Mitochondrial Transcriptome Control and Intercompartment Cross-Talk During Plant Development

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
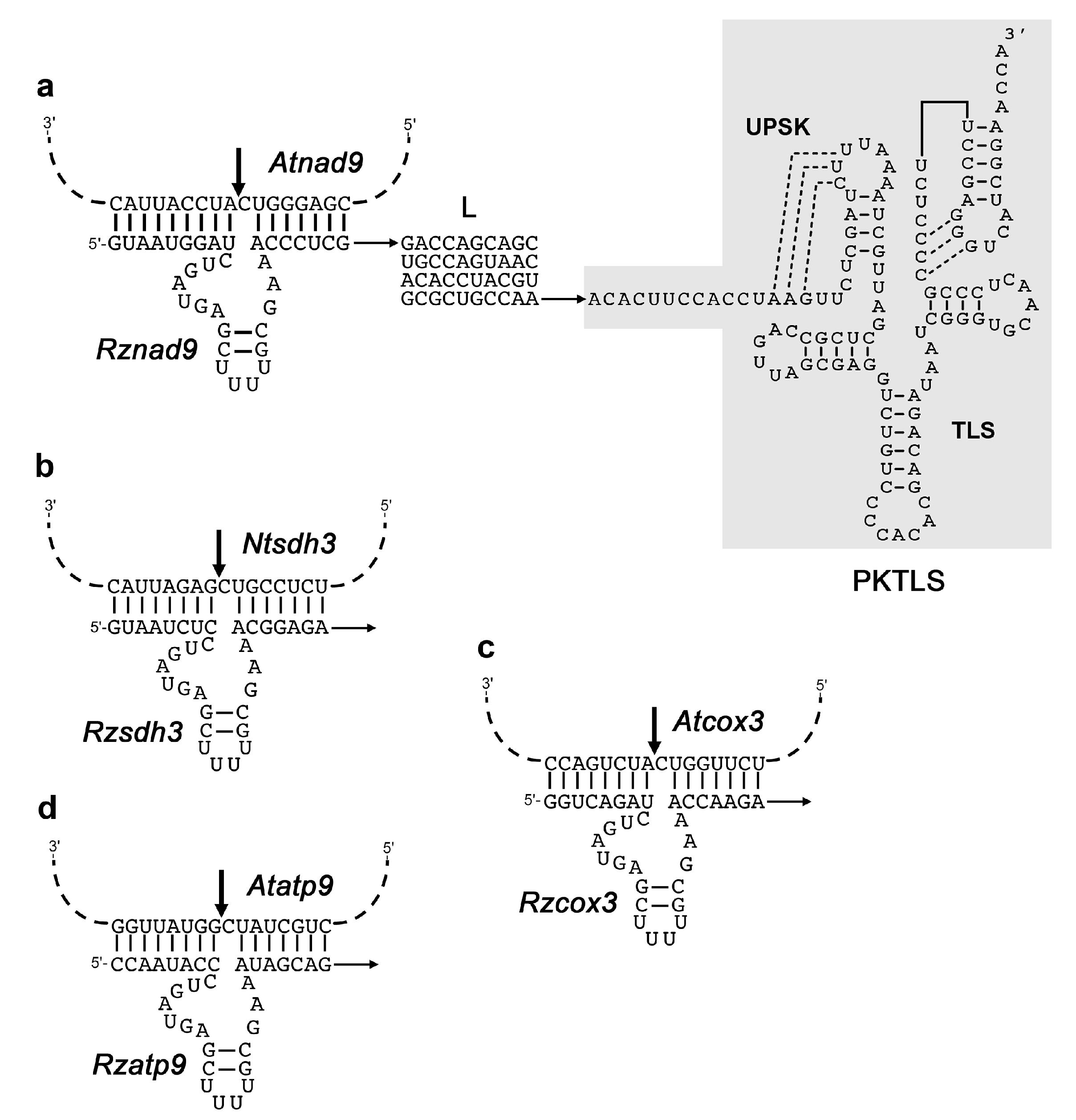
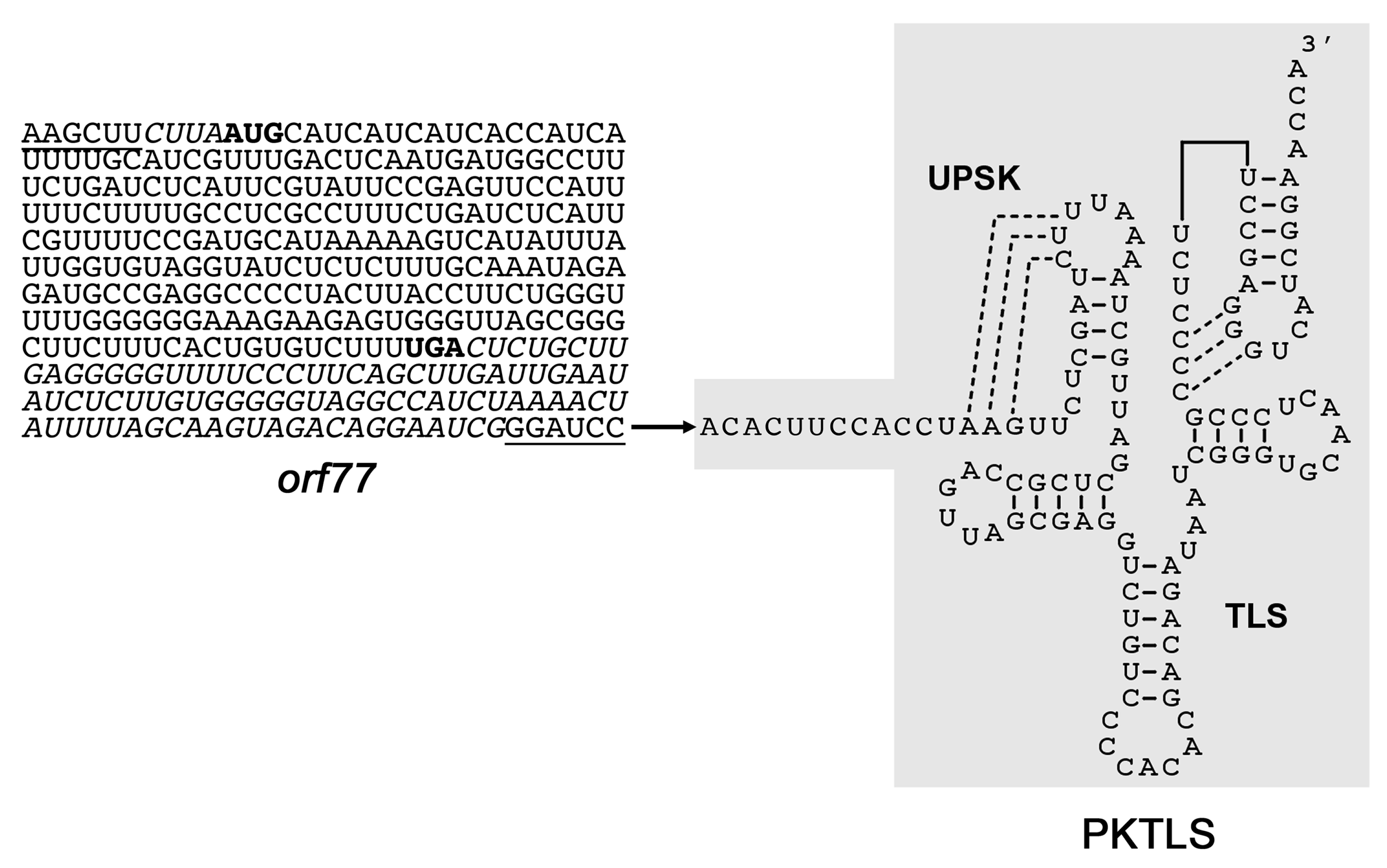
2.1. Preparation of Gene Constructs for In Vivo Expression
2.2. Nuclear Transformations
2.3. Seedling Growth and Trangene Induction
2.4. RNA Extraction, Northern Bloting and RT-qPCR Analyses
2.5. Isolation of Mitochondria and Import Assessment
2.6. Microarray Assays
2.7. Microarray Data Analyses
3. Results
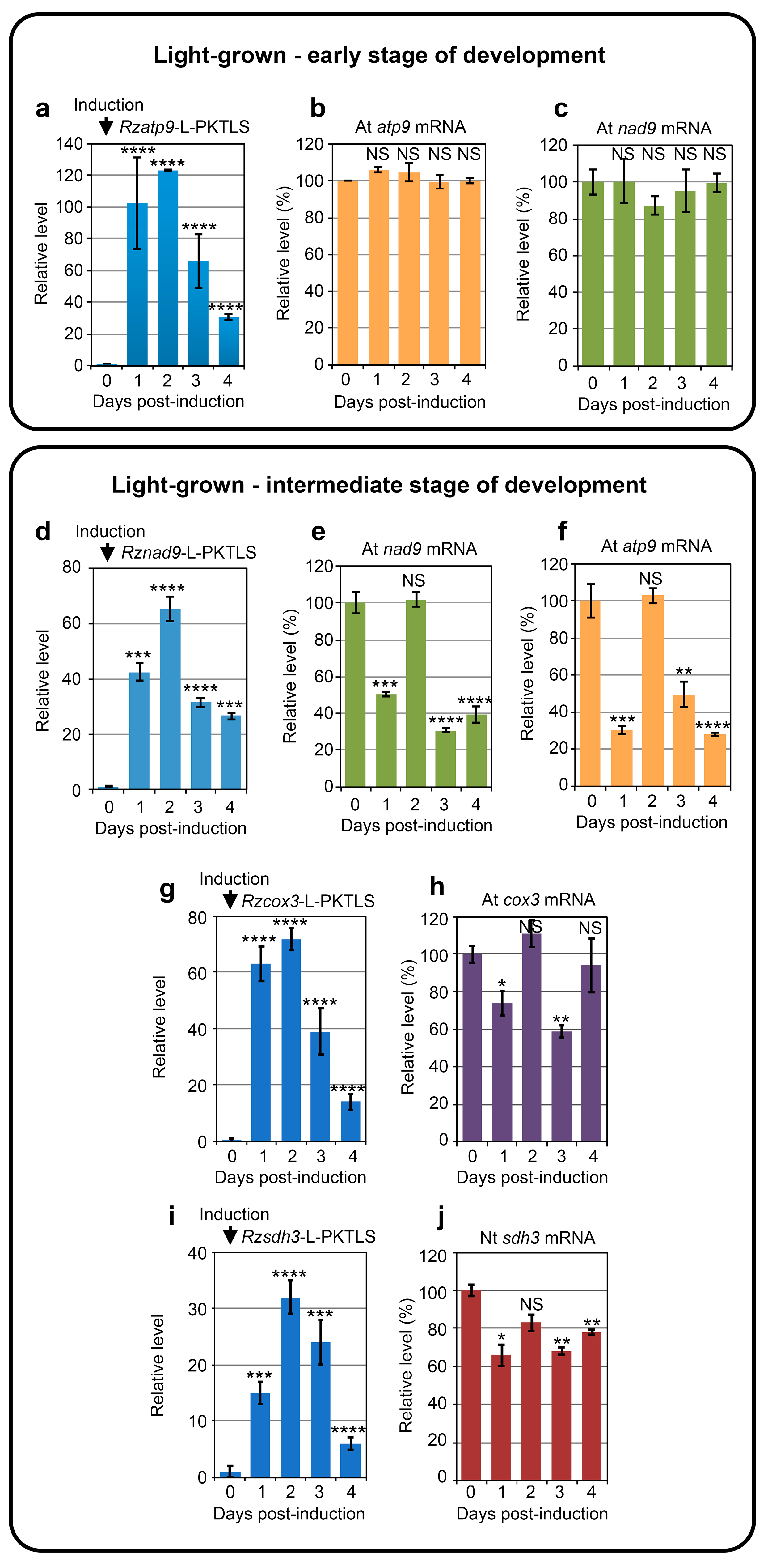
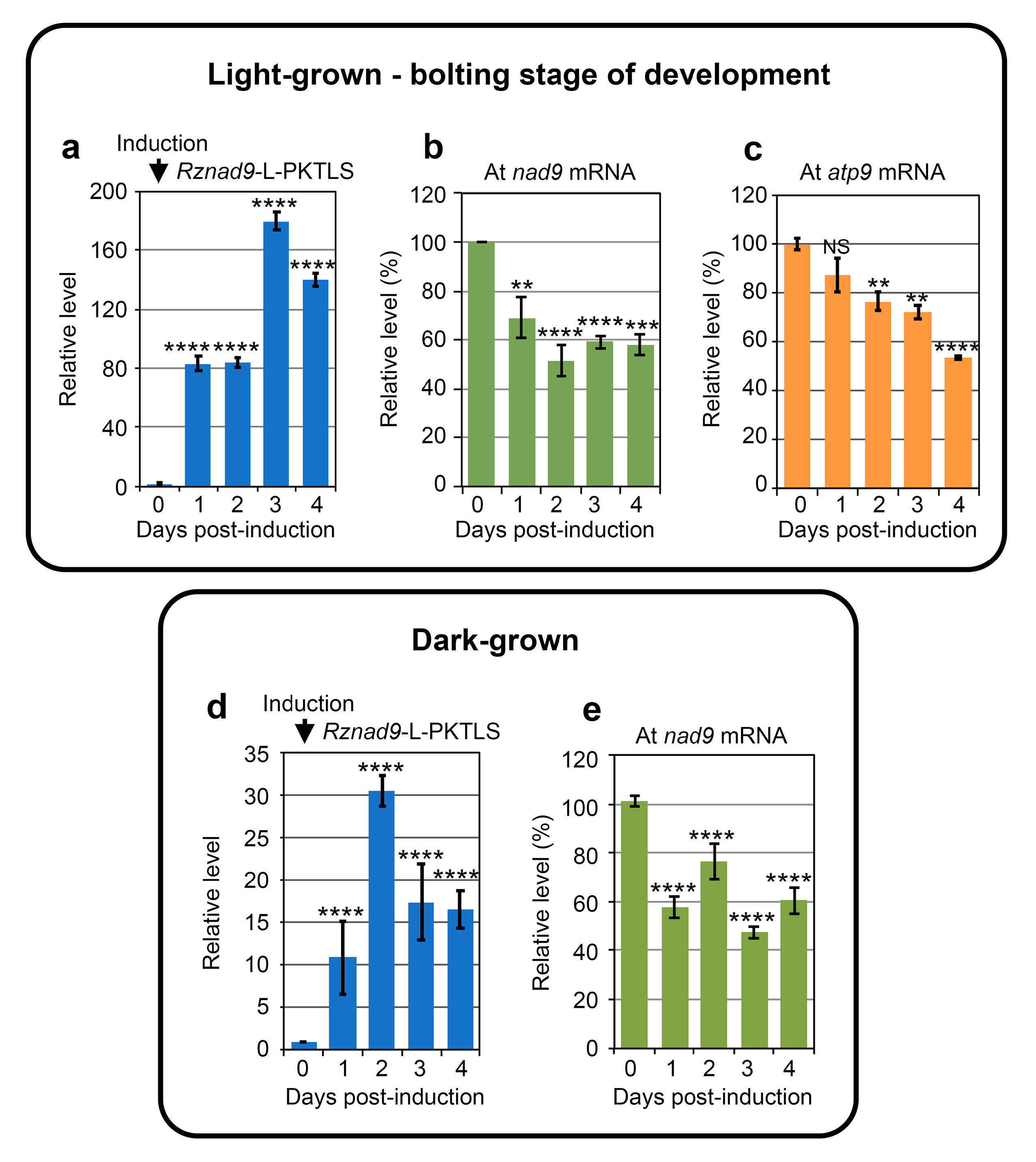
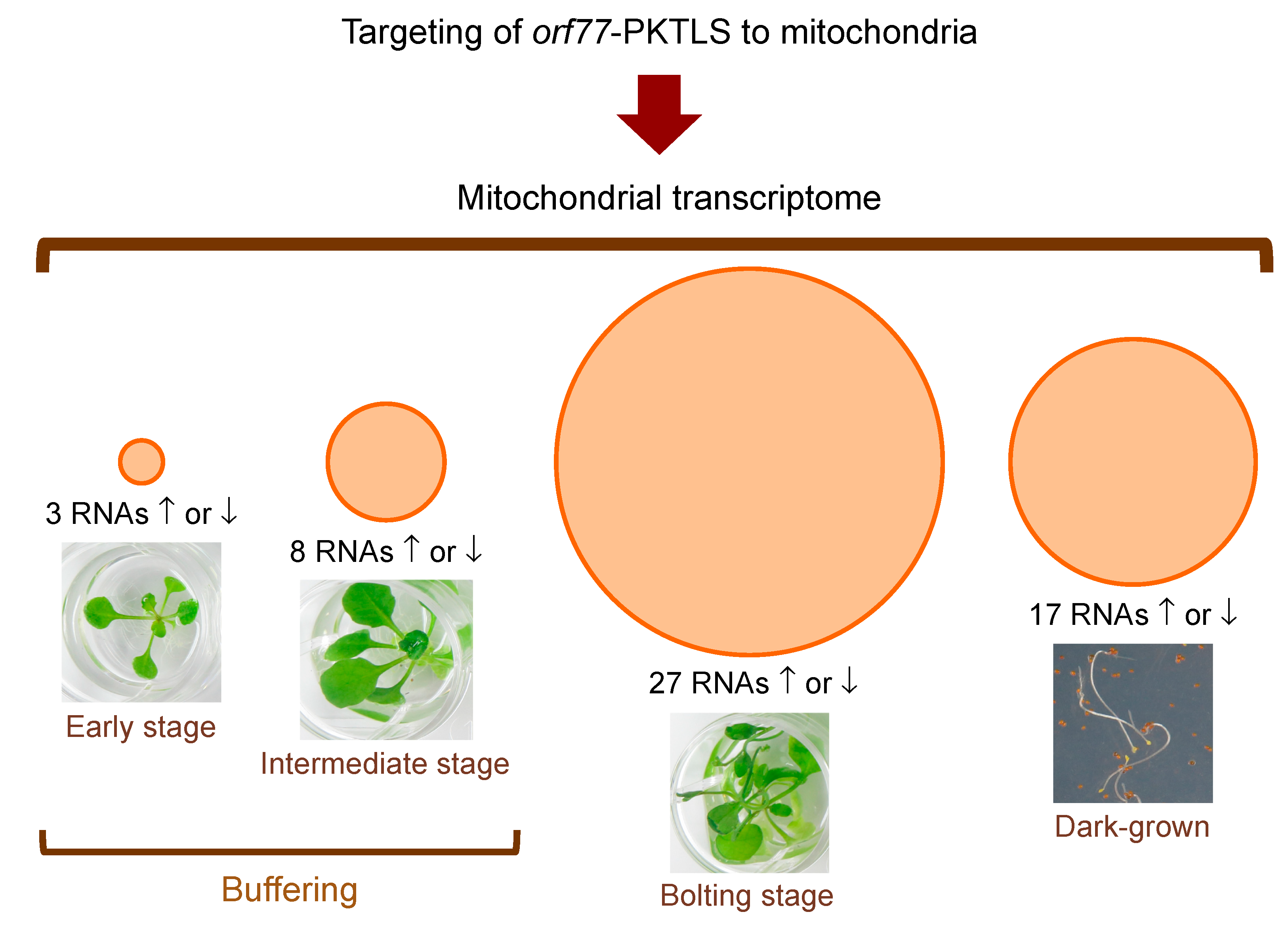
3.1. Trans-Ribozyme-Mediated Modulation of Mitochondrial RNA Steady State Levels Depends on the Plant Developmental Stage
3.2. CMS RNA-Triggered Modulation of Mitochondrial RNA Steady State Levels Depends on the Plant Developmental Stage
3.3. Ribozyme-Mediated Knockdown of a Mitochondrial mRNA Triggers a Nuclear Transcriptome Response
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shokolenko, I.N.; Alexeyev, M.F. Mitochondrial transcription in mammalian cells. Front. Biosci. 2017, 22, 835–853. [Google Scholar] [Green Version]
- Holec, S.; Lange, H.; Kühn, K.; Alioua, M.; Börner, T.; Gagliardi, D. Relaxed transcription in Arabidopsis mitochondria is counterbalanced by RNA stability control mediated by polyadenylation and polynucleotide phosphorylase. Mol. Cell. Biol. 2006, 26, 2869–2876. [Google Scholar] [CrossRef] [PubMed]
- Rackham, O.; Mercer, T.R.; Filipovska, A. The human mitochondrial transcriptome and the RNA-binding proteins that regulate its expression. Wiley Interdiscip. Rev. RNA 2012, 3, 675–695. [Google Scholar] [CrossRef] [PubMed]
- Small, I.D.; Rackham, O.; Filipovska, A. Organelle transcriptomes: Products of a deconstructed genome. Curr. Opin. Microbiol. 2013, 16, 652–658. [Google Scholar] [CrossRef] [PubMed]
- Pearce, S.F.; Rebelo-Guiomar, P.; D’Souza, A.R.; Powell, C.A.; Van Haute, L.; Minczuk, M. Regulation of mammalian mitochondrial gene expression: Recent advances. Trends Biochem. Sci. 2017, 42, 625–639. [Google Scholar] [CrossRef] [PubMed]
- Rebelo-Guiomar, P.; Powell, C.A.; Van Haute, L.; Minczuk, M. The mammalian mitochondrial epitranscriptome. Biochim. Biophys. Acta Gene Regul. Mech. 2019, 1862, 429–446. [Google Scholar] [CrossRef] [PubMed]
- Pohjoismaki, J.L.; Kruger, M.; Al-Furoukh, N.; Lagerstedt, A.; Karhunen, P.J.; Braun, T. Postnatal cardiomyocyte growth and mitochondrial reorganization cause multiple changes in the proteome of human cardiomyocytes. Mol. BioSyst. 2013, 9, 1210–1219. [Google Scholar] [CrossRef] [PubMed]
- Mposhi, A.; Van der Wijst, M.G.; Faber, K.N.; Rots, M.G. Regulation of mitochondrial gene expression, the epigenetic enigma. Front. Biosci. (Landmark Ed) 2017, 22, 1099–1113. [Google Scholar]
- Barshad, G.; Marom, S.; Cohen, T.; Mishmar, D. Mitochondrial DNA transcription and its regulation: An evolutionary perspective. Trends Genet. 2018, 34, 682–692. [Google Scholar] [CrossRef]
- Robles, P.; Quesada, V. Transcriptional and post-transcriptional regulation of organellar gene expression (OGE) and its roles in plant salt tolerance. Int. J. Mol. Sci. 2019, 20, 1056. [Google Scholar] [CrossRef]
- Woodson, J.D.; Chory, J. Coordination of gene expression between organellar and nuclear genomes. Nat. Rev. Genet. 2008, 9, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Cogliati, S.; Lorenzi, I.; Rigoni, G.; Caicci, F.; Soriano, M.E. Regulation of mitochondrial electron transport chain assembly. J. Mol. Biol. 2018, 430, 4849–4873. [Google Scholar] [CrossRef] [PubMed]
