A Direct Effect of Sex Hormones on Epithelial Barrier Function in Inflammatory Bowel Disease Models



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Cell Lines
2.2. Organoid Culture
2.3. Reverse Transcriptase Polymerase Chain Reaction (rt-PCR)
2.4. Scratch Assay
2.5. MTT
2.6. TEER
2.7. Enzyme Linked Immunosorbent Assay (ELISA)
2.8. Western Blotting
2.9. Statistical Analysis
3. Results
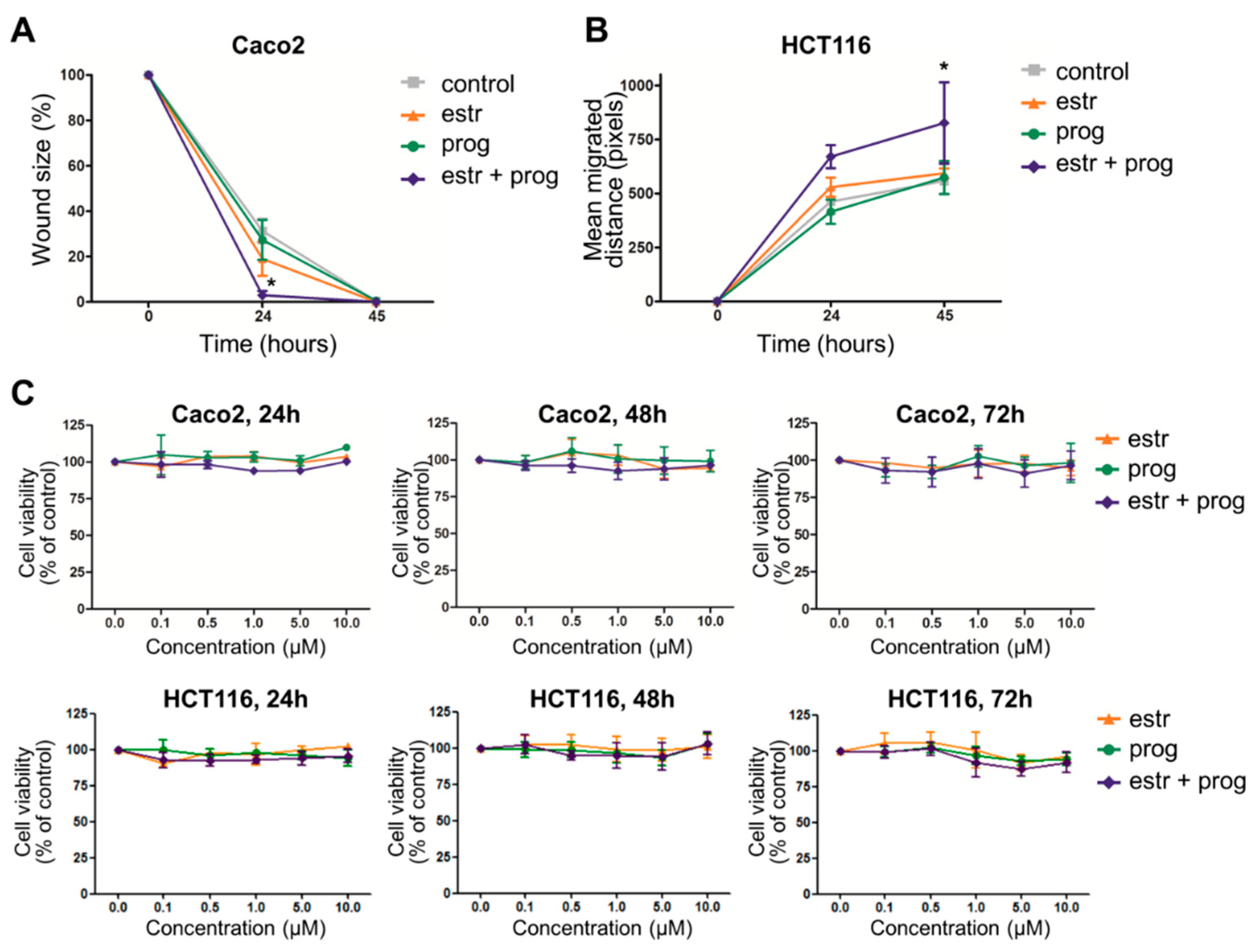
3.1. Estrogen and Progesterone Stimulate Wound Healing in Intestinal Epithelium
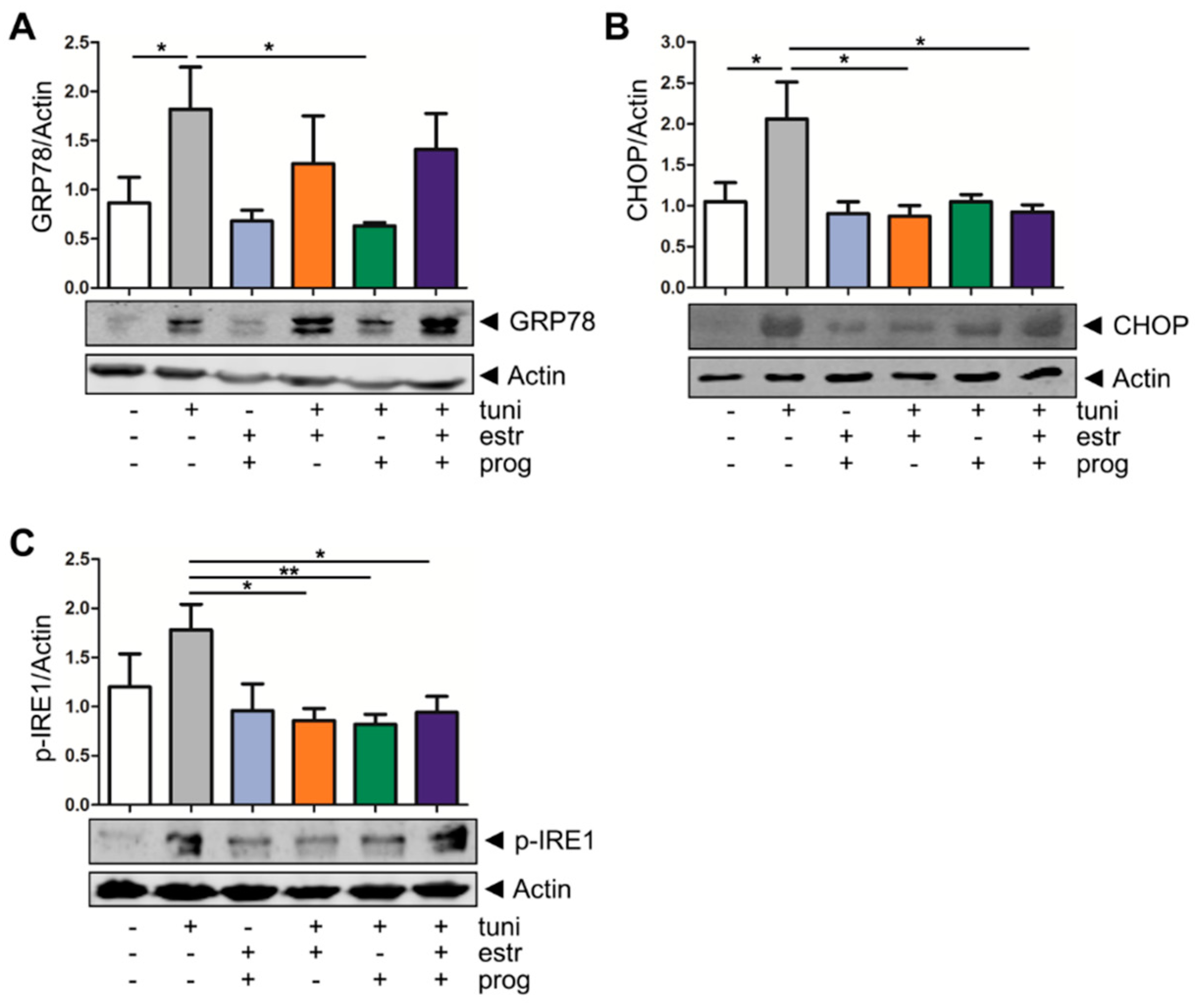
3.2. Estrogen and Progesterone Alleviate ER Stress in Intestinal Epithelium
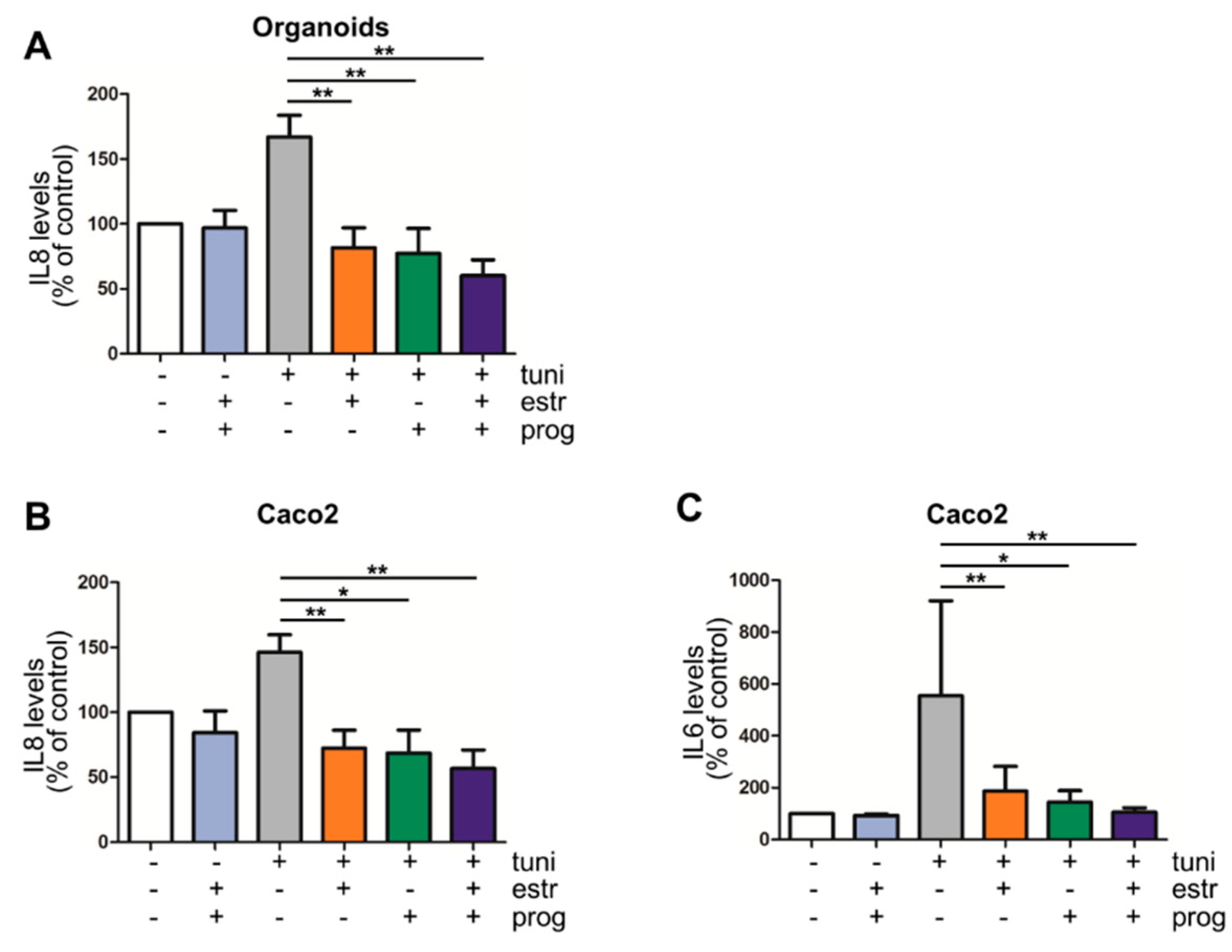
3.3. Pro-Inflammatory Cytokine Production by Intestinal Epithelial Cells Is Decreased in the Presence of Progesterone and Estrogen
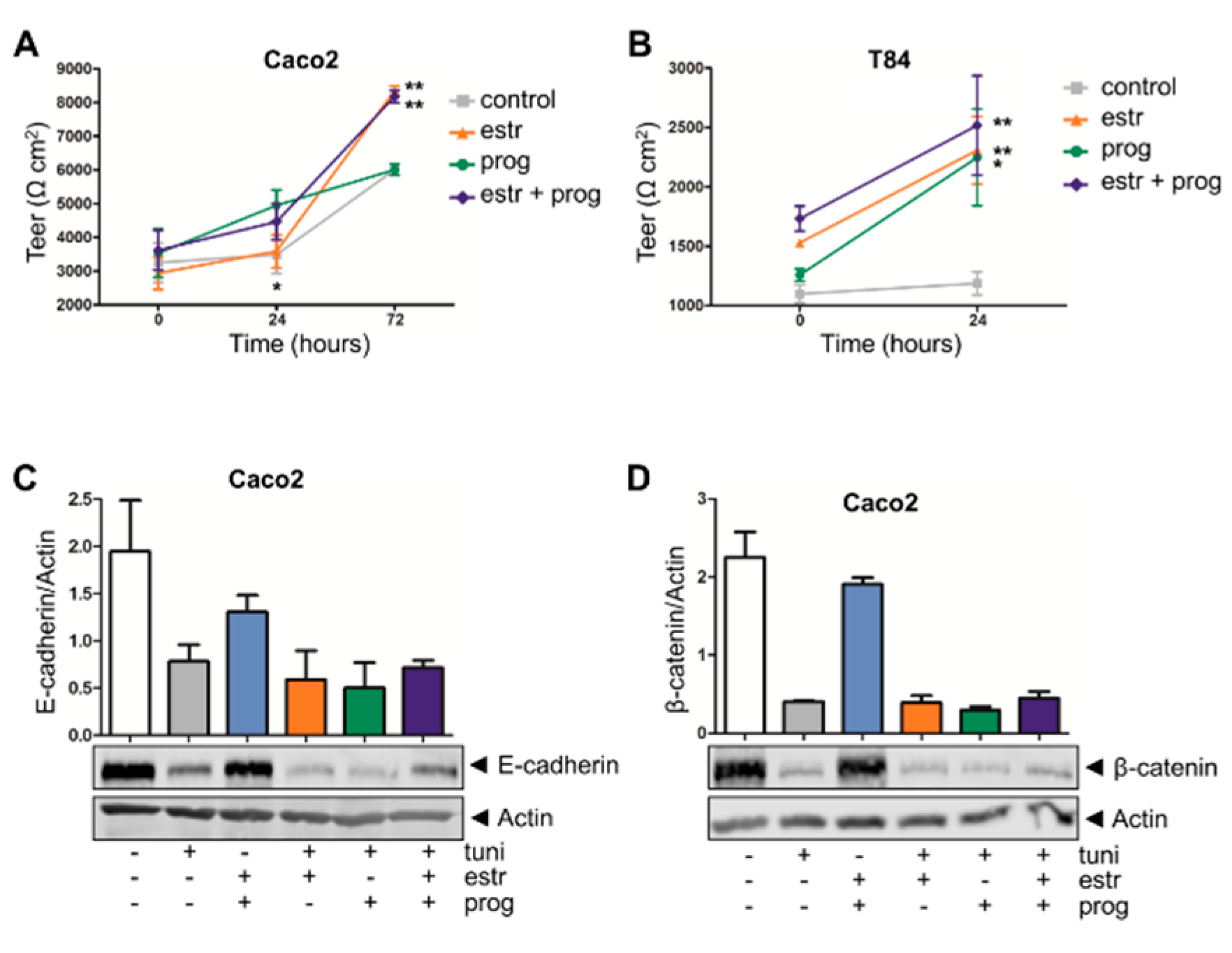
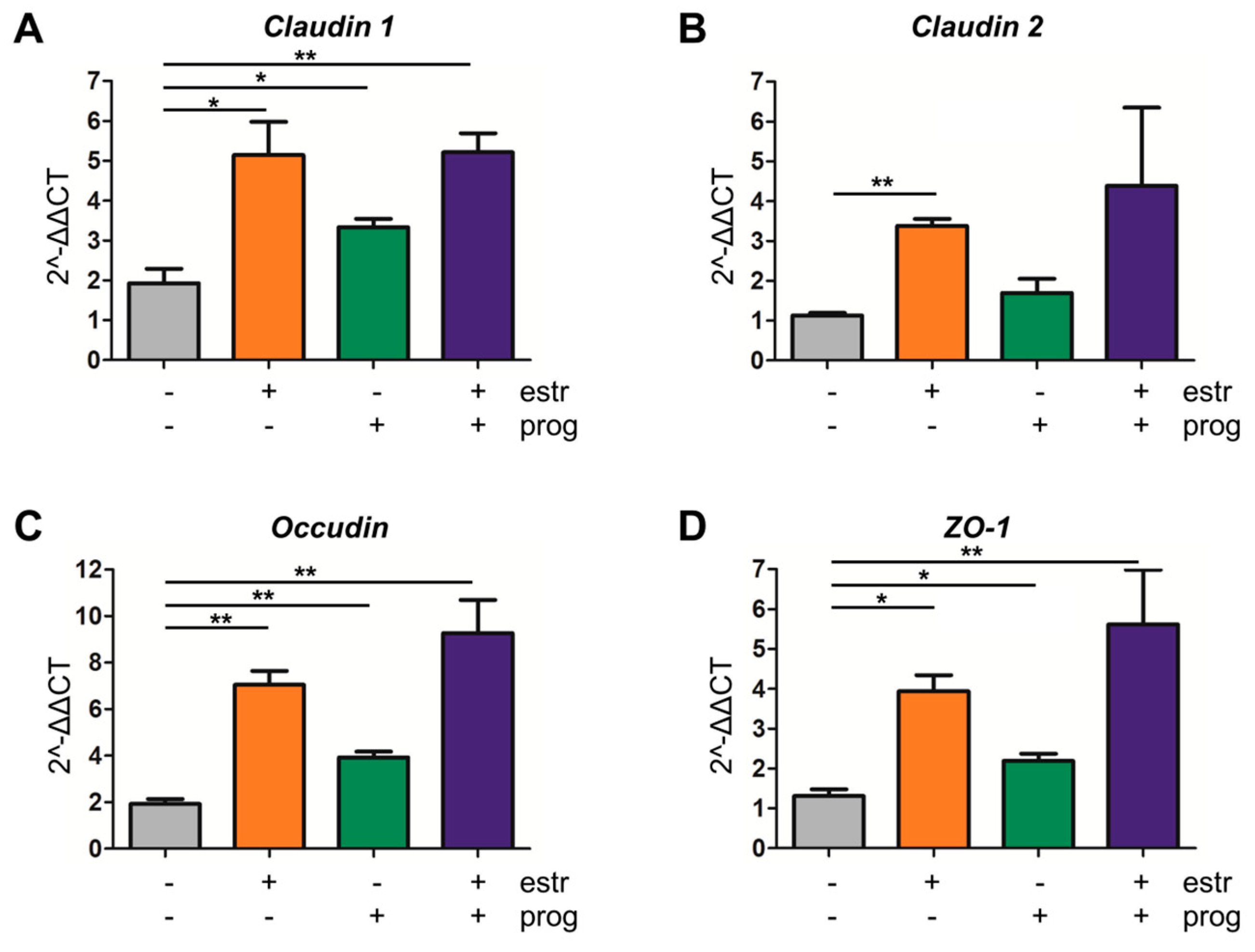
3.4. Improved Barrier Function Strength in the Presence of Estrogen and Progesterone
4. Discussion
Supplementary Materials
Author Contributions
Conflicts of Interest
References
- Kaplan, G.G. The global burden of IBD: From 2015 to 2025. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 720–727. [Google Scholar] [CrossRef]
- Johnston, R.D.; Logan, R.F.A. What is the peak age for onset of IBD? Inflamm. Bowel. Dis. 2008, 14, S4–S5. [Google Scholar] [CrossRef] [PubMed]
- Castiglione, F. Effect of pregnancy on the clinical course of a cohort of women with IBD. Ital. J. Gastroenterol. 1996, 28, 199–204. [Google Scholar] [PubMed]
- Riis, L.; Vind, I.; Politi, P.; Wolters, F.; Vermeire, S.; Tsianos, E.; Freitas, J.; Mouzas, I.; Ruiz Ochoa, V.; O’Morain, C.; et al. Does pregnancy change the disease course? A study in a European cohort of patients with inflammatory bowel disease. Am. J. Gastroenterol. 2006, 101, 1539–1545. [Google Scholar] [CrossRef]
- de Lima, A.; Zelinkova, Z.; van der Ent, C.; Steegers, E.A.P.; van der Woude, C.J. Tailored anti-TNF therapy during pregnancy in patients with IBD: Maternal and fetal safety. Gut 2016, 65, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Zelinkova, Z.; van der Ent, C.; Bruin, K.F.; van Baalen, O.; Vermeulen, H.G.; Smalbraak, H.J.T.; Ouwendijk, R.J.; Hoek, A.C.; van der Werf, S.D.; Kuipers, E.J.; et al. Effects of discontinuing anti-tumor necrosis factor therapy during pregnancy on the course of inflammatory bowel disease and neonatal exposure. Clin. Gastroenterol. Hepatol. 2013, 11, 318–321. [Google Scholar] [CrossRef]
- Pedersen, N.; Bortoli, A.; Duricova, D.; Dinca, R.; Panelli, M.R.; Gisbert, J.P.; Zoli, G.; López-Sanromán, A.; Castiglione, F.; Riegler, G.; et al. The course of inflammatory bowel disease during pregnancy and postpartum: A prospective European ECCO-EpiCom Study of 209 pregnant women. Aliment. Pharmacol. Ther. 2013, 38, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Lei, B.; Mace, B.; Dawson, H.N.; Warner, D.S.; Laskowitz, D.T.; James, M.L. Anti-Inflammatory Effects of Progesterone in Lipopolysaccharide-Stimulated BV-2 Microglia. PLoS ONE 2014, 9, e103969. [Google Scholar] [CrossRef] [PubMed]
- Straub, R.H. The Complex Role of Estrogens in Inflammation. Endocr. Rev. 2007, 28, 521–574. [Google Scholar] [CrossRef] [Green Version]
- Preciado-Martínez, E.; García-Ruíz, G.; Flores-Espinosa, P.; Bermejo-Martínez, L.; Espejel-Nuñez, A.; Estrada-Gutiérrez, G.; Razo-Aguilera, G.; Granados-Cepeda, M.; Helguera-Repetto, A.C.; Irles, C.; et al. Progesterone suppresses the lipopolysaccharide-induced pro-inflammatory response in primary mononuclear cells isolated from human placental blood. Immunol. Investig. 2018, 47, 181–195. [Google Scholar] [CrossRef]
- Khalili, H.; Higuchi, L.M.; Ananthakrishnan, A.N.; Manson, J.E.; Feskanich, D.; Richter, J.M.; Fuchs, C.S.; Chan, A.T. Hormone Therapy Increases Risk of Ulcerative Colitis but not Crohn’s Disease. Gastroenterology 2012, 143, 1199–1206. [Google Scholar] [CrossRef] [Green Version]
- Cornish, J.A.; Tan, E.; Simillis, C.; Clark, S.K.; Teare, J.; Tekkis, P.P. The Risk of Oral Contraceptives in the Etiology of Inflammatory Bowel Disease: A Meta-Analysis. Am. J. Gastroenterol. 2008, 103, 2394–2400. [Google Scholar] [CrossRef]
- Kane, S.V.; Reddy, D. Hormonal Replacement Therapy After Menopause Is Protective of Disease Activity in Women with Inflammatory Bowel Disease. Am. J. Gastroenterol. 2008, 103, 1193–1196. [Google Scholar] [CrossRef]
- Harnish, D.C.; Albert, L.M.; Leathurby, Y.; Eckert, A.M.; Ciarletta, A.; Kasaian, M.; Keith, J.C., Jr. Beneficial effects of estrogen treatment in the HLA-B27 transgenic rat model of inflammatory bowel disease. Am. J. Physiol. Liver Physiol. 2004, 286, G118–G125. [Google Scholar] [CrossRef]
- Verd, E.F.; Deng, Y.; Bercik, P.; Collins, S.M. Modulatory Effects of Estrogen in Two Murine Models of Experimental Colitis. Available online: http://www.physiology.org/doi/pdf/10.1152/ajpgi.00460.2001 (accessed on 2 February 2018).
- Karatepe, O.I.; Altiok, M.I.; Battal, M.; Kamali, G.I.; Kemik, A.V.; Aydin, T.V.; Karahan, S. The effect of progesterone in the prevention of the chemically induced experimental colitis in rats Efeito da progesterona na prevenção de colite experimental induzida quimicamente em ratos. Acta Cirúrgica Bras 2012, 27, 23–29. [Google Scholar] [CrossRef]
- Bouma, G.; Strober, W. The immunological and genetic basis of inflammatory bowel disease. Nat. Rev. Immunol. 2003, 3, 521–533. [Google Scholar] [CrossRef]
- Shen, L.; Su, L.; Turner, J.R. Mechanisms and functional implications of intestinal barrier defects. Dig. Dis. 2009, 27, 443–449. [Google Scholar] [CrossRef]
- Gassler, N.; Rohr, C.; Schneider, A.; Kartenbeck, J.; Bach, A.; Obermüller, N.; Otto, H.F.; Autschbach, F. Inflammatory bowel disease is associated with changes of enterocytic junctions. Am. J. Physiol. Liver Physiol. 2001, 281, G216–G228. [Google Scholar] [CrossRef]
- Konstantinopoulos, P.A.; Kominea, A.; Vandoros, G.; Sykiotis, G.P.; Andricopoulos, P.; Varakis, I.; Sotiropoulou-Bonikou, G.; Papavassiliou, A.G. Oestrogen receptor beta (ERbeta) is abundantly expressed in normal colonic mucosa, but declines in colon adenocarcinoma paralleling the tumour’s dedifferentiation. Eur. J. Cancer 2003, 39, 1251–1258. [Google Scholar] [CrossRef]
- Braniste, V.; Leveque, M.; Buisson-Brenac, C.; Bueno, L.; Fioramonti, J.; Houdeau, E. Oestradiol decreases colonic permeability through oestrogen receptor beta-mediated up-regulation of occludin and junctional adhesion molecule-A in epithelial cells. J. Physiol. 2009, 587, 3317–3328. [Google Scholar] [CrossRef]
- Homma, H.; Hoy, E.; Xu, D.-Z.; Lu, Q.; Feinman, R.; Deitch, E.A. The female intestine is more resistant than the male intestine to gut injury and inflammation when subjected to conditions associated with shock states. Am. J. Physiol. Liver Physiol. 2005, 288, G466–G472. [Google Scholar] [CrossRef] [PubMed]
- Dekkers, J.F.; Wiegerinck, C.L.; de Jonge, H.R.; Bronsveld, I.; Janssens, H.M.; de Winter-de Groot, K.M.; Brandsma, A.M.; de Jong, N.W.; Bijvelds, M.J.; Scholte, B.J.; et al. A functional CFTR assay using primary cystic fibrosis intestinal organoids. Nat. Med. 2013, 19, 939–945. [Google Scholar] [CrossRef]
- Yin, Y.; Wang, Y.; Dang, W.; Xu, L.; Su, J.; Zhou, X.; Wang, W.; Felczak, K.; van der Laan, L.J.; Pankiewicz, K.W.; et al. Mycophenolic Acid Potently Inhibits Rotavirus Infection with a High Barrier to Resistance Development. 2016. Available online: https://ac.els-cdn.com/S0166354216301097/1-s2.0-S0166354216301097-main.pdf?_tid=6f7dfc30-12f2-43c0-82f3-2c4048172105&acdnat=1527171066_52bdb7527063bd4ac0e47a84685cc304 (accessed on 24 May 2018).
- Lie, M.R.; van der Giessen, J.; Fuhler, G.M.; De Lima, A.; Peppelenbosch, M.P.; Van Der Ent, C.; van der Woude, C.J. Low dose Naltrexone for induction of remission in inflammatory bowel disease patients. J. Transl. Med. 2018, 16. [Google Scholar] [CrossRef]
- Queiroz, K.; Ruela-De-Sousa, R.; Fuhler, G.; Aberson, H.; Ferreira, C.; Peppelenbosch, M.; Spek, C.A. Hedgehog signaling maintains chemoresistance in myeloid leukemic cells. Oncogene 2010, 29, 6314–6322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somasundaram, R.; Nuij, V.J.; Van Der Woude, C.J.; Kuipers, E.J.; Peppelenbosch, M.P.; Fuhler, G.M. Peripheral Neutrophil Functions and Cell Signalling in Crohs Disease. PLoS ONE 2013, 8, e84521. [Google Scholar] [CrossRef] [PubMed]
- Peyrin-Biroulet, L.; Ferrante, M.; Magro, F.; Campbell, S.; Franchimont, D.; Fidder, H.; et al. Results from the 2nd Scientific Workshop of the ECCO (I): Impact of mucosal healing on the course of inflammatory bowel disease. J. Crohn’s Colitis 2011, 5, 477–483. [Google Scholar] [CrossRef] [Green Version]
- Arnauts, K.; Verstockt, B.; Vancamelbeke, M.; Vermeire, S.; Verfaillie, C.; Ferrante, M. Organoids Derived from Inflamed Intestinal Biopsies of Patients with Ulcerative Colitis Lose Their Inflammatory Phenotype during ex Vivo Culture. Preliminary Data Presented at the European Crohns and Colitis Organisation Meeting 2019. Available online: https://www.ecco-ibd.eu/publications/congress-abstract-s/abstracts-2019/item/op11-organoids-derived-from-inflamed-intestinal-biopsies-of-patients-with-ulcerative-colitis-lose-their-inflammatory-phenotype-during-italic-ex-vivo-italic-culture.html (accessed on 28 February 2019).
- Tschurtschenthaler, M.; Adolph, T.E.; Ashcroft, J.W.; Niederreiter, L.; Bharti, R.; Saveljeva, S.; Bhattacharyya, J.; Flak, M.B.; Shih, D.Q.; Fuhler, G.M.; et al. Defective ATG16L1-mediated removal of IRE1α drives Crohn’s disease-like ileitis. J. Exp. Med. 2017, 214, 401–422. [Google Scholar] [CrossRef] [PubMed]
- Powell, N.; Lo, J.W.; Biancheri, P.; Vossenkämper, A.; Pantazi, E.; Walker, A.W.; Stolarczyk, E.; Ammoscato, F.; Goldberg, R.; Scott, P.; et al. Interleukin 6 Increases Production of Cytokines by Colonic Innate Lymphoid Cells in Mice and Patients with Chronic Intestinal Inflammation. Gastroenterology 2015, 149, 456–467. [Google Scholar] [CrossRef]
- Tian, X.; Liu, Z.; Niu, B.; Zhang, J.; Tan, T.K.; Lee, S.R.; Zhao, Y.; Harris, D.C.; Zheng, G. E-cadherin/β-catenin complex and the epithelial barrier. J. Biomed. Biotechnol. 2011, 2011, 567305. [Google Scholar] [CrossRef]
- Ysrraelit, M.C.; Correale, J. Impact of sex hormones on immune function and multiple sclerosis development. Immunology 2019, 156, 9–22. [Google Scholar] [CrossRef]
- Barrett, J.H.; Brennan, P.; Fiddler, M.; Silman, A.J. Does rheumatoid arthritis remit during pregnancy and relapse postpartum? Results from a nationwide study in the United Kingdom performed prospectively from late pregnancy. Arthritis Rheum 1999, 42, 1219–1227. [Google Scholar] [CrossRef]
- de Man, Y.A.; Dolhain, R.J.; van de Geijn, F.E.; Willemsen, S.P.; Hazes, J.M. Disease activity of rheumatoid arthritis during pregnancy: Results from a nationwide prospective study. Arthritis Rheum 2008, 59, 1241–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soma-Pillay, P.; Nelson-Piercy, C.; Tolppanen, H.; Mebazaa, A.; Tolppanen, H.; Mebazaa, A. Physiological changes in pregnancy. Cardiovasc. J. Afr. 2016, 27, 89–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konstantinov, S.R.; van der Woude, C.J.; Peppelenbosch, M.P. Do pregnancy-related changes in the microbiome stimulate innate immunity? Trends Mol. Med. 2013, 19, 454–459. [Google Scholar] [CrossRef] [PubMed]
- Jostins, L.; Ripke, S.; Weersma, R.K.; Duerr, R.H.; McGovern, D.P.; Hui, K.Y.; Lee, J.C.; Schumm, L.P.; Sharma, Y.; Anderson, C.A.; et al. Host-microbe interactions have shaped the genetic architecture of inflammatory bowel disease. Nature 2012, 491, 119–124. [Google Scholar] [CrossRef] [Green Version]
- Ishihara, S.; Aziz, M.M.; Yuki, T.; Kazumori, H.; Kinoshita, Y. Inflammatory bowel disease: Review from the aspect of genetics. J. Gastroenterol. 2009, 44, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Deuring, J.J.; Fuhler, G.M.; Konstantinov, S.R.; Peppelenbosch, M.P.; Kuipers, E.J.; de Haar, C.; van der Woude, C.J. Genomic ATG16L1 risk allele-restricted Paneth cell ER stress in quiescent Crohn’s disease. Gut 2014, 63, 1081–1091. [Google Scholar] [CrossRef]
- Kim, S.; Joe, Y.; Kim, H.J.; Kim, Y.-S.; Jeong, S.O.; Pae, H.-O.; Ryter, S.W.; Surh, Y.J.; Chung, H.T. Endoplasmic reticulum stress-induced IRE1α activation mediates cross-talk of GSK-3β and XBP-1 to regulate inflammatory cytokine production. J. Immunol. 2015, 194, 4498–4506. [Google Scholar] [CrossRef]
- Xu, P.; Becker, H.; Elizalde, M.; Masclee, A.; Jonkers, D. Intestinal organoid culture model is a valuable system to study epithelial barrier function in IBD. Gut 2018, 67, 1905–1906. [Google Scholar] [CrossRef]
- Noben, M.; Verstockt, B.; de Bruyn, M.; Hendriks, N.; Van Assche, G.; Vermeire, S.; Verfaillie, C.; Ferrante, M. Epithelial organoid cultures from patients with ulcerative colitis and Crohn’s disease: A truly long-term model to study the molecular basis for inflammatory bowel disease? Gut 2017, 66, 2193–2195. [Google Scholar] [CrossRef]
- Grimm, M.C.; Elsbury, S.K.; Pavli, P.; Doe, W.F. Interleukin 8: Cells of origin in inflammatory bowel disease. Gut 1996, 38, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Mazzucchelli, L.; Hauser, C.; Zgraggen, K.; Wagner, H.; Hess, M.; Laissue, J.A.; Mueller, C. Expression of interleukin-8 gene in inflammatory bowel disease is related to the histological grade of active inflammation. Am. J. Pathol. 1994, 144, 997–1007. [Google Scholar] [PubMed]
- Frink, M.; Pape, H.-C.; Van Griensven, M.; Krettek, C.; Chaudry, I.H.; Hildebrand, F. Influence of Sex and Age on Mods and Cytokines After Multiple Injuries. Available online: https://insights.ovid.com/pubmed?pmid=17224789 (accessed on 28 January 2019).
- Khare, V.; Krnjic, A.; Frick, A.; Gmainer, C.; Asboth, M.; Jimenez, K.; Lang, M.; Baumgartner, M.; Evstatiev, R.; Gasche, C. Mesalamine and azathioprine modulate junctional complexes and restore epithelial barrier function in intestinal inflammation. Sci. Rep. 2019, 9, 2842. [Google Scholar] [CrossRef] [PubMed]
- Di Paolo, M.C.; Merrett, M.N.; Crotty, B.; Jewell, D.P. 5-Aminosalicylic acid inhibits the impaired epithelial barrier function induced by gamma interferon. Gut 1996, 38, 115. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
van der Giessen, J.; van der Woude, C.J.; Peppelenbosch, M.P.; Fuhler, G.M. A Direct Effect of Sex Hormones on Epithelial Barrier Function in Inflammatory Bowel Disease Models. Cells 2019, 8, 261. https://doi.org/10.3390/cells8030261
van der Giessen J, van der Woude CJ, Peppelenbosch MP, Fuhler GM. A Direct Effect of Sex Hormones on Epithelial Barrier Function in Inflammatory Bowel Disease Models. Cells. 2019; 8(3):261. https://doi.org/10.3390/cells8030261
Chicago/Turabian Stylevan der Giessen, Janine, C. Janneke van der Woude, Maikel P. Peppelenbosch, and Gwenny M. Fuhler. 2019. "A Direct Effect of Sex Hormones on Epithelial Barrier Function in Inflammatory Bowel Disease Models" Cells 8, no. 3: 261. https://doi.org/10.3390/cells8030261
APA Stylevan der Giessen, J., van der Woude, C. J., Peppelenbosch, M. P., & Fuhler, G. M. (2019). A Direct Effect of Sex Hormones on Epithelial Barrier Function in Inflammatory Bowel Disease Models. Cells, 8(3), 261. https://doi.org/10.3390/cells8030261