Skeletal Muscle Response to Deflazacort, Dexamethasone and Methylprednisolone

 , , , , ,
, , , , ,
Abstract
:
1. Introduction
2. Material and Methods
2.1. Animals and Drug Treatment
2.2. Glucocorticoid Administration
2.3. Cross-Sectional Areas Evaluation
2.4. SDS-PAGE (Western Blotting)
2.5. Quantitative PCR
2.6. Statistical Analysis
3. Results
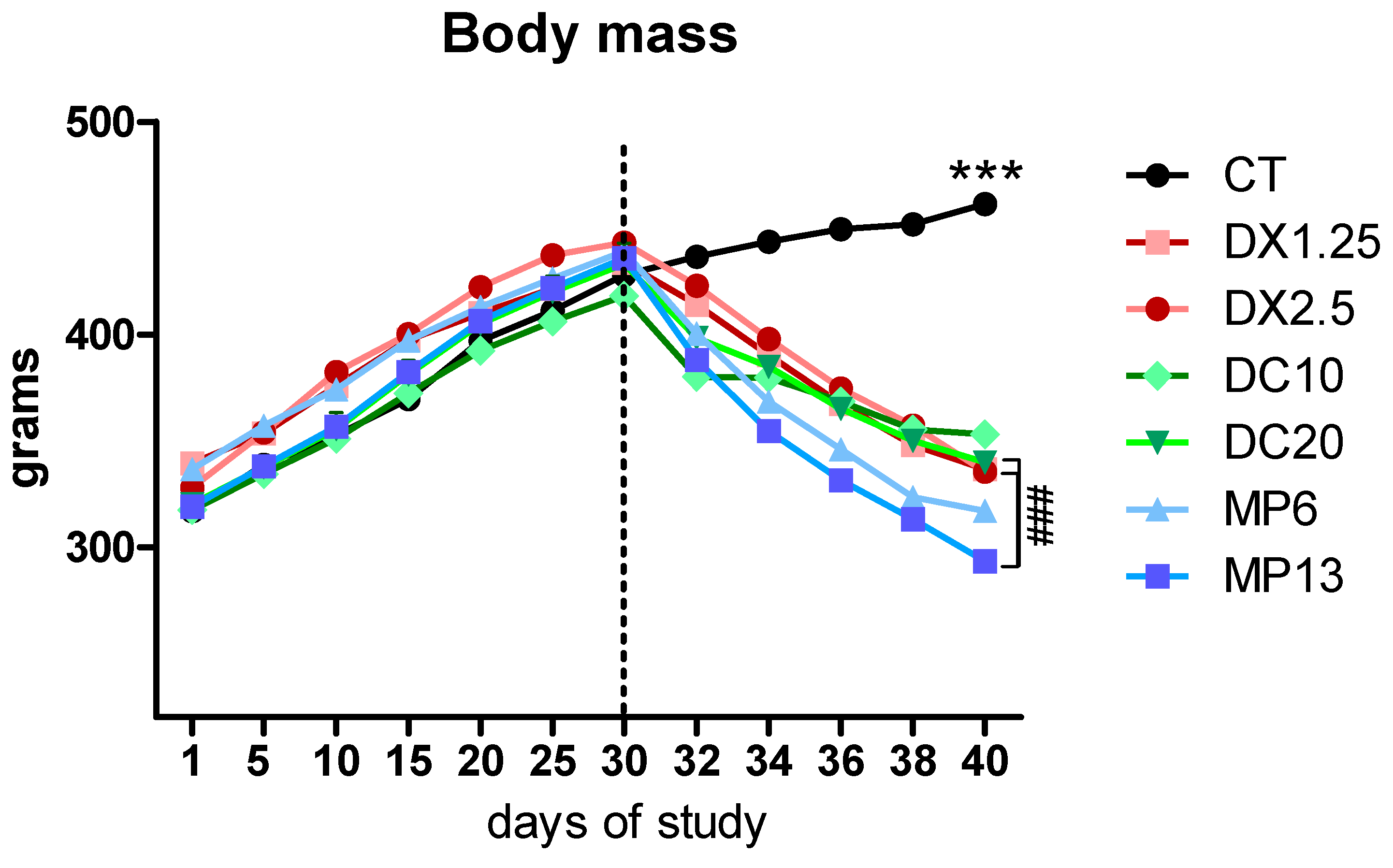
3.1. Body and Tissue Weights Resulting from Different Glucocorticoids Administration
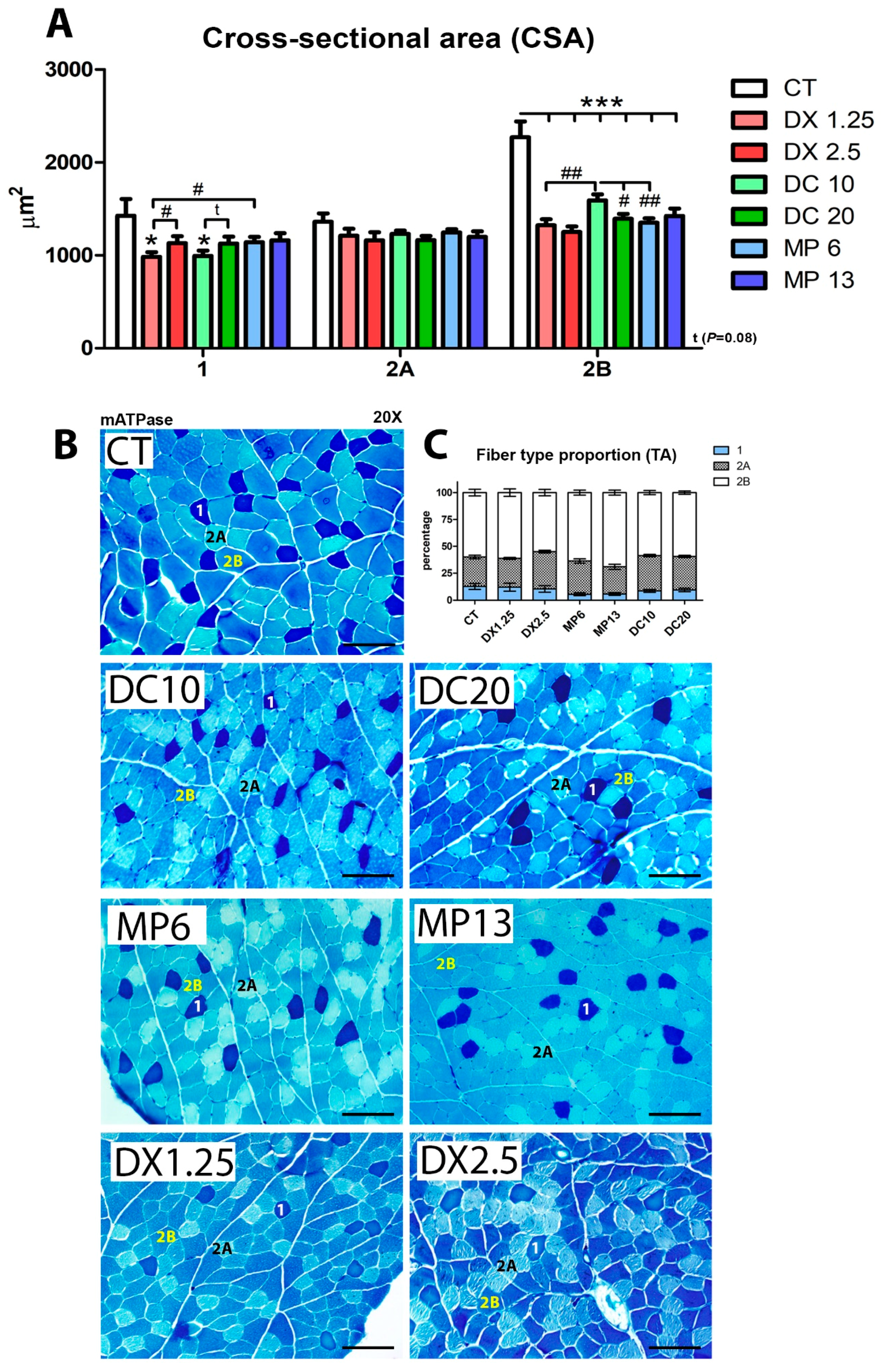
3.2. Cross-Sectional Areas (CSA) and Fiber Type Proportion Resulting from Different Glucocorticoids Administration
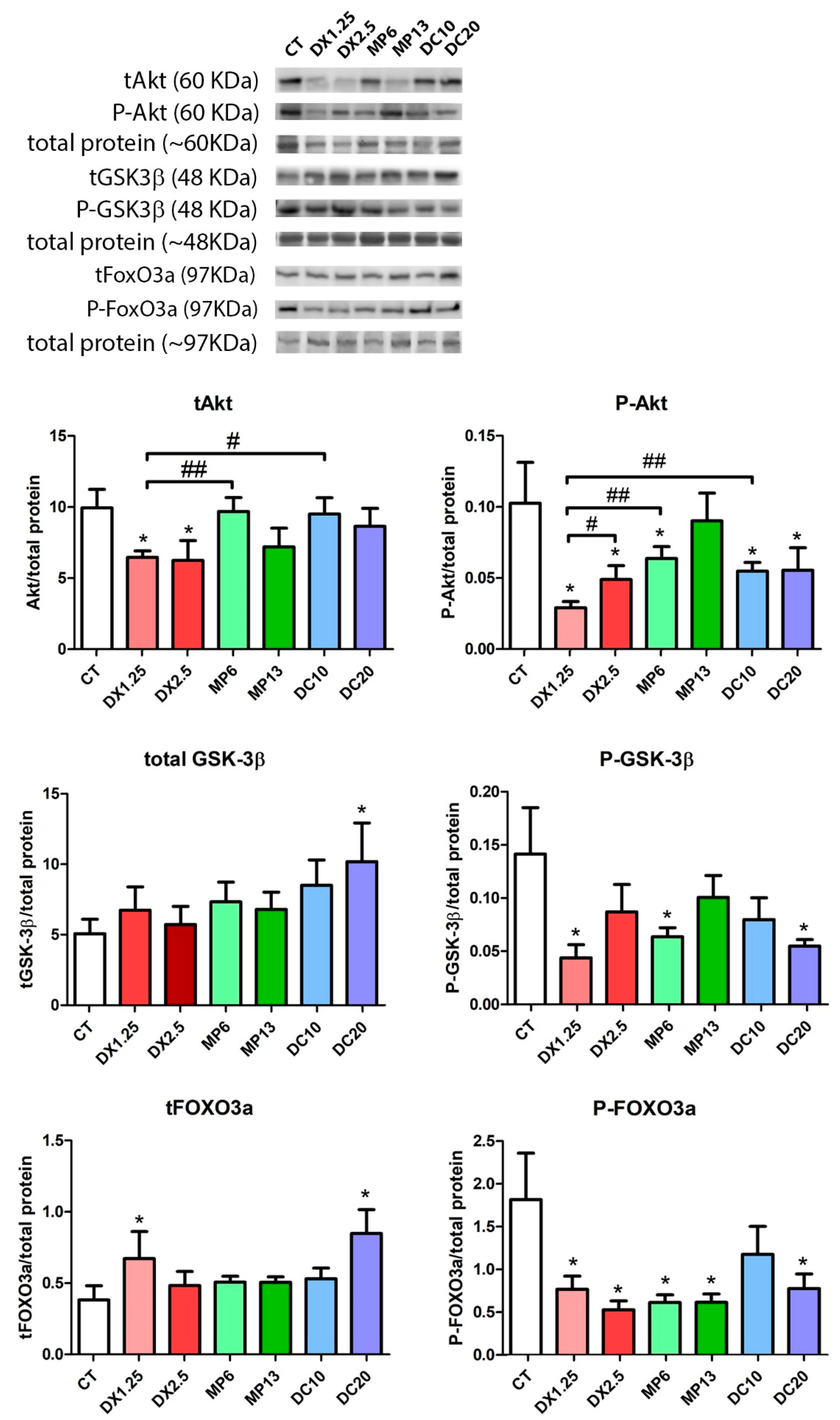
3.3. IGF-1 and Atrogenes Expression Resulting from Different Glucocorticoids Administration
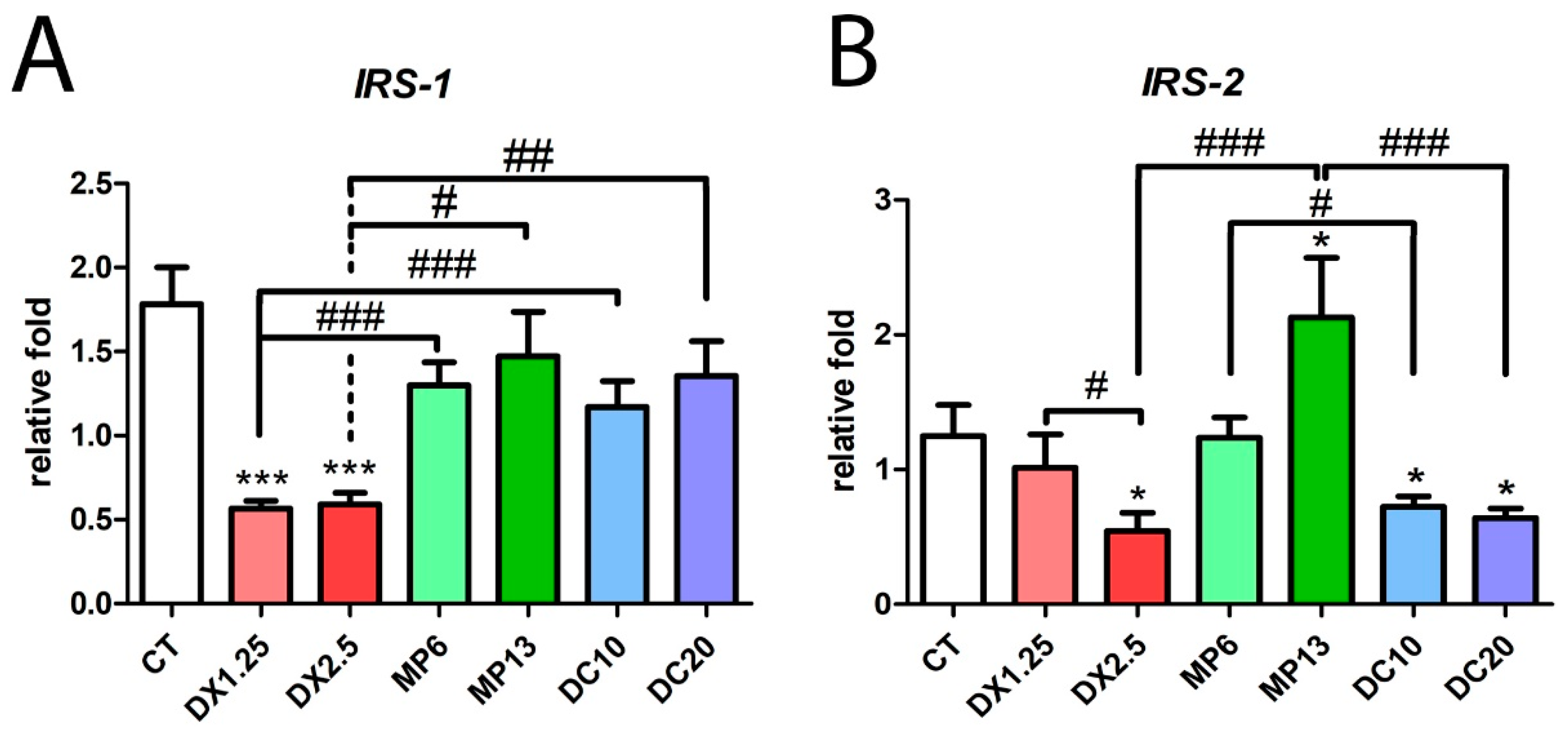
3.4. Insulin Receptor Substrate 1/2 Gene Expression Resulting from Different Glucocorticoids Administration
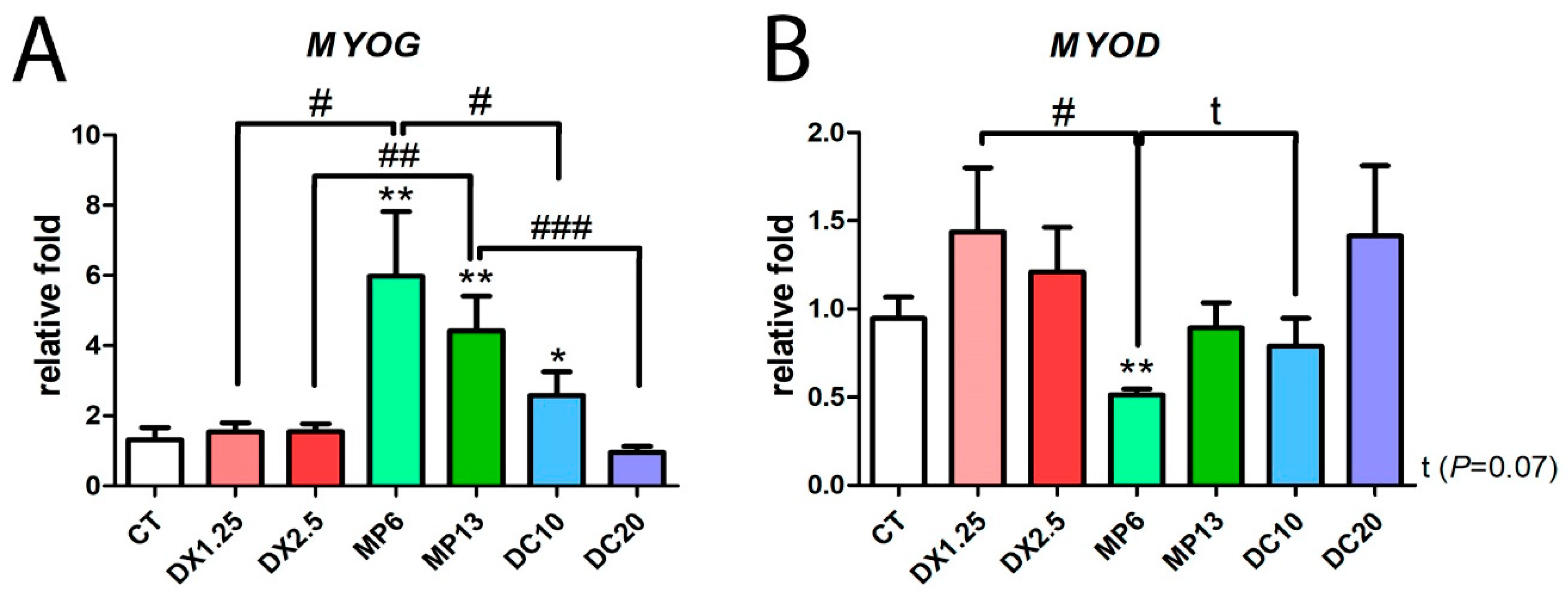
3.5. Methylprednisolone Administration Leads to An Inversion of Myogenic mRNA Expression
3.6. MEK/ERK Pathway Changes Resulting from Different Glucocorticoids Administration
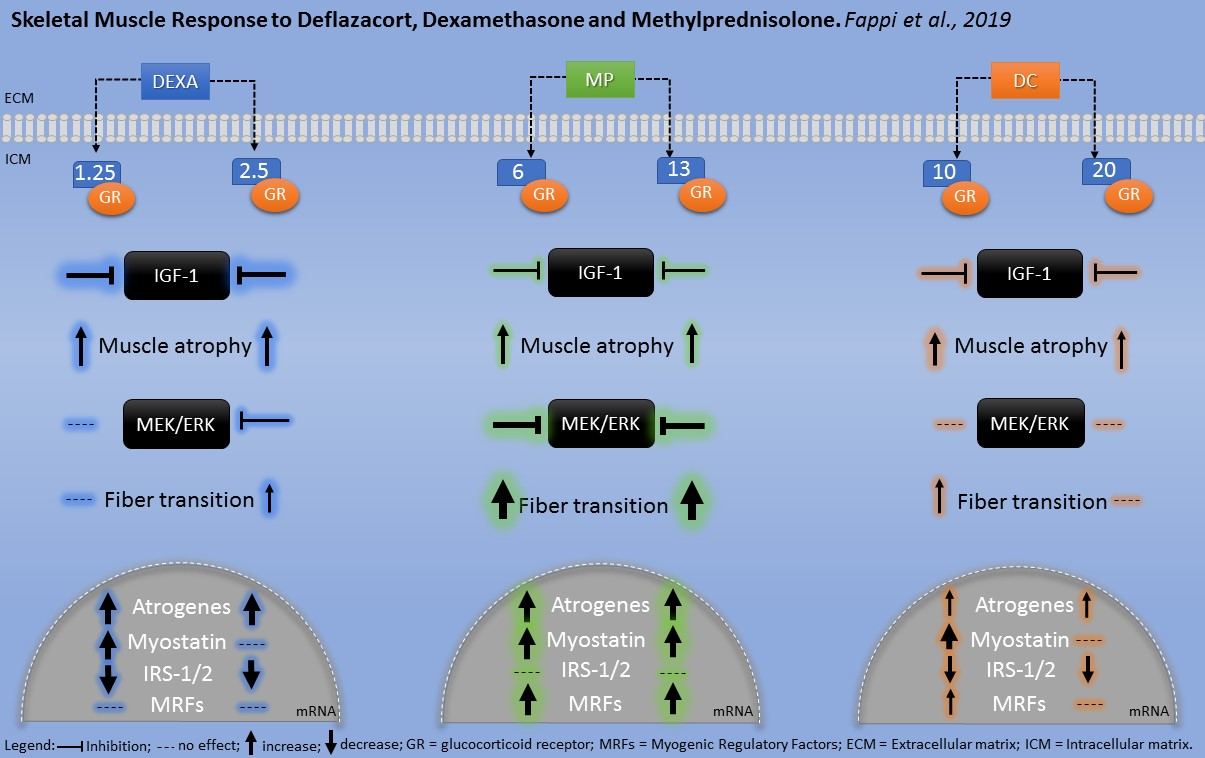
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Barnes, P.J. Glucocorticoids. Chem. Immunol. Allergy. 2014, 100, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Davies, E.; MacKenzie, S.M. Extra-adrenal production of corticosteroids. Clin. Exp. Pharmacol. Physiol. 2003, 30, 437–445. [Google Scholar] [CrossRef]
- Taves, M.D.; Gomez-Sanchez, C.E.; Soma, K.K. Extra-adrenal glucocorticoids and mineralocorticoids: Evidence for local synthesis, regulation, and function. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E11–E24. [Google Scholar] [CrossRef] [PubMed]
- Chrousos, G.P.; Charmandari, E.; Kino, T. Glucocorticoid action networks—An introduction to systems biology. J. Clin. Endocrinol. Metab. 2004, 89, 563–564. [Google Scholar] [CrossRef] [PubMed]
- Amon, A.; Brune, K.; Pahl, A. The anti-inflammatory action of glucocorticoids and PDE4-inhibitors is mediated in part by modulation of MAPK signaling pathways. N-S Arch. Pharmacol. 2005, 371, R72. [Google Scholar]
- Nicolaides, N.C.; Charmandari, E.; Chrousos, G.P.; Kino, T. Recent advances in the molecular mechanisms determining tissue sensitivity to glucocorticoids: Novel mutations, circadian rhythm and ligand-induced repression of the human glucocorticoid receptor. BMC Endocr. Disord. 2014, 14. [Google Scholar] [CrossRef]
- Birnkrant, D.J.; Bushby, K.; Bann, C.M.; Apkon, S.D.; Blackwell, A.; Brumbaugh, D.; Case, L.E.; Clemens, P.R.; Hadjiyannakis, S.; Pandya, S.; et al. Diagnosis and management of Duchenne muscular dystrophy, part 1: Diagnosis, and neuromuscular, rehabilitation, endocrine, and gastrointestinal and nutritional management. Lancet Neurol. 2018, 17, 251–267. [Google Scholar] [CrossRef]
