DAF-16/FoxO in Caenorhabditis elegans and Its Role in Metabolic Remodeling


{kind=link}
Abstract
:1. Introduction
2. Identification of DAF-16: The Genetics of Dauer Arrest
3. DAF-16/FoxO Structure and Isoforms
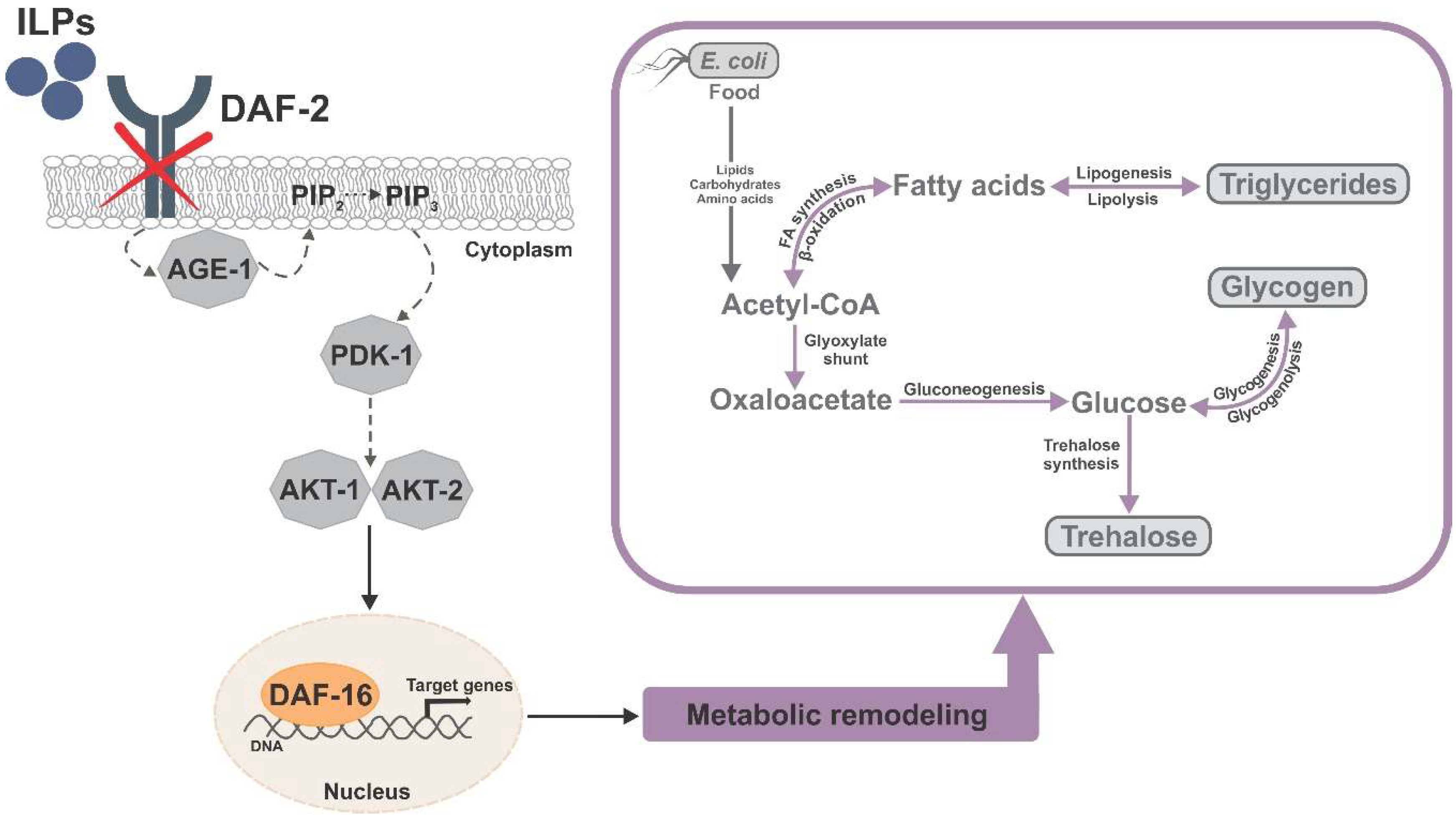
4. The IIS Pathway: A Central Regulator of DAF-16 Activity
5. IIS-Independent Regulation of DAF-16 Activity
6. Identification of DAF-16 Targets
7. DAF-16-Mediated Enhanced Stress Resistance in Long-Lived daf-2 Mutants
8. The Metabolism in daf-2 Mutants is Extensively Remodeled
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Stiernagle, T. Maintenance of C. elegans. Available online: http://www.wormbook.org/chapters/www_strainmaintain/strainmaintain.html (accessed on 31 December 2019).