- Weinhouse, C. Mitochondrial-epigenetic crosstalk in environmental toxicology. Toxicology 2017, 391, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.; De Clercq, I.; Van Aken, O.; Law, S.R.; Ivanova, A.; Willems, P.; Giraud, E.; Van Breusegem, F.; Whelan, J. Anterograde and retrograde regulation of nuclear genes encoding mitochondrial proteins during growth, development, and stress. Mol. Plant 2014, 7, 1075–1093. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Butow, R.A. Mitochondrial retrograde signaling. Annu. Rev. Genet. 2006, 40, 159–185. [Google Scholar] [CrossRef]
- Dekkers, B.J.; Pearce, S.; van Bolderen-Veldkamp, R.P.; Marshall, A.; Widera, P.; Gilbert, J.; Drost, H.G.; Bassel, G.W.; Müller, K.; King, J.R.; et al. Transcriptional dynamics of two seed compartments with opposing roles in Arabidopsis seed germination. Plant Physiol. 2013, 163, 205–215. [Google Scholar] [CrossRef]
- Law, S.R.; Narsai, R.; Taylor, N.L.; Delannoy, E.; Carrie, C.; Giraud, E.; Millar, A.H.; Small, I.; Whelan, J. Nucleotide and RNA metabolism prime translational initiation in the earliest events of mitochondrial biogenesis during Arabidopsis germination. Plant Physiol. 2012, 158, 1610–1627. [Google Scholar] [CrossRef]
- Law, S.R.; Narsai, R.; Whelan, J. Mitochondrial biogenesis in plants during seed germination. Mitochondrion 2014, 19, 214–221. [Google Scholar] [CrossRef]
- Salinas, T.; Duchêne, A.M.; Maréchal-Drouard, L. Recent advances in tRNA mitochondrial import. Trends Biochem. Sci. 2008, 33, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Val, R.; Wyszko, E.; Valentin, C.; Szymanski, M.; Cosset, A.; Alioua, M.; Dreher, T.W.; Barciszewski, J.; Dietrich, A. Organelle trafficking of chimeric ribozymes and genetic manipulation of mitochondria. Nucleic Acids Res. 2011, 39, 9262–9274. [Google Scholar] [CrossRef] [PubMed]
- Sultan, L.D.; Mileshina, D.; Grewe, F.; Rolle, K.; Abudraham, S.; Glodowicz, P.; Niazi, A.K.; Keren, I.; Shevtsov, S.; Klipcan, L.; et al. The reverse transcriptase/RNA maturase protein MatR is required for the splicing of various group II introns in Brassicaceae mitochondria. Plant Cell 2016, 28, 2805–2829. [Google Scholar] [CrossRef] [PubMed]
- Perrotta, A.T.; Been, M.D. A pseudoknot-like structure required for efficient self-cleavage of hepatitis delta virus RNA. Nature 1991, 350, 434–436. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Niu, Q.W.; Chua, N.H. Technical advance: An estrogen receptor-based transactivator XVE mediates highly inducible gene expression in transgenic plants. Plant J. 2000, 24, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Mathews, D.H.; Sabina, J.; Zuker, M.; Turner, D.H. Expanded sequence dependence of thermodynamic parameters improves prediction of RNA secondary structure. J. Mol. Biol. 1999, 288, 911–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Engelen, F.A.; Molthoff, J.W.; Conner, A.J.; Nap, J.P.; Pereira, A.; Stiekema, W.J. pBINPLUS: An improved plant transformation vector based on pBIN19. Transgenic Res. 1995, 4, 288–290. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3–new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Sweetlove, L.J.; Taylor, N.L.; Leaver, C.J. Isolation of intact, functional mitochondria from the model plant Arabidopsis thaliana. Methods Mol. Biol. 2007, 372, 125–136. [Google Scholar]
