-Tubulin redistribution in taxol-treated mitotic cells probed by monoclonal antibodies. Cell Motil. Cytoskeleton 1996, 33, 38–51. [Google Scholar] [CrossRef]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chumová, J.; Kourová, H.; Trögelová, L.; Halada, P.; Binarová, P. Microtubular and Nuclear Functions of γ-Tubulin: Are They LINCed? Cells 2019, 8, 259. https://doi.org/10.3390/cells8030259
Chumová J, Kourová H, Trögelová L, Halada P, Binarová P. Microtubular and Nuclear Functions of γ-Tubulin: Are They LINCed? Cells. 2019; 8(3):259. https://doi.org/10.3390/cells8030259
Chicago/Turabian StyleChumová, Jana, Hana Kourová, Lucie Trögelová, Petr Halada, and Pavla Binarová. 2019. "Microtubular and Nuclear Functions of γ-Tubulin: Are They LINCed?" Cells 8, no. 3: 259. https://doi.org/10.3390/cells8030259
APA StyleChumová, J., Kourová, H., Trögelová, L., Halada, P., & Binarová, P. (2019). Microtubular and Nuclear Functions of γ-Tubulin: Are They LINCed? Cells, 8(3), 259. https://doi.org/10.3390/cells8030259