- C. elegans Sequencing Consortium. Genome sequence of the nematode C. elegans: A platform for investigating biology. Science 1998, 282, 2012–2018. [Google Scholar] [CrossRef]
- Kamath, R.S.; Fraser, A.G.; Dong, Y.; Poulin, G.; Durbin, R.; Gotta, M.; Kanapin, A.; Le Bot, N.; Moreno, S.; Sohrmann, M.; et al. Systematic functional analysis of the Caenorhabditis elegans genome using rnai. Nature 2003, 421, 231–237. [Google Scholar] [CrossRef]
- Kutscher, L.M.; Shaham, S. Forward and Reverse Mutagenesis in C. elegans. Available online: http://www.wormbook.org/chapters/www_frmutagenesis/frmutagenesis.html (accessed on 31 December 2019).
- Kenyon, C.; Chang, J.; Gensch, E.; Rudner, A.; Tabtiang, R. A C. elegans mutant that lives twice as long as wild type. Nature 1993, 366, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Dorman, J.B.; Rodan, A.; Kenyon, C. Daf-16: An hnf-3/forkhead family member that can function to double the life-span of Caenorhabditis elegans. Science 1997, 278, 1319–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogg, S.; Paradis, S.; Gottlieb, S.; Patterson, G.I.; Lee, L.; Tissenbaum, H.A.; Ruvkun, G. The fork head transcription factor daf-16 transduces insulin-like metabolic and longevity signals in C. elegans. Nature 1997, 389, 994–999. [Google Scholar] [CrossRef] [PubMed]
- Albert, P.S.; Brown, S.J.; Riddle, D.L. Sensory control of dauer larva formation in Caenorhabditis elegans. J. Comp. Neurol. 1981, 198, 435–451. [Google Scholar] [CrossRef] [PubMed]
- Riddle, D.L.; Swanson, M.M.; Albert, P.S. Interacting genes in nematode dauer larva formation. Nature 1981, 290, 668–671. [Google Scholar] [CrossRef]
- Braendle, C.; Milloz, J.; Félix, M.A. Mechanisms and evolution of environmental responses in Caenorhabditis elegans. In Current Topics in Developmental Biology; Academic Press: Cambridge, MA, USA, 2007; Volume 80, pp. 171–207. [Google Scholar]
- Félix, M.-A.; Braendle, C. The natural history of Caenorhabditis elegans. Curr. Biol. 2010, 20, R965–R969. [Google Scholar] [CrossRef]
- Golden, J.W.; Riddle, D.L. The Caenorhabditis elegans dauer larva: Developmental effects of pheromone, food, and temperature. Dev. Biol. 1984, 102, 368–378. [Google Scholar] [CrossRef]
- Hu, P.J. Dauer. Available online: http://www.wormbook.org/chapters/www_dauer/dauer.html (accessed on 31 December 2019).
- Klass, M.; Hirsh, D. Non-ageing developmental variant of Caenorhabditis elegans. Nature 1976, 260, 523–525. [Google Scholar] [CrossRef]
- Anderson, G.L. Responses of dauerlarvae of Caenorhabditis elegans (nematoda: Rhabditidae) to thermal stress and oxygen deprivation. Can. J. Zool. 1978, 56, 1786–1791. [Google Scholar] [CrossRef]
- Vowels, J.J.; Thomas, J.H. Genetic analysis of chemosensory control of dauer formation in Caenorhabditis elegans. Genetics 1992, 130, 105–123. [Google Scholar] [PubMed]
- Klass, M.R. A method for the isolation of longevity mutants in the nematode Caenorhabditis elegans and initial results. Mech. Ageing Dev. 1983, 22, 279–286. [Google Scholar] [CrossRef]
- Friedman, D.B.; Johnson, T.E. A mutation in the age-1 gene in Caenorhabditis elegans lengthens life and reduces hermaphrodite fertility. Genetics 1988, 118, 75–86. [Google Scholar] [PubMed]
- Malone, E.A.; Inoue, T.; Thomas, J.H. Genetic analysis of the roles of daf-28 and age-1 in regulating Caenorhabditis elegans dauer formation. Genetics 1996, 143, 1193–1205. [Google Scholar] [PubMed]
- Morris, J.Z.; Tissenbaum, H.A.; Ruvkun, G. A phosphatidylinositol-3-oh kinase family member regulating longevity and diapause in Caenorhabditis elegans. Nature 1996, 382, 536–539. [Google Scholar] [CrossRef]