- Lurin, C.; Andrés, C.; Aubourg, S.; Bellaoui, M.; Bitton, F.; Bruyère, C.; Caboche, M.; Debast, C.; Gualberto, J.; Hoffmann, B.; et al. Genome-wide analysis of Arabidopsis pentatricopeptide repeat proteins reveals their essential role in organelle biogenesis. Plant Cell 2004, 16, 2089–2103. [Google Scholar] [CrossRef]
- Yang, Y.H.; Dudoit, S.; Luu, P.; Lin, D.M.; Peng, V.; Ngai, J.; Speed, T.P. Normalization for cDNA microarray data: A robust composite method addressing single and multiple slide systematic variation. Nucleic Acids Res. 2002, 30, e15. [Google Scholar] [CrossRef]
- Matthew, E.R.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar]
- Smyth, G.K. Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat. Appl. Genet. Mol. Biol. 2004, 3, 3. [Google Scholar] [CrossRef] [PubMed]
- Guedj, M.; Robin, S.; Celisse, A.; Nuel, G. Kerfdr: A semi-parametric kernel-based approach to local false discovery rate estimation. BMC Bioinform. 2009, 10, 84. [Google Scholar] [CrossRef] [PubMed]
- Storey, J.D.; Tibshirani, R. Statistical significance for genomewide studies. Proc. Natl. Acad. Sci. USA 2003, 100, 9440–9445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chun, H.; Keleş, S. Sparse partial least squares regression for simultaneous dimension reduction and variable selection. J. R. Stat. Soc. Series B Stat. Methodol. 2010, 72, 3–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagnot, S.; Tamby, J.P.; Martin-Magniette, M.L.; Bitton, F.; Taconnat, L.; Balzergue, S.; Aubourg, S.; Renou, J.P.; Lecharny, A.; Brunaud, V. CATdb: A public access to Arabidopsis transcriptome data from the URGV-CATMA platform. Nucleic Acids Res. 2008, 36, D986–D990. [Google Scholar] [CrossRef] [PubMed]
- Zaag, R.; Tamby, J.P.; Guichard, C.; Tariq, Z.; Rigaill, G.; Delannoy, E.; Renou, J.P.; Balzergue, S.; Mary-Huard, T.; Aubourg, S.; et al. GEM2Net: From gene expression modeling to–Omics networks, a new CATdb module to investigate Arabidopsis thaliana genes involved in stress response. Nucleic Acids Res. 2015, 43, D1010–D1017. [Google Scholar] [CrossRef]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for functional genomics data sets--update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef]
- Brazma, A.; Hingamp, P.; Quackenbush, J.; Sherlock, G.; Spellman, P.; Stoeckert, C.; Aach, J.; Ansorge, W.; Ball, C.A.; Causton, H.C.; et al. Minimum information about a microarray experiment (MIAME)–toward standards for microarray data. Nat. Genet. 2001, 29, 365–371. [Google Scholar] [CrossRef]
- Matsuda, D.; Dreher, T.W. The tRNA-like structure of Turnip yellow mosaic virus RNA is a 3’-translational enhancer. Virology 2004, 321, 36–46. [Google Scholar] [CrossRef]
- Delage, L.; Duchêne, A.M.; Zaepfel, M.; Maréchal-Drouard, L. The anticodon and the D-domain sequences are essential determinants for plant cytosolic tRNA(Val) import into mitochondria. Plant J. 2003, 34, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, A.; Maréchal-Drouard, L.; Carneiro, V.; Cosset, A.; Small, I. A single base change prevents import of cytosolic tRNA(Ala) into mitochondria in transgenic plants. Plant J. 1996, 10, 913–918. [Google Scholar] [CrossRef]
- Matera, J.T.; Monroe, J.; Smelser, W.; Gabay-Laughnan, S.; Newton, K.J. Unique changes in mitochondrial genomes associated with reversions of S-type cytoplasmic male sterility in maizemar. PLoS ONE 2011, 6, e23405. [Google Scholar] [CrossRef] [PubMed]
- Heazlewood, J.L.; Verboom, R.E.; Tonti-Filippini, J.; Small, I.; Millar, A.H. SUBA: The Arabidopsis subcellular database. Nucleic Acids Res. 2007, 35, D213–D218. [Google Scholar] [CrossRef]
- Millar, A.H.; Whelan, J.; Soole, K.L.; Day, D.A. Organization and regulation of mitochondrial respiration in plants. Annu. Rev. Plant Biol. 2011, 62, 79–104. [Google Scholar] [CrossRef]
- Topping, J.F.; Leaver, C.J. Mitochondrial gene expression during wheat leaf development. Planta 1990, 182, 399–407. [Google Scholar] [CrossRef]
- Howell, K.A.; Millar, A.H.; Whelan, J. Ordered assembly of mitochondria during rice germination begins with pro-mitochondrial structures rich in components of the protein import apparatus. Plant Mol. Biol. 2006, 60, 201–223. [Google Scholar] [CrossRef] [PubMed]
- Howell, K.A.; Cheng, K.; Murcha, M.W.; Jenkin, L.E.; Millar, A.H.; Whelan, J. Oxygen initiation of respiration and mitochondrial biogenesis in rice. J. Biol. Chem. 2007, 282, 15619–15631. [Google Scholar] [CrossRef] [PubMed]
- Ishizaki, K.; Larson, T.R.; Schauer, N.; Fernie, A.R.; Graham, I.A.; Leaver, C.J. The critical role of Arabidopsis electron-transfer flavoprotein:ubiquinone oxidoreductase during dark-induced starvation. Plant Cell 2005, 17, 2587–2600. [Google Scholar] [CrossRef]
- Sung, T.Y.; Tseng, C.C.; Hsieh, M.H. The SLO1 PPR protein is required for RNA editing at multiple sites with similar upstream sequences in Arabidopsis mitochondria. Plant J. 2010, 63, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, W.Y.; Liao, J.C.; Chang, C.Y.; Harrison, T.; Boucher, C.; Hsieh, M.H. The SLOW GROWTH3 pentatricopeptide repeat protein is required for the splicing of mitochondrial NADH dehydrogenase subunit 7 intron 2 in Arabidopsis. Plant Physiol. 2015, 168, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Koprivova, A.; Des Francs-Small, C.C.; Calder, G.; Mugford, S.T.; Tanz, S.; Lee, B.R.; Zechmann, B.; Small, I.; Kopriva, S. Identification of a pentatricopeptide repeat protein implicated in splicing of intron 1 of mitochondrial nad7 transcripts. J. Biol. Chem. 2010, 285, 32192–32199. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, N.; Sakurai, N. A mutation in At-nMat1a, which encodes a nuclear gene having high similarity to group II intron maturase, causes impaired splicing of mitochondrial NAD4 transcript and altered carbon metabolism in Arabidopsis thaliana. Plant Cell Physiol. 2006, 47, 772–783. [Google Scholar] [CrossRef] [PubMed]
- Woo, D.K.; Phang, T.L.; Trawick, J.D.; Poyton, R.O. Multiple pathways of mitochondrial-nuclear communication in yeast: Intergenomic signaling involves ABF1 and affects a different set of genes than retrograde regulation. Biochim. Biophys. Acta 2009, 1789, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Börner, T. The discovery of plastid-to-nucleus retrograde signaling–A personal perspective. Protoplasma 2017, 254, 1845–1855. [Google Scholar] [CrossRef] [PubMed]
- Woodson, J.D.; Perez-Ruiz, J.M.; Schmit, R.J.; Ecker, J.R.; Chory, J. Sigma factor-mediated plastid retrograde signals control nuclear gene expression. Plant J. 2013, 73, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Holec, S.; Lange, H.; Canaday, J.; Gagliardi, D. Coping with cryptic and defective transcripts in plant mitochondria. Biochim. Biophys. Acta 2008, 1779, 566–573. [Google Scholar] [CrossRef]
- Nam, M.; Akie, T.E.; Sanosaka, M.; Craige, S.M.; Kant, S.; Keaney, J.F., Jr.; Cooper, M.P. Mitochondrial retrograde signaling connects respiratory capacity to thermogenic gene expression. Sci. Rep. 2017, 7, 2013. [Google Scholar] [CrossRef] [Green Version]
- da Cunha, F.M.; Torelli, N.Q.; Kowaltowski, A.J. Mitochondrial retrograde signaling: Triggers, pathways, and outcomes. Oxid. Med. Cell. Longev. 2015, 2015, 482582. [Google Scholar] [CrossRef]
- Jazwinski, S.M. The retrograde response: A conserved compensatory reaction to damage from within and from without. Prog. Mol. Biol. Transl. Sci. 2014, 127, 133–154. [Google Scholar]
- Berkowitz, O.; De Clercq, I.; Van Breusegem, F.; Whelan, J. Interaction between hormonal and mitochondrial signalling during growth, development and in plant defence responses. Plant Cell Environ. 2016, 39, 1127–1139. [Google Scholar] [CrossRef] [PubMed]
- Kühn, K.; Yin, G.; Duncan, O.; Law, S.R.; Kubiszewski-Jakubiak, S.; Kaur, P.; Meyer, E.; Wang, Y.; Colas des Francs-Small, C.; Giraud, E.; et al. Decreasing electron flux through the cytochrome and/or alternative respiratory pathways triggers common and distinct cellular responses dependent on growth conditions. Plant Physiol. 2015, 167, 228–250. [Google Scholar] [CrossRef]
- Zhang, F.; Zhao, J.; Xu, S.; Fang, W.; Chen, F.; Teng, N. MicroRNA and putative target discoveries in Chrysanthemum polyploidy breeding. Int. J. Genomics 2017, 2017, 6790478. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Yang, J.; Cheng, Q.; Mao, A.; Zhang, J.; Wang, S.; Weng, Y.; Wen, C. Comparative analysis of miRNA and mRNA abundance in determinate cucumber by high-throughput sequencing. PLoS ONE 2018, 13, e0190691. [Google Scholar]
- Scott, M.S.; Ono, M. From snoRNA to miRNA: Dual function regulatory non-coding RNAs. Biochimie 2011, 93, 1987–1992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepanov, G.A.; Filippova, J.A.; Komissarov, A.B.; Kuligina, E.V.; Richter, V.A.; Semenov, D.V. Regulatory role of small nucleolar RNAs in human diseases. Biomed. Res. Int. 2015, 2015, 206849. [Google Scholar] [CrossRef] [PubMed]
- Stortenbeker, N.; Bemer, M. The SAUR gene family: The plant’s toolbox for adaptation of growth and development. J. Exp. Bot. 2019, 70, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, Y.X.; Li, C.; Shi, Y.; Song, Y.; Zhang, D.; Li, Y.; Wang, T. Genome-wide analysis of the pentatricopeptide repeat gene family in different maize genomes and its important role in kernel development. BMC Plant Biol. 2018, 18, 366. [Google Scholar] [CrossRef] [PubMed]
- Gorchs Rovira, A.; Smith, A.G. PPR proteins–Orchestrators of organelle RNA metabolism. Physiol. Plant 2019. [Google Scholar] [CrossRef]
- Xing, H.; Fu, X.; Yang, C.; Tang, X.; Guo, L.; Li, C.; Xu, C.; Luo, K. Genome-wide investigation of pentatricopeptide repeat gene family in poplar and their expression analysis in response to biotic and abiotic stresses. Sci. Rep. 2018, 8, 2817. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conditions and Developmental Stage | Day 1 | Day 2 | Day 3 | Day 4 |
|---|---|---|---|---|
| Light Early stage | ↓nad5a* | |||
| ↓atp4** | ||||
| ↑rps7* | ||||
| ↑AOX1d* | ↓AOX1d*** | ↓AOX1d*** | ||
| Light Intermediate stage | ↑cob** | ↑rps7* | ||
| ↑nad5a** | ↓ccmFN2*** | ↑rpl2** | ||
| ↑nad5b** | ↑atp4* | ↑rpl5* | ||
| ↓ccmFN2** | ↑rpl2* | |||
| ↑rpl5* | ||||
| ↑AOX1d*** | ↑AOX1d*** | |||
| Light Bolting stage | ↓nad2b** | ↓atp4* | ↑nad1b* | |
| ↑nad5a** | ↓atp9**** | ↓nad2b* | ||
| ↓nad5b** | ↓rps4** | ↓nad4** | ||
| ↓nad9** | ↓rps7**** | ↑nad5a*** | ||
| ↓cox1** ↓ccmB** | ↓rpl2**** | ↓nad5b* | ||
| ↓mttB* | ↓rpl5** | ↓nad6* | ||
| ↓rpl5* | ↓rrn18** | ↓nad9** | ||
| ↓cob*** | ||||
| ↓cox1* | ↓cox1**** | |||
| ↓cox2** | ↓atp9* | |||
| ↓cox3**** | ↓ccmFC*** | |||
| ↑atp9* | ↑mttB* | |||
| ↓ccmB*** | ↑rpl5** | |||
| ↓ccmFC* | ↓rrn26* | |||
| ↓ccmFN1*** | ||||
| ↓ccmFN2** | ||||
| ↓mttB** | ||||
| ↓rrn18*** | ||||
| ↓rrn26** | ||||
| ↓AOX1a* | ||||
| ↓AOX1d** | ↓AOX1d* | ↓AOX1d* | ||
| Dark | ↑nad2b** | ↑nad4* | ↓nad1b*** | ↓ccmFC*** |
| ↓nad5a** | ↑nad5b* | ↓nad2b** | ↓matR* | |
| ↑nad5b**** | ↑cox1* | ↑nad5b** | ↓mttB* | |
| ↑nad9* | ↓ccmFN2** | ↓ccmFN2** | ↓rrn18**** | |
| ↓cox2* | ↓mttB** | ↑rrn26** | ||
| ↓matR* | ↓rrn26** | |||
| ↑rps7** | ||||
| ↓rpl5* | ||||
| ↓rrn26* | ||||
| ↑AOX1d** |
| Modulation | Annotation / Function / Organellar Localization | Gene |
|---|---|---|
| Upregulated (opposite to nad9) | Basic helix-loop-helix (bHLH) DNA-binding superfamily protein; transcription factor; response to ethylene | AT1G05710 |
| ABI3-interacting protein 3, AIP3, PFD4, PREFOLDIN 4; protein chaperone; ABI3 is an auxin-inducible transcription factor | AT1G08780 | |
| ATMYB60, Myb domain protein 60, MYB60; transcription factor; response to abscisic acid, jasmonic acid, salicylic acid | AT1G08810 | |
| Agamous-like 87, AGL87, MADS-box family protein; transcription factor | AT1G22590 | |
| GL2, GLABRA 2, HD-ZIP IV family of homeobox-leucine zipper protein with lipid-binding START domain; transcription factor | AT1G79840 | |
| DREB subfamily A-6 of ERF/AP2 transcription factor family; one AP2 domain; ethylene-activated | AT2G22200 | |
| AP2/B3-like transcriptional factor family protein; transcription factor | AT2G33720 | |
| FMA (FAMA), basic helix-loop-helix (bHLH) DNA-binding superfamily protein; transcription factor/ transcriptional activator | AT3G24140 | |
| EDF3, ethylene response DNA-binding factor 3; transcription factor; AP2 domain; ethylene responding | AT3G25730 | |
| IAA30, indole-3-acetic acid inducible protein 30; transcription factor; response to auxin | AT3G62100 | |
| B-BOX domain protein 23, BBX23; transcription factor | AT4G10240 | |
| PUCHI, ethylene response factor (ERF) subfamily B-1 of ERF/AP2 transcription factor family; one AP2 domain; ethylene response | AT5G18560 | |
| ARGOS, Auxin-regulated gene involved in organ size; response to ethylene, auxin; membrane, cytoplasm, mitochondrion | AT3G59900 | |
| EMB3103, Embryo-defective 3103, PDM2, Pigment-Defective Mutant2; pentatricopeptide repeat (PPR) superfamily protein; endonuclease; chloroplast/mitochondrion | AT1G10910 | |
| RNH1C, RNase H family protein, RNase H domain-containing protein; chloroplast | AT1G24090 | |
| RING-finger, DEAD-like helicase, PHD and SNF2 domain-containing protein | AT2G40770 | |
| AT-SR34B, Serine/arginine-rich protein splicing factor 34B, SR34B | AT4G02430 | |
| Mitochondrial nuclease 1, MNU1; putative endonuclease or glycosyl hydrolase; mitochondrial RNA 5'-end processing; chloroplast/mitochondrion | AT5G64710 | |
| EMB2394, Embryo-defective 2394; structural constituent of chloroplast ribosome; response to cytokinin | AT1G05190 | |
| rRNA, cytosolic small ribosomal subunit | AT2G01010 | |
| 5.8S rRNA, cytosolic large ribosomal subunit | AT2G01020 | |
| 40S ribosomal protein S23 (RPS23A), ribosomal protein S12/S23 family protein; cytosolic small ribosomal subunit | AT3G09680 | |
| Ribosomal protein L15, RPL15; large subunit of the chloroplast ribosome; response to cytokinin | AT3G25920 | |
| EMB3126, Embryo-defective 3126, plastid ribosomal protein L1, PRPL1; ribosomal protein L1p/L10e family; chloroplast large ribosomal subunit | AT3G63490 | |
| 60S ribosomal protein L31, RPL31B; cytosolic large ribosomal subunit | AT4G26230 | |
| U3 ribonucleoprotein, Utp family protein; rRNA processing | AT5G08600 | |
| MA3 domain-containing translation regulatory factor 1, MRF1; colocalizes with cytosolic large ribosomal subunit; isomerase activity | AT5G63190 | |
| snoRNA | AT1G03743; AT1G19373; AT1G19376; AT1G75166; AT2G35387; AT3G27865; AT3G47342; AT3G47347; AT3G58193; AT3G58196; AT4G39366; AT5G44286 |
| Modulation | Annotation/Function/Organellar Localization | Gene |
|---|---|---|
| Downregulated (as nad9) | MicroRNA414, mir414, short open reading frame 16, SORF16; miRNA; identified as a translated small open reading frame by ribosome profiling | AT1G67195 |
| B-BOX domain protein 25, BBX25, Salt tolerance homologue, STH; transcription factor; zinc ion binding | AT2G31380 | |
| ATIBH1, IBH1, ILI1 binding BHLH 1 ILI1 binding bHLH 1; transcription factor; brassinosteroid signaling; gibberellic acid signaling | AT2G43060 | |
| Arabidopsis thaliana response regulator 2, ARR5, ATRR2, IBC6, induced by cytokinin 6, response regulator 5, RR5; transcription repressor; cytokinin signaling | AT3G48100 | |
| Basic helix-loop-helix (bHLH) DNA-binding superfamily protein; transcription factor | AT4G01460 | |
| KELP; homodimers or heterodimers with the kiwi protein; transcriptional co-activator | AT4G10920 | |
| WRKY DNA-binding protein 24, ATWRKY24, WRKY24; WRKY transcription factor group II-c | AT5G41570 | |
| BOA, Brother of lux ARRHYTHMO; transcription factor; circadian clock; mRNA cell-to-cell mobile | AT5G59570 | |
| SAUR53, small auxin-upregulated RNA 53, SAUR-like auxin-responsive protein family; mitochondrion | AT1G19840 | |
| SAUR65, small auxin-upregulated RNA 65, SAUR-like auxin-responsive protein family; membrane; mitochondrion | AT1G29460 | |
| ATCLE19, CLAVATA3/ESR-related 19, CLE19, embryo surrounding region 19, ESR19; receptor binding; signal transduction; mitochondrion | AT3G24225 | |
| VQ motif-containing protein 29, VQ29; response to hypoxia | AT4G37710 | |
| RNA-binding (RRM/RBD/RNP motifs) family protein | AT1G33470 | |
| ATRNS1, Ribonuclease 1, RNS1; endoribonuclease | AT2G02990 | |
| Eukaryotic translation initiation factor 2 (eIF-2) family protein; cytosol | AT1G76820 | |
| EMB3113, Embryo-defective 3113, ribosomal protein S5, RPS5, SCA1, SCABRA 1; structural component of the 70S chloroplast ribosome; mitochondrial small ribosomal subunit | AT2G33800 | |
| U3 containing 90S pre-ribosomal complex subunit | AT2G43110 | |
| ATRAB8D, ATRABE1B, RAB GTPase homolog E1B, RABE1B; translation elongation factor; membrane; chloroplast | AT4G20360 | |
| snoRNA | AT2G35382; AT2G43137; AT2G43138; AT2G43139; AT2G43141; AT4G02550; AT4G02555; AT4G13245 |
| Modulation | Annotation/Function/Organellar Localization | Gene |
|---|---|---|
| Upregulated (opposite to nad9) | Small nuclear RNA U6acat, mRNA splicing | AT5G40395 |
| Downregulated (as nad9) | ANAC028, NAC domain-containing protein 28, NAC028; transcription factor | AT1G65910 |
| Transcription elongation factor Spt5; KOW domain | AT2G34210 | |
| MYR2, homeodomain-like superfamily protein; transcription factor | AT3G04030 | |
| HVA22-like protein F, HVA22F; membrane protein; response to abscisic acid | AT2G42820 | |
| ALY2 RNA-binding (RRM/RBD/RNP motifs) family protein; mRNA transport | AT5G02530 |
| Modulation | Annotation/Function/Organellar Localization | Gene |
|---|---|---|
| Upregulated (opposite to nad9) | auxin-responsive family protein (SAUR72) | AT3G12830 |
| auxin-responsive protein, putative (SAUR9) | AT4G36110 | |
| auxin-responsive family protein (SAUR41) | AT1G16510 | |
| auxin-responsive protein-related (SAUR77) | AT1G17345 | |
| auxin-responsive family protein (SAUR53); mitochondrion | AT1G19840 | |
| auxin-responsive family protein (SAUR71) | AT1G56150 | |
| auxin-responsive family protein (SAUR52); mitochondrion | AT1G75590 | |
| auxin-responsive protein-related (SAUR36); mitochondrion/nucleus (nucleus confirmed by GFP targeting experiments) | AT2G45210 | |
| auxin-responsive family protein (SAUR59); mitochondrion | AT3G60690 | |
| auxin-responsive protein, putative (SAUR25); mitochondrion | AT4G13790 | |
| auxin-responsive family protein (SAUR1); chloroplast | AT4G34770 | |
| auxin-responsive protein, putative (SAUR23) | AT5G18060 | |
| MIF1 (MINI ZINC FINGER 1); transcription factor; response to abscisic acid, auxin, brassinosteroid, cytokinin, gibberellin; nucleus (cytosol reported from GFP data) | AT1G74660 | |
| PLS (POLARIS), cytokinin and auxin responses; mitochondrion | AT4G39403 | |
| Tetratricopeptide repeat (TPR)-like superfamily protein; mitochondrion | AT1G28690 | |
| S-RBP11, SMALL RNA-BINDING PROTEIN 11; salt stress response; chloroplast | AT5G06210 | |
| Downregulated (as nad9) | ARGAH2, Arginine amidohydrolase 2, response to jasmonate; chloroplast/mitochondrion (confirmed by MS data) | AT4G08870 |
| Tetratricopeptide repeat (TPR)-like superfamily protein; chloroplast/mitochondrion (chloroplast confirmed by MS data) | AT2G37230 | |
| pentatricopeptide (PPR) repeat-containing protein | AT3G62470 | |
| pentatricopeptide (PPR) repeat-containing protein | AT4G01030 | |
| EMB1417 (embryo-defective 1417), PPR protein; RNA binding; endonuclease activity | AT4G21190 | |
| pentatricopeptide (PPR) repeat-containing protein | AT4G21880 | |
| pentatricopeptide (PPR) repeat-containing protein | AT5G65560 | |
| EMB1586, Embryo-defective 1586, increased size exclusion limit 1, ISE1; DEAD-box RNA helicase; chloroplast/mitochondrion (mitochondrion confirmed by GFP data) | AT1G12770 | |
| AGS1, AHG2-1 suppressor 1, bacterial-type poly(A) polymerase; mRNA polyadenylation; chloroplast/mitochondrion | AT2G17580 | |
| ATTRM2A, TRM2A, tRNA methyltransferase 2A, RNA methyltransferase family protein; cytoplasm | AT3G21300 | |
| ATP-dependent RNA helicase | AT5G39840 | |
| ARFB1A, ATARFB1A (ADP-ribosylation factor B1A); GTP binding; protein transport; Golgi apparatus | AT2G15310 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niazi, A.K.; Delannoy, E.; Iqbal, R.K.; Mileshina, D.; Val, R.; Gabryelska, M.; Wyszko, E.; Soubigou-Taconnat, L.; Szymanski, M.; Barciszewski, J.; et al. Mitochondrial Transcriptome Control and Intercompartment Cross-Talk During Plant Development. Cells 2019, 8, 583. https://doi.org/10.3390/cells8060583
Niazi AK, Delannoy E, Iqbal RK, Mileshina D, Val R, Gabryelska M, Wyszko E, Soubigou-Taconnat L, Szymanski M, Barciszewski J, et al. Mitochondrial Transcriptome Control and Intercompartment Cross-Talk During Plant Development. Cells. 2019; 8(6):583. https://doi.org/10.3390/cells8060583
Chicago/Turabian StyleNiazi, Adnan Khan, Etienne Delannoy, Rana Khalid Iqbal, Daria Mileshina, Romain Val, Marta Gabryelska, Eliza Wyszko, Ludivine Soubigou-Taconnat, Maciej Szymanski, Jan Barciszewski, and et al. 2019. "Mitochondrial Transcriptome Control and Intercompartment Cross-Talk During Plant Development" Cells 8, no. 6: 583. https://doi.org/10.3390/cells8060583
APA StyleNiazi, A. K., Delannoy, E., Iqbal, R. K., Mileshina, D., Val, R., Gabryelska, M., Wyszko, E., Soubigou-Taconnat, L., Szymanski, M., Barciszewski, J., Weber-Lotfi, F., Gualberto, J. M., & Dietrich, A. (2019). Mitochondrial Transcriptome Control and Intercompartment Cross-Talk During Plant Development. Cells, 8(6), 583. https://doi.org/10.3390/cells8060583