- Moxley, R.T., 3rd; Pandya, S.; Ciafaloni, E.; Fox, D.J.; Campbell, K. Change in natural history of Duchenne muscular dystrophy with long-term corticosteroid treatment: Implications for management. J. Child. Neurol. 2010, 25, 1116–1129. [Google Scholar] [CrossRef]
- Houde, S.; Filiatrault, M.; Fournier, A.; Dube, J.; D’Arcy, S.; Berube, D.; Brousseau, Y.; Lapierre, G.; Vanasse, M. Deflazacort use in Duchenne muscular dystrophy: An 8-year follow-up. Pediatr. Neurol. 2008, 38, 200–206. [Google Scholar] [CrossRef]
- Schacke, H.; Docke, W.D.; Asadullah, K. Mechanisms involved in the side effects of glucocorticoids. Pharmacol. Therapeut. 2002, 96, 23–43. [Google Scholar] [CrossRef]
- Schakman, O.; Kalista, S.; Barbe, C.; Loumaye, A.; Thissen, J.P. Glucocorticoid-induced skeletal muscle atrophy. Int. J. Biochem. Cell Biol. 2013, 45, 2163–2172. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Gupta, Y. Glucocorticoid-induced myopathy: Pathophysiology, diagnosis, and treatment. Indian J. Endocrinol. Metab. 2013, 17, 913–916. [Google Scholar] [CrossRef] [PubMed]
- Gordon, B.S.; Kelleher, A.R.; Kimball, S.R. Regulation of muscle protein synthesis and the effects of catabolic states. Int. J. Biochem. Cell Biol. 2013, 45, 2147–2157. [Google Scholar] [CrossRef]
- Bentzinger, C.F.; Romanino, K.; Cloetta, D.; Lin, S.; Mascarenhas, J.B.; Oliveri, F.; Xia, J.; Casanova, E.; Costa, C.F.; Brink, M.; et al. Skeletal muscle-specific ablation of raptor, but not of rictor, causes metabolic changes and results in muscle dystrophy. Cell Metab. 2008, 8, 411–424. [Google Scholar] [CrossRef] [PubMed]
- Brugarolas, J.; Lei, K.; Hurley, R.L.; Manning, B.D.; Reiling, J.H.; Hafen, E.; Witters, L.A.; Ellisen, L.W.; Kaelin, W.G., Jr. Regulation of mTOR function in response to hypoxia by REDD1 and the TSC1/TSC2 tumor suppressor complex. Genes Dev. 2004, 18, 2893–2904. [Google Scholar] [CrossRef] [PubMed]
- Sandri, M.; Lin, J.; Handschin, C.; Yang, W.; Arany, Z.P.; Lecker, S.H.; Goldberg, A.L.; Spiegelman, B.M. PGC-1alpha protects skeletal muscle from atrophy by suppressing FoxO3 action and atrophy-specific gene transcription. Proc. Natl. Acad. Sci. USA 2006, 103, 16260–16265. [Google Scholar] [CrossRef]
- Shimizu, N.; Yoshikawa, N.; Ito, N.; Maruyama, T.; Suzuki, Y.; Takeda, S.; Nakae, J.; Tagata, Y.; Nishitani, S.; Takehana, K.; et al. Crosstalk between glucocorticoid receptor and nutritional sensor mTOR in skeletal muscle. Cell Metab. 2011, 13, 170–182. [Google Scholar] [CrossRef]
- Braun, T.P.; Marks, D.L. The regulation of muscle mass by endogenous glucocorticoids. Front. Physiol. 2015, 6, 12. [Google Scholar] [CrossRef]
- Bodine, S.C.; Latres, E.; Baumhueter, S.; Lai, V.K.; Nunez, L.; Clarke, B.A.; Poueymirou, W.T.; Panaro, F.J.; Na, E.; Dharmarajan, K.; et al. Identification of ubiquitin ligases required for skeletal muscle atrophy. Science 2001, 294, 1704–1708. [Google Scholar] [CrossRef]