- Kimura, K.D.; Tissenbaum, H.A.; Liu, Y.; Ruvkun, G. Daf-2, an insulin receptor-like gene that regulates longevity and diapause in Caenorhabditis elegans. Science 1997, 277, 942–946. [Google Scholar] [CrossRef]
- Weigel, D.; Jürgens, G.; Küttner, F.; Seifert, E.; Jäckle, H. The homeotic gene fork head encodes a nuclear protein and is expressed in the terminal regions of the Drosophila embryo. Cell 1989, 57, 645–658. [Google Scholar] [CrossRef] [Green Version]
- Hannenhalli, S.; Kaestner, K.H. The evolution of fox genes and their role in development and disease. Nat. Rev. Genet. 2009, 10, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Accili, D.; Arden, K.C. Foxos at the crossroads of cellular metabolism, differentiation, and transformation. Cell 2004, 117, 421–426. [Google Scholar] [CrossRef] [Green Version]
- Greer, E.L.; Brunet, A. Foxo transcription factors at the interface between longevity and tumor suppression. Oncogene 2005, 24, 7410–7425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Tindall, D.J. Dynamic foxo transcription factors. J. Cell Sci. 2007, 120, 2479–2487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaestner, K.H.; Knochel, W.; Martinez, D.E. Unified nomenclature for the winged helix/forkhead transcription factors. Genes Dev. 2000, 14, 142–146. [Google Scholar]
- Kramer, J.M.; Davidge, J.T.; Lockyer, J.M.; Staveley, B.E. Expression of Drosophila foxo regulates growth and can phenocopy starvation. BMC Dev. Biol. 2003, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.Y.N.; Hench, J.; Ruvkun, G. Regulation of C. elegans daf-16 and its human ortholog fkhrl1 by the daf-2 insulin-like signaling pathway. Curr. Biol. 2001, 11, 1950–1957. [Google Scholar] [CrossRef] [Green Version]
- Lin, K.; Hsin, H.; Libina, N.; Kenyon, C. Regulation of the Caenorhabditis elegans longevity protein daf-16 by insulin/igf-1 and germline signaling. Nat. Genet. 2001, 28, 139–145. [Google Scholar] [CrossRef]
- Kwon, E.-S.; Narasimhan, S.D.; Yen, K.; Tissenbaum, H.A. A new daf-16 isoform regulates longevity. Nature 2010, 466, 498–502. [Google Scholar] [CrossRef] [Green Version]
- Bansal, A.; Kwon, E.-S.; Conte, D.; Liu, H.; Gilchrist, M.J.; MacNeil, L.T.; Tissenbaum, H.A. Transcriptional regulation of Caenorhabditis elegans foxo/daf-16 modulates lifespan. Longev. Healthspan 2014, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Christensen, R.; De La Torre-Ubieta, L.; Bonni, A.; Colón-Ramos, D.A. A conserved pten/foxo pathway regulates neuronal morphology during C. elegans development. Development 2011, 138, 5257–5367. [Google Scholar] [CrossRef] [Green Version]
- Chen, A.T.-Y.; Guo, C.; Itani, O.A.; Budaitis, B.G.; Williams, T.W.; Hopkins, C.E.; McEachin, R.C.; Pande, M.; Grant, A.R.; Yoshina, S.; et al. Longevity genes revealed by integrative analysis of isoform-specific daf-16/foxo mutants of Caenorhabditis elegans. Genetics 2015, 201, 613–629. [Google Scholar] [CrossRef] [Green Version]
- Furuyama, T.; Nakazawa, T.; Nakano, I.; Mori, N. Identification of the differential distribution patterns of mrnas and consensus binding sequences for mouse daf-16 homologues. Biochem. J. 2000, 349, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.T.; McCarroll, S.A.; Bargmann, C.I.; Fraser, A.; Kamath, R.S.; Ahringer, J.; Li, H.; Kenyon, C. Genes that act downstream of daf-16 to influence the lifespan of Caenorhabditis elegans. Nature 2003, 424, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Schuster, E.; McElwee, J.J.; Tullet, J.M.A.; Doonan, R.; Matthijssens, F.; Reece-Hoyes, J.S.; Hope, I.A.; Vanfleteren, J.R.; Thornton, J.M.; Gems, D. Damid in C. elegans reveals longevity-associated targets of daf-16/foxo. Mol. Syst. Biol. 2010, 6, 399. [Google Scholar] [CrossRef]
- McElwee, J.J.; Schuster, E.; Blanc, E.; Thomas, J.H.; Gems, D. Shared transcriptional signature in Caenorhabditis elegans dauer larvae and long-lived daf-2 mutants implicates detoxification system in longevity assurance. J. Biol. Chem. 2004, 279, 44533–44543. [Google Scholar] [CrossRef] [Green Version]
- Tepper, R.G.; Ashraf, J.; Kaletsky, R.; Kleemann, G.; Murphy, C.T.; Bussemaker, H.J. Pqm-1 complements daf-16 as a key transcriptional regulator of daf-2-mediated development and longevity. Cell 2013, 154, 676–690. [Google Scholar] [CrossRef] [Green Version]
- Clancy, D.J.; Gems, D.; Harshman, L.G.; Oldham, S.; Stocker, H.; Hafen, E.; Leevers, S.J.; Partridge, L. Extension of life-span by loss of chico, a Drosophila insulin receptor substrate protein. Science 2001, 292, 104–106. [Google Scholar] [CrossRef] [Green Version]
- Tatar, M.; Kopelman, A.; Epstein, D.; Tu, M.P.; Yin, C.M.; Garofalo, R.S. A mutant Drosophila insulin receptor homolog that extends life-span and impairs neuroendocrine function. Science 2001, 292, 107–110. [Google Scholar] [CrossRef] [Green Version]
- Holzenberger, M.; Dupont, J.; Ducos, B.; Leneuve, P.; Géloën, A.; Even, P.C.; Cervera, P.; Le Bouc, Y. Igf-1 receptor regulates lifespan and resistance to oxidative stress in mice. Nature 2003, 421, 182–187. [Google Scholar] [CrossRef]
- Blüher, M.; Kahn, B.B.; Kahn, C.R. Extended longevity in mice lacking the insulin receptor in adipose tissue. Science 2003, 299, 572–574. [Google Scholar] [CrossRef] [Green Version]
- Pierce, S.B.; Costa, M.; Wisotzkey, R.; Devadhar, S.; Homburger, S.A.; Buchman, A.R.; Ferguson, K.C.; Heller, J.; Platt, D.M.; Pasquinelli, A.A.; et al. Regulation of daf-2 receptor signaling by human insulin and ins-1, a member of the unusually large and diverse C. elegans insulin gene family. Genes Dev. 2001, 15, 672–686. [Google Scholar] [CrossRef] [Green Version]
- Cornils, A.; Gloeck, M.; Chen, Z.; Zhang, Y.; Alcedo, J. Specific insulin-like peptides encode sensory information to regulate distinct developmental processes. Development 2011, 138, 1183–1193. [Google Scholar] [CrossRef] [Green Version]
- Kodama, E.; Kuhara, A.; Mohri-Shiomi, A.; Kimura, K.D.; Okumura, M.; Tomioka, M.; Iino, Y.; Mori, I. Insulin-like signaling and the neural circuit for integrative behavior in C. elegans. Genes Dev. 2006, 20, 2955–2960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes De Abreu, D.A.; Caballero, A.; Fardel, P.; Stroustrup, N.; Chen, Z.; Lee, K.; Keyes, W.D.; Nash, Z.M.; López-Moyado, I.F.; Vaggi, F.; et al. An insulin-to-insulin regulatory network orchestrates phenotypic specificity in development and physiology. PLoS Genet. 2014, 10, e1004225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, S.; Chiu, H.; Boudreau, J.; Papanicolaou, T.; Bendena, W.; Chin-Sang, I. A functional study of all 40 Caenorhabditis elegans insulin-like peptides. J. Biol. Chem. 2018, 293, 16912–16922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brogiolo, W.; Stocker, H.; Ikeya, T.; Rintelen, F.; Fernandez, R.; Hafen, E. An evolutionarily conserved function of the Drosophila insulin receptor and insulin-like peptides in growth control. Curr. Biol. 2001, 11, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Garelli, A.; Gontijo, A.M.; Miguela, V.; Caparros, E.; Dominguez, M. Imaginal discs secrete insulin-like peptide 8 to mediate plasticity of growth and maturation. Science 2012, 336, 579–582. [Google Scholar] [CrossRef]
- Bedarkar, S.; Turnell, W.G.; Blundell, T.L.; Schwabe, C. Relaxin has conformational homology with insulin. Nature 1977, 270, 449–451. [Google Scholar] [CrossRef]
- Rinderknecht, E.; Humbel, R.E. The amino acid sequence of human insulin-like growth factor i and its structural homology with proinsulin. J. Biol. Chem. 1978, 253, 2769–2776. [Google Scholar]
- Bell, G.I.; Pictet, R.L.; Rutter, W.J.; Cordell, B.; Tischer, E.; Goodman, H.M. Sequence of the human insulin gene. Nature 1980, 284, 26–32. [Google Scholar] [CrossRef]
- Büllesbach, E.E.; Schwabe, C. A novel leydig cell cdna-derived protein is a relaxin-like factor. J. Biol. Chem. 1995, 270, 16011–16015. [Google Scholar] [CrossRef] [Green Version]
- Chassin, D.; Laurent, A.; Janneau, J.-L.; Berger, R.; Bellet, D. Cloning of a new member of the insulin gene superfamily (insl4) expressed in human placenta. Genomics 1995, 29, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Clayton, P.E.; Banerjee, I.; Murray, P.G.; Renehan, A.G. Growth hormone, the insulin-like growth factor axis, insulin and cancer risk. Nat. Rev. Endocrinol. 2011, 7, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Weinkove, D.; Halstead, J.R.; Gems, D.; Divecha, N. Long-term starvation and ageing induce age-1/pi 3-kinase-dependent translocation of daf-16/foxo to the cytoplasm. BMC Biol. 2006, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taniguchi, C.M.; Emanuelli, B.; Kahn, C.R. Critical nodes in signalling pathways: Insights into insulin action. Nat. Rev. Mol. Cell Biol. 2006, 7, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Wolkow, C.A.; Muñoz, M.J.; Riddle, D.L.; Ruvkun, G. Insulin receptor substrate and p55 orthologous adaptor proteins function in the Caenorhabditis elegans daf-2/insulin-like signaling pathway. J. Biol. Chem. 2002, 277, 49591–49597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paradis, S.; Ailion, M.; Toker, A.; Thomas, J.H.; Ruvkun, G. A pdk1 homolog is necessary and sufficient to transduce age-1 pi3 kinase signals that regulate diapause in Caenorhabditis elegans. Genes Dev. 1999, 13, 1438–1452. [Google Scholar] [CrossRef]
- Paradis, S.; Ruvkun, G. Caenorhabditis elegans akt/pkb transduces insulin receptor-like signals from age-1 pi3 kinase to the daf-16 transcription factor. Genes Dev. 1998, 12, 2488–2498. [Google Scholar] [CrossRef] [Green Version]
- Hertweck, M.; Göbel, C.; Baumeister, R.C. Elegans sgk-1 is the critical component in the akt/pkb kinase complex to control stress response and life span. Dev. Cell 2004, 6, 577–588. [Google Scholar] [CrossRef] [Green Version]
- Brunet, A.; Bonni, A.; Zigmond, M.J.; Lin, M.Z.; Juo, P.; Hu, L.S.; Anderson, M.J.; Arden, K.C.; Blenis, J.; Greenberg, M.E. Akt promotes cell survival by phosphorylating and inhibiting a forkhead transcription factor. Cell 1999, 96, 857–868. [Google Scholar] [CrossRef] [Green Version]
- Brunet, A.; Park, J.; Tran, H.; Hu, L.S.; Hemmings, B.A.; Greenberg, M.E. Protein kinase sgk mediates survival signals by phosphorylating the forkhead transcription factor fkhrl1 (foxo3a). Mol. Cell Biol. 2001, 21, 952–965. [Google Scholar] [CrossRef] [Green Version]
- Chen, A.T.-Y.; Guo, C.; Dumas, K.J.; Ashrafi, K.; Hu, P.J. Effects of Caenorhabditis elegans sgk-1 mutations on lifespan, stress resistance, and daf-16/foxo regulation. Aging Cell 2013, 12, 932–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berdichevsky, A.; Viswanathan, M.; Horvitz, H.R.; Guarente, L.C. Elegans sir-2.1 interacts with 14-3-3 proteins to activate daf-16 and extend life span. Cell 2006, 125, 1165–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Tewari, M.; Vidal, M.; Sylvia Lee, S. The 14-3-3 protein ftt-2 regulates daf-16 in Caenorhabditis elegans. Dev. Biol. 2007, 301, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Kim, S.K. Global analysis of dauer gene expression in Caenorhabditis elegans. Development 2003, 130, 1621–1634. [Google Scholar] [CrossRef] [Green Version]
- Evans, E.A.; Kawli, T.; Tan, M.-W. Pseudomonas aeruginosa suppresses host immunity by activating the daf-2 insulin-like signaling pathway in Caenorhabditis elegans. PLoS Pathog. 2008, 4, e1000175. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.W.; Mukhopadhyay, A.; Svrzikapa, N.; Jiang, F.; Davis, R.J.; Tissenbaum, H.A. Jnk regulates lifespan in Caenorhabditis elegans by modulating nuclear translocation of forkhead transcription factor/daf-16. Proc. Natl. Acad. Sci. USA 2005, 102, 4494–4499. [Google Scholar] [CrossRef] [Green Version]
- Lehtinen, M.K.; Yuan, Z.; Boag, P.R.; Yang, Y.; Villén, J.; Becker, E.B.E.; DiBacco, S.; De La Iglesia, N.; Gygi, S.; Blackwell, T.K.; et al. A conserved mst-foxo signaling pathway mediates oxidative-stress responses and extends life span. Cell 2006, 125, 987–1001. [Google Scholar] [CrossRef] [Green Version]
- Greer, E.L.; Dowlatshahi, D.; Banko, M.R.; Villen, J.; Hoang, K.; Blanchard, D.; Gygi, S.P.; Brunet, A. An ampk-foxo pathway mediates longevity induced by a novel method of dietary restriction in C. elegans. Curr. Biol. 2007, 17, 1646–1656. [Google Scholar] [CrossRef] [Green Version]
- Tullet, J.M.A.; Araiz, C.; Sanders, M.J.; Au, C.; Benedetto, A.; Papatheodorou, I.; Clark, E.; Schmeisser, K.; Jones, D.; Schuster, E.F.; et al. Daf-16/foxo directly regulates an atypical amp-activated protein kinase gamma isoform to mediate the effects of insulin/igf-1 signaling on aging in Caenorhabditis elegans. PLoS Genet. 2014, 10, e1004109. [Google Scholar] [CrossRef]
- Robida-Stubbs, S.; Glover-Cutter, K.; Lamming, D.W.; Mizunuma, M.; Narasimhan, S.D.; Neumann-Haefelin, E.; Sabatini, D.M.; Blackwell, T.K. Tor signaling and rapamycin influence longevity by regulating skn-1/nrf and daf-16/foxo. Cell Metab. 2012, 15, 713–724. [Google Scholar] [CrossRef] [Green Version]
- Honjoh, S.; Yamamoto, T.; Uno, M.; Nishida, E. Signalling through rheb-1 mediates intermittent fasting-induced longevity in C. elegans. Nature 2009, 457, 726–730. [Google Scholar] [CrossRef] [PubMed]
- Berman, J.R.; Kenyon, C. Germ-cell loss extends C. elegans life span through regulation of daf-16 by kri-1 and lipophilic-hormone signaling. Cell 2006, 124, 1055–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulias, K.; Horvitz, H.R. The C. elegans microrna mir-71 acts in neurons to promote germline-mediated longevity through regulation of daf-16/foxo. Cell Metab. 2012, 15, 439–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, C.T. The search for daf-16/foxo transcriptional targets: Approaches and discoveries. Exp. Gerontol. 2006, 41, 910–921. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.S.; Kennedy, S.; Tolonen, A.C.; Ruvkun, G. Daf-16 target genes that control C. elegans life-span and metabolism. Science 2003, 300, 644–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McElwee, J.; Bubb, K.; Thomas, J.H. Transcriptional outputs of the Caenorhabditis elegans forkhead protein daf-16. Aging Cell 2003, 2, 111–121. [Google Scholar] [CrossRef] [Green Version]
- Halaschek-Wiener, J.; Khattra, J.S.; McKay, S.; Pouzyrev, A.; Stott, J.M.; Yang, G.S.; Holt, R.A.; Jones, S.J.M.; Marra, M.A.; Brooks-Wilson, A.R.; et al. Analysis of long-lived C. elegans daf-2 mutants using serial analysis of gene expression. Genome Res. 2005, 15, 603–615. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.W.; Mukhopadhyay, A.; Dixit, B.L.; Raha, T.; Green, M.R.; Tissenbaum, H.A. Identification of direct daf-16 targets controlling longevity, metabolism and diapause by chromatin immunoprecipitation. Nat. Genet. 2006, 38, 251–257. [Google Scholar]
- Kumar, N.; Jain, V.; Singh, A.; Jagtap, U.; Verma, S.; Mukhopadhyay, A. Genome-wide endogenous daf-16/foxo recruitment dynamics during lowered insulin signalling in C. elegans. Oncotarget 2015, 6, 41418–41433. [Google Scholar] [CrossRef] [Green Version]
- Webb, A.E.; Kundaje, A.; Brunet, A. Characterization of the direct targets of foxo transcription factors throughout evolution. Aging Cell 2016, 15, 673–685. [Google Scholar] [CrossRef]
- McElwee, J.J.; Schuster, E.; Blanc, E.; Thornton, J.; Gems, D. Diapause-associated metabolic traits reiterated in long-lived daf-2 mutants in the nematode Caenorhabditis elegans. Mech. Ageing Dev. 2006, 127, 458–472. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.-Q.; Venable, J.D.; Au, N.; Xu, T.; Park, S.K.; Cociorva, D.; Johnson, J.R.; Dillin, A.; Yates, J.R. Quantitative mass spectrometry identifies insulin signaling targets in C. elegans. Science 2007, 317, 660–663. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.M.; Staffa, K.; Perally, S.; LaCourse, E.J.; Brophy, P.M.; Hamilton, J.V. Proteomic analyses of Caenorhabditis elegans dauer larvae and long-lived daf-2 mutants implicates a shared detoxification system in longevity assurance. J. Proteome Res. 2010, 9, 2871–2881. [Google Scholar] [CrossRef] [PubMed]
- Stout, G.J.; Stigter, E.C.A.; Essers, P.B.; Mulder, K.W.; Kolkman, A.; Snijders, D.S.; Van Den Broek, N.J.F.; Betist, M.C.; Korswagen, H.C.; MacInnes, A.W.; et al. Insulin/igf-1-mediated longevity is marked by reduced protein metabolism. Mol. Syst. Biol. 2013, 9, 679. [Google Scholar] [CrossRef]
- Depuydt, G.; Xie, F.; Petyuk, V.A.; Smolders, A.; Brewer, H.M.; Camp, D.G., 2nd; Smith, R.D.; Braeckman, B.P. Lc-ms proteomics analysis of the insulin/igf-1-deficient Caenorhabditis elegans daf-2(e1370) mutant reveals extensive restructuring of intermediary metabolism. J. Proteome Res. 2014, 13, 1938–1956. [Google Scholar] [CrossRef]
- Walther, D.M.; Kasturi, P.; Zheng, M.; Pinkert, S.; Vecchi, G.; Ciryam, P.; Morimoto, R.I.; Dobson, C.M.; Vendruscolo, M.; Mann, M.; et al. Widespread proteome remodeling and aggregation in aging C. elegans. Cell 2015, 161, 919–932. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, S.; Bundy, J.G.; Davies, S.K.; Viney, J.M.; Swire, J.S.; Leroi, A.M. A metabolic signature of long life in Caenorhabditis elegans. BMC Biol. 2010, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Gao, A.W.; Smith, R.L.; Van Weeghel, M.; Kamble, R.; Janssens, G.E.; Houtkooper, R.H. Identification of key pathways and metabolic fingerprints of longevity in C. elegans. Exp. Gerontol. 2018, 113, 128–140. [Google Scholar] [CrossRef]
- Shaw, W.M.; Luo, S.; Landis, J.; Ashraf, J.; Murphy, C.T. The C. elegans tgf-beta dauer pathway regulates longevity via insulin signaling. Curr. Biol. 2007, 17, 1635–1645. [Google Scholar] [CrossRef] [Green Version]
- Gems, D.; Sutton, A.J.; Sundermeyer, M.L.; Albert, P.S.; King, K.V.; Edgley, M.L.; Larsen, P.L.; Riddle, D.L. Two pleiotropic classes of daf-2 mutation affect larval arrest, adult behavior, reproduction and longevity in Caenorhabditis elegans. Genetics 1998, 150, 129–155. [Google Scholar]
- Ewald, C.Y.; Landis, J.N.; Porter Abate, J.; Murphy, C.T.; Blackwell, T.K. Dauer-independent insulin/igf-1-signalling implicates collagen remodelling in longevity. Nature 2015, 519, 97–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewald, C.Y.; Castillo-Quan, J.I.; Blackwell, T.K. Untangling longevity, dauer, and healthspan in Caenorhabditis elegans insulin/igf-1-signalling. Gerontology 2018, 64, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Lithgow, G.J.; White, T.M.; Melov, S.; Johnson, T.E. Thermotolerance and extended life-span conferred by single-gene mutations and induced by thermal stress. Proc. Natl. Acad. Sci. USA 1995, 92, 7540–7544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanfleteren, J.R. Oxidative stress and ageing in Caenorhabditis elegans. Biochem. J. 1993, 292, 605–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, Y.; Honda, S. The daf-2 gene network for longevity regulates oxidative stress resistance and mn-superoxide dismutase gene expression in Caenorhabditis elegans. FASEB J. 1999, 13, 1385–1393. [Google Scholar] [CrossRef] [Green Version]
- Murakami, S.; Johnson, T.E. A genetic pathway conferring life extension and resistance to uv stress in Caenorhabditis elegans. Genetics 1996, 143, 1207–1218. [Google Scholar]
- Lamitina, S.T.; Strange, K. Transcriptional targets of daf-16 insulin signaling pathway protect C. elegans from extreme hypertonic stress. Am. J. Physiol. Cell Physiol. 2005, 288, C467–C474. [Google Scholar] [CrossRef]
- Scott, B.A.; Avidan, M.S.; Crowder, C.M. Regulation of hypoxic death in C. elegans by the insulin/igf receptor homolog daf-2. Science 2002, 296, 2388–2391. [Google Scholar] [CrossRef]
- Mabon, M.E.; Scott, B.A.; Crowder, C.M. Divergent mechanisms controlling hypoxic sensitivity and lifespan by the daf-2/insulin/igf-receptor pathway. PLoS ONE 2009, 4, e7937. [Google Scholar] [CrossRef]
- Barsyte, D.; Lovejoy, D.A.; Lithgow, G.J. Longevity and heavy metal resistance in daf-2 and age-1 long-lived mutants of Caenorhabditis elegans. FASEB J. 2001, 15, 627–634. [Google Scholar] [CrossRef] [Green Version]
- Garsin, D.A.; Villanueva, J.M.; Begun, J.; Kim, D.H.; Sifri, C.D.; Calderwood, S.B.; Ruvkun, G.; Ausubel, F.M. Long-lived C. elegans daf-2 mutants are resistant to bacterial pathogens. Science 2003, 300, 1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, E.A.; Chen, W.C.; Tan, M.-W. The daf-2 insulin-like signaling pathway independently regulates aging and immunity in C. elegans. Aging Cell 2008, 7, 879–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, W.-C.; Ching, T.-T.; Lee, H.C.; Mousigian, C.; Hsu, A.-L. Hsf-1 regulators ddl-1/2 link insulin-like signaling to heat-shock responses and modulation of longevity. Cell 2012, 148, 322–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, V.; Aballay, A. Heat-shock transcription factor (hsf)-1 pathway required for Caenorhabditis elegans immunity. Proc. Natl. Acad. Sci. USA 2006, 103, 13092–13097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, J.H.; Blackwell, T.K. Skn-1 links C. elegans mesendodermal specification to a conserved oxidative stress response. Genes Dev. 2003, 17, 1882–1893. [Google Scholar] [CrossRef] [Green Version]
- Tullet, J.M.A.; Hertweck, M.; An, J.H.; Baker, J.; Hwang, J.Y.; Liu, S.; Oliveira, R.P.; Baumeister, R.; Blackwell, T.K. Direct inhibition of the longevity-promoting factor skn-1 by insulin-like signaling in C. elegans. Cell 2008, 132, 1025–1038. [Google Scholar] [CrossRef] [Green Version]
- Papp, D.; Csermely, P.; Sőti, C. A role for skn-1/nrf in pathogen resistance and immunosenescence in Caenorhabditis elegans. PLoS Pathog. 2012, 8, e1002673. [Google Scholar] [CrossRef] [Green Version]
- Wolff, S.; Ma, H.; Burch, D.; Maciel, G.A.; Hunter, T.; Dillin, A. Smk-1, an essential regulator of daf-16-mediated longevity. Cell 2006, 124, 1039–1053. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Feinbaum, R.; Alloing, G.; Emerson, F.E.; Garsin, D.A.; Inoue, H.; Tanaka-Hino, M.; Hisamoto, N.; Matsumoto, K.; Tan, M.-W.; et al. A conserved p38 map kinase pathway in Caenorhabditis elegans