- Sacheck, J.M.; Hyatt, J.P.; Raffaello, A.; Jagoe, R.T.; Roy, R.R.; Edgerton, V.R.; Lecker, S.H.; Goldberg, A.L. Rapid disuse and denervation atrophy involve transcriptional changes similar to those of muscle wasting during systemic diseases. FASEB J. 2007, 21, 140–155. [Google Scholar] [CrossRef]
- Gennari, C.; Imbimbo, B.; Montagnani, M.; Bernini, M.; Nardi, P.; Avioli, L.V. Effects of Prednisone and Deflazacort on Mineral Metabolism and Parathyroid-Hormone Activity in Humans. Calcified Tissue Int. 1984, 36, 245–252. [Google Scholar] [CrossRef]
- Gennari, C.; Imbimbo, B. Effects of prednisone and deflazacort on vertebral bone mass. Calcified Tissue Int. 1985, 37, 592–593. [Google Scholar] [CrossRef]
- Pagano, G.; Bruno, A.; Cavallo-Perin, P.; Cesco, L.; Imbimbo, B. Glucose intolerance after short-term administration of corticosteroids in healthy subjects. Prednisone, deflazacort, and betamethasone. Arch. Intern. Med. 1989, 149, 1098–1101. [Google Scholar] [CrossRef]
- Reitter, B. Deflazacort vs. prednisone in Duchenne muscular dystrophy: Trends of an ongoing study. Brain Dev. 1995, 17, 39–43. [Google Scholar] [CrossRef]
- Angelini, C. The role of corticosteroids in muscular dystrophy: A critical appraisal. Muscle Nerve 2007, 36, 424–435. [Google Scholar] [CrossRef]
- Angelini, C.; Pegoraro, E.; Turella, E.; Intino, M.T.; Pini, A.; Costa, C. Deflazacort in Duchenne dystrophy: Study of long-term effect. Muscle Nerve 1994, 17, 386–391. [Google Scholar] [CrossRef]
- Dekhuijzen, P.N.; Gayan-Ramirez, G.; Bisschop, A.; de Bock, V.; Dom, R.; Bouillon, R.; Decramer, M. Rat diaphragm contractility and histopathology are affected differently by low dose treatment with methylprednisolone and deflazacort. Eur. Respir. J. 1995, 8, 824–830. [Google Scholar]
- Anderson, J.E.; McIntosh, L.M.; Poettcker, R. Deflazacort but not prednisone improves both muscle repair and fiber growth in diaphragm and limb muscle in vivo in the mdx dystrophic mouse. Muscle Nerve 1996, 19, 1576–1585. [Google Scholar] [CrossRef]
- Kelly, F.J.; McGrath, J.A.; Goldspink, D.F.; Cullen, M.J. A morphological/biochemical study on the actions of corticosteroids on rat skeletal muscle. Muscle Nerve 1986, 9, 1–10. [Google Scholar] [CrossRef]
- Kelly, F.J.; Goldspink, D.F. The differing responses of four muscle types to dexamethasone treatment in the rat. Biochem. J. 1982, 208, 147–151. [Google Scholar] [CrossRef]
- Meikle, A.W.; Tyler, F.H. Potency and duration of action of glucocorticoids. Effects of hydrocortisone, prednisone and dexamethasone on human pituitary-adrenal function. Am. J. Med. 1977, 63, 200–207. [Google Scholar] [CrossRef]
- Nayak, S.; Acharjya, B. Deflazacort versus other glucocorticoids: A comparison. Indian J. Dermatol. 2008, 53, 167–170. [Google Scholar] [CrossRef]
- Ogilvie, R.W.; Feeback, D.L. A metachromatic dye-ATPase method for the simultaneous identification of skeletal muscle fiber types I, IIA, IIB and IIC. Stain Technol. 1990, 65, 231–241. [Google Scholar] [CrossRef]
- Waters, S.B.; Pessin, J.E. Insulin receptor substrate 1 and 2 (IRS1 and IRS2): What a tangled web we weave. Trends Cell Biol. 1996, 6, 1–4. [Google Scholar] [CrossRef]
- Higginson, J.; Wackerhage, H.; Woods, N.; Schjerling, P.; Ratkevicius, A.; Grunnet, N.; Quistorff, B. Blockades of mitogen-activated protein kinase and calcineurin both change fibre-type markers in skeletal muscle culture. Pflugers Arch. 2002, 445, 437–443. [Google Scholar] [CrossRef]
- Nava, S.; Gayan-Ramirez, G.; Rollier, H.; Bisschop, A.; Dom, R.; de Bock, V.; Decramer, M. Effects of acute steroid administration on ventilatory and peripheral muscles in rats. Am. J. Resp. Crit.Care Med. 1996, 153, 1888–1896. [Google Scholar] [CrossRef]
- Yoshimura, M.; Nakamura, A.; Kobayashi, M.; Takeda, S. Deflazacort induced severe skeletal muscle wasting and inguinal herniation in normal Beagle dogs. Neuromuscular Disord. 2007, 17, 775. [Google Scholar] [CrossRef]
- Maes, K.; Testelmans, D.; Cadot, P.; Deruisseau, K.; Powers, S.K.; Decramer, M.; Gayan-Ramirez, G. Effects of acute administration of corticosteroids during mechanical ventilation on rat diaphragm. Am. J. Resp. Crit. Care Med. 2008, 178, 1219–1226. [Google Scholar] [CrossRef]
- Maes, K.; Agten, A.; Smuder, A.; Powers, S.K.; Decramer, M.; Gayan-Ramirez, G. Corticosteroid effects on ventilator-induced diaphragm dysfunction in anesthetized rats depend on the dose administered. Respir. Res. 2010, 11, 178. [Google Scholar] [CrossRef]
- Pey, P.; Daminet, S.; Smets, P.M.; Duchateau, L.; Travetti, O.; Saunders, J.H. Effect of glucocorticoid administration on adrenal gland size and sonographic appearance in beagle dogs. Vet. Radiol. Ultrasound 2012, 53, 204–209. [Google Scholar] [CrossRef]
- Karatsoreos, I.N.; Bhagat, S.M.; Bowles, N.P.; Weil, Z.M.; Pfaff, D.W.; McEwen, B.S. Endocrine and physiological changes in response to chronic corticosterone: A potential model of the metabolic syndrome in mouse. Endocrinology 2010, 151, 2117–2127. [Google Scholar] [CrossRef]
- Jefferson, L.S.; Fabian, J.R.; Kimball, S.R. Glycogen synthase kinase-3 is the predominant insulin-regulated eukaryotic initiation factor 2B kinase in skeletal muscle. Int. J. Biochem. Cell Biol. 1999, 31, 191–200. [Google Scholar] [CrossRef]
- Bodine, S.C.; Stitt, T.N.; Gonzalez, M.; Kline, W.O.; Stover, G.L.; Bauerlein, R.; Zlotchenko, E.; Scrimgeour, A.; Lawrence, J.C.; Glass, D.J.; et al. Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo. Nat. Cell Biol. 2001, 3, 1014–1019. [Google Scholar] [CrossRef]
- Leger, B.; Cartoni, R.; Praz, M.; Lamon, S.; Deriaz, O.; Crettenand, A.; Gobelet, C.; Rohmer, P.; Konzelmann, M.; Luthi, F.; et al. Akt signalling through GSK-3beta, mTOR and Foxo1 is involved in human skeletal muscle hypertrophy and atrophy. J. Physiol 2006, 576, 923–933. [Google Scholar] [CrossRef]
- Gundersen, K. Excitation-transcription coupling in skeletal muscle: The molecular pathways of exercise. Biol. Rev. Camb. Philos. Soc. 2011, 86, 564–600. [Google Scholar] [CrossRef]
- Ciaraldi, T.P.; Carter, L.; Mudaliar, S.; Henry, R.R. GSK-3beta and control of glucose metabolism and insulin action in human skeletal muscle. Mol. Cell. Endocrinol. 2010, 315, 153–158. [Google Scholar] [CrossRef]
- Schiaffino, S.; Dyar, K.A.; Ciciliot, S.; Blaauw, B.; Sandri, M. Mechanisms regulating skeletal muscle growth and atrophy. FEBS J. 2013, 280, 4294–4314. [Google Scholar] [CrossRef]
- Kuo, T.; Lew, M.J.; Mayba, O.; Harris, C.A.; Speed, T.P.; Wang, J.C. Genome-wide analysis of glucocorticoid receptor-binding sites in myotubes identifies gene networks modulating insulin signaling. Proc. Natl. Acad. Sci. USA 2012, 109, 11160–11165. [Google Scholar] [CrossRef]
- Metz, H.E.; Houghton, A.M. Insulin receptor substrate regulation of phosphoinositide 3-kinase. Clin. Cancer Res. 2011, 17, 206–211. [Google Scholar] [CrossRef]
- Katiyar, S.; Liu, E.; Knutzen, C.A.; Lang, E.S.; Lombardo, C.R.; Sankar, S.; Toth, J.I.; Petroski, M.D.; Ronai, Z.; Chiang, G.G. REDD1, an inhibitor of mTOR signalling, is regulated by the CUL4A-DDB1 ubiquitin ligase. EMBO Rep. 2009, 10, 866–872. [Google Scholar] [CrossRef]
- Ellisen, L.W.; Ramsayer, K.D.; Johannessen, C.M.; Yang, A.; Beppu, H.; Minda, K.; Oliner, J.D.; McKeon, F.; Haber, D.A. REDD1, a developmentally regulated transcriptional target of p63 and p53, links p63 to regulation of reactive oxygen species. Mol. Cell 2002, 10, 995–1005. [Google Scholar] [CrossRef]
- Lin, L.; Stringfield, T.M.; Shi, X.; Chen, Y. Arsenite induces a cell stress-response gene, RTP801, through reactive oxygen species and transcription factors Elk-1 and CCAAT/enhancer-binding protein. Biochem. J. 2005, 392, 93–102. [Google Scholar] [CrossRef]
- McClung, J.M.; Judge, A.R.; Powers, S.K.; Yan, Z. p38 MAPK links oxidative stress to autophagy-related gene expression in cachectic muscle wasting. Am. J. Physiol. Cell Physiol. 2010, 298, C542–C549. [Google Scholar] [CrossRef]
- Nishida, H.; Ikegami, A.; Kaneko, C.; Kakuma, H.; Nishi, H.; Tanaka, N.; Aoyama, M.; Usami, M.; Okimura, Y. Dexamethasone and BCAA Failed to Modulate Muscle Mass and mTOR Signaling in GH-Deficient Rats. PLoS ONE 2015, 10, e0128805. [Google Scholar] [CrossRef]
- Britto, F.A.; Begue, G.; Rossano, B.; Docquier, A.; Vernus, B.; Sar, C.; Ferry, A.; Bonnieu, A.; Ollendorff, V.; Favier, F.B. REDD1 deletion prevents dexamethasone-induced skeletal muscle atrophy. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E983–E993. [Google Scholar] [CrossRef]
- Wang, H.; Kubica, N.; Ellisen, L.W.; Jefferson, L.S.; Kimball, S.R. Dexamethasone represses signaling through the mammalian target of rapamycin in muscle cells by enhancing expression of REDD1. J. Biol. Chem. 2006, 281, 39128–39134. [Google Scholar] [CrossRef]
- Tsuchida, W.; Iwata, M.; Akimoto, T.; Matsuo, S.; Asai, Y.; Suzuki, S. Heat Stress Modulates Both Anabolic and Catabolic Signaling Pathways Preventing Dexamethasone-Induced Muscle Atrophy In Vitro. J. Cell. Physiol. 2017, 232, 650–664. [Google Scholar] [CrossRef]
- Kelleher, A.R.; Pereira, S.L.; Jefferson, L.S.; Kimball, S.R. REDD2 expression in rat skeletal muscle correlates with nutrient-induced activation of mTORC1: Responses to aging, immobilization, and remobilization. Am. J. Physiol. Endocrinol. Metab. 2015, 308, E122–E129. [Google Scholar] [CrossRef]
- Frost, R.A.; Huber, D.; Pruznak, A.; Lang, C.H. Regulation of REDD1 by insulin-like growth factor-I in skeletal muscle and myotubes. J. Cell. Biochem. 2009, 108, 1192–1202. [Google Scholar] [CrossRef]
- Hasselgren, P.O.; Alamdari, N.; Aversa, Z.; Gonnella, P.; Smith, I.J.; Tizio, S. Corticosteroids and muscle wasting: Role of transcription factors, nuclear cofactors, and hyperacetylation. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 423–428. [Google Scholar] [CrossRef]
- Passaquin, A.C.; Lhote, P.; Ruegg, U.T. Calcium influx inhibition by steroids and analogs in C2C12 skeletal muscle cells. Br. J. Pharmacol. 1998, 124, 1751–1759. [Google Scholar] [CrossRef]
- Agell, N.; Bachs, O.; Rocamora, N.; Villalonga, P. Modulation of the Ras/Raf/MEK/ERK pathway by Ca(2+), and calmodulin. Cell. Signal. 2002, 14, 649–654. [Google Scholar] [CrossRef]
- Shi, H.; Scheffler, J.M.; Pleitner, J.M.; Zeng, C.; Park, S.; Hannon, K.M.; Grant, A.L.; Gerrard, D.E. Modulation of skeletal muscle fiber type by mitogen-activated protein kinase signaling. FASEB J. 2008, 22, 2990–3000. [Google Scholar] [CrossRef]
- Seaberg, B.; Henslee, G.; Wang, S.; Paez-Colasante, X.; Landreth, G.E.; Rimer, M. Muscle-derived extracellular signal-regulated kinases 1 and 2 are required for the maintenance of adult myofibers and their neuromuscular junctions. Mol. Cell. Biol. 2015, 35, 1238–1253. [Google Scholar] [CrossRef]
- DuBois, D.C.; Almon, R.R. Glucocorticoid sites in skeletal muscle: Adrenalectomy, maturation, fiber type, and sex. Am. J. Physiol. 1984, 247, E118–E125. [Google Scholar] [CrossRef]
- Schakman, O.; Gilson, H.; Kalista, S.; Thissen, J.P. Mechanisms of muscle atrophy induced by glucocorticoids. Horm. Res. Paediat. 2009, 72, 36–41. [Google Scholar] [CrossRef]
- Schakman, O.; Gilson, H.; Thissen, J.P. Mechanisms of glucocorticoid-induced myopathy. J. Endocrinol. 2008, 197, 1–10. [Google Scholar] [CrossRef]
- Macedo, A.G.; Krug, A.L.; Souza, L.M.; Martuscelli, A.M.; Constantino, P.B.; Zago, A.S.; Rush, J.W.; Santos, C.F.; Amaral, S.L. Time-course changes of catabolic proteins following muscle atrophy induced by dexamethasone. Steroids 2016, 107, 30–36. [Google Scholar] [CrossRef]
- Ma, K.; Mallidis, C.; Bhasin, S.; Mahabadi, V.; Artaza, J.; Gonzalez-Cadavid, N.; Arias, J.; Salehian, B. Glucocorticoid-induced skeletal muscle atrophy is associated with upregulation of myostatin gene expression. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E363–E371. [Google Scholar] [CrossRef]
- Wang, R.; Jiao, H.; Zhao, J.; Wang, X.; Lin, H. Glucocorticoids Enhance Muscle Proteolysis through a Myostatin-Dependent Pathway at the Early Stage. PLoS ONE 2016, 11, e0156225. [Google Scholar] [CrossRef]
- Gonzalez-Perez, O.; Luquin, S.; Garcia-Estrada, J.; Ramos-Remus, C. Deflazacort: A glucocorticoid with few metabolic adverse effects but important immunosuppressive activity. Adv. Ther. 2007, 24, 1052–1060. [Google Scholar] [CrossRef]
- Kasperk, C.; Schneider, U.; Sommer, U.; Niethard, F.; Ziegler, R. Differential effects of glucocorticoids on human osteoblastic cell metabolism in vitro. Calcified Tissue Int. 1995, 57, 120–126. [Google Scholar] [CrossRef]
- Ferraris, J.R.; Pasqualini, T.; Legal, S.; Sorroche, P.; Galich, A.M.; Pennisi, P.; Domene, H.; Jasper, H. Effect of deflazacort versus methylprednisone on growth, body composition, lipid profile, and bone mass after renal transplantation. The Deflazacort Study Group. Pediatr. Nephrol. 2000, 14, 682–688. [Google Scholar] [CrossRef]
- Bello, L.; Gordish-Dressman, H.; Morgenroth, L.P.; Henricson, E.K.; Duong, T.; Hoffman, E.P.; Cnaan, A.; McDonald, C.M.; Investigators, C. Prednisone/prednisolone and deflazacort regimens in the CINRG Duchenne Natural History Study. Neurology 2015, 85, 1048–1055. [Google Scholar] [CrossRef]
- Shieh, P.B.; McIntosh, J.; Jin, F.; Souza, M.; Elfring, G.; Narayanan, S.; Trifillis, P.; Peltz, S.W.; McDonald, C.M.; Darras, B.T.; et al. Deflazacort versus prednisone/prednisolone for maintaining motor function and delaying loss of ambulation: A post HOC analysis from the ACT DMD trial. Muscle Nerve 2018, 58, 639–645. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Days of Study | ||||||
|---|---|---|---|---|---|---|---|
| 1 | 30 | 32 | 34 | 36 | 38 | 40 | |
| CT | 317.0 g | 427.9 g | 436.5 g | 443.7 g | 449.5 g | 451.8 g | 461.4 g |
| DX1.25 | 339.4 g | 433.9 g | 414.1 g | 390.4 g *** | 367.4 g *** | 348.0 g *** | 336.7 g *** |
| DX2.5 | 328.1 g | 443.3 g | 422.9 g | 398.0 g ** | 374.6 g *** | 357.1 g *** | 335.5 g *** |
| DC10 | 317.5 g | 418.2 g | 380.2 g*** | 379.7 g *** | 368.5 g *** | 355.3 g *** | 353.0 g *** |
| DC20 | 320.3 g | 433.2 g | 398.4 g ** | 385.0 g*** | 365.2 g *** | 350.0 g *** | 339.8 g *** |
| MP6 | 336.6 g | 439.0 g | 400.5 g * | 368.5 g *** | 346.0 g *** | 323.4 g *** | 317.0 g *** |
| MP13 | 319.1 g | 436.0 g | 388.1 g *** | 354.8 g *** ## vs. DX2.5 | 331.4 g *** ## vs. DX2.5 | 313.1 g *** ## vs. DX2.5 # vs. DC20 | 293.4 g *** ## vs. DX2.5 ## vs. DC20 |
| Group | Tissue Collected (Grams) | ||
|---|---|---|---|
| Adrenal | GA | TA | |
| CT | 0.0415 ± 0.007 | 2.398 ± 0.344 | 0.734 ± 0.103 |
| DX1.25 | 0.0196 ± 0.002 *** | 1.613 ± 0.257 *** | 0.512 ± 0.038 *** |
| DX2.5 | 0.0208 ± 0.006 *** | 1.774 ± 0.275 *** | 0.477 ± 0.067 *** |
| MP6 | 0.0300 ± 0.012 | 1.570 ± 0.084 *** | 0.500 ± 0.030 *** |
| MP13 | 0.0216 ± 0.003 *** | 1.375 ± 0.088 ***; ## vs. DC20 and # vs. DX2.5 | 0.436 ± 0.043 ***; # vs. DC20 |
| DC10 | 0.0311 ± 0.013 | 1.919 ± 0.205 *** and # vs. MP6 | 0.575 ± 0.053 ***; ## vs. MP6 and DX1.25 |
| DC20 | 0.0227 ± 0.003 *** | 1.801 ± 0.151 *** | 0.549 ± 0.033 *** |
| Fiber 1 | P Value | Fiber 2A | P Value | Fiber 2B | P Value | |
|---|---|---|---|---|---|---|
| CT | 12.83% | P = 0.0040 ** vs. MP6 P = 0.0213 * vs. MP13 | 27.17% | P = 0.0023 ** vs. DX2.5 P = 0.0180 * vs. DC10 | 60.00% | P = 0.0296 * vs. MP13 |
| DX1.25 | 12.18% | - | 26.50% | P = 0.0006 ** vs. DX2.5 | 61.32% | - |
| DX2.5 | 10.52% | - | 34.56% | - | 54.92% | - |
| MP6 | 5.60% | P = 0.0406 * vs. DX1.25 P = 0.0464* vs. DC10 | 30.66% | P = 0.0652 vs. MP13 | 63.74% | - |
| MP13 | 5.87% | - | 25.00% | P = 0.0273 * vs. DC20 P = 0.0019 ** vs. DX2.5 | 69.13% | P = 0.0011 ** vs. DX2.5 P = 0.0010 ** vs. DC20 |
| DC10 | 8.72% | - | 32.60% | - | 58.69% | - |
| DC20 | 9.54% | - | 31.03% | - | 59.43% | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fappi, A.; Neves, J.d.C.; Sanches, L.N.; Massaroto e Silva, P.V.; Sikusawa, G.Y.; Brandão, T.P.C.; Chadi, G.; Zanoteli, E. Skeletal Muscle Response to Deflazacort, Dexamethasone and Methylprednisolone. Cells 2019, 8, 406. https://doi.org/10.3390/cells8050406
Fappi A, Neves JdC, Sanches LN, Massaroto e Silva PV, Sikusawa GY, Brandão TPC, Chadi G, Zanoteli E. Skeletal Muscle Response to Deflazacort, Dexamethasone and Methylprednisolone. Cells. 2019; 8(5):406. https://doi.org/10.3390/cells8050406
Chicago/Turabian StyleFappi, Alan, Juliana de Carvalho Neves, Leandro Nunes Sanches, Pedro Victor Massaroto e Silva, Guilherme Yuiti Sikusawa, Thayane Pereira Correa Brandão, Gerson Chadi, and Edmar Zanoteli. 2019. "Skeletal Muscle Response to Deflazacort, Dexamethasone and Methylprednisolone" Cells 8, no. 5: 406. https://doi.org/10.3390/cells8050406
APA StyleFappi, A., Neves, J. d. C., Sanches, L. N., Massaroto e Silva, P. V., Sikusawa, G. Y., Brandão, T. P. C., Chadi, G., & Zanoteli, E. (2019). Skeletal Muscle Response to Deflazacort, Dexamethasone and Methylprednisolone. Cells, 8(5), 406. https://doi.org/10.3390/cells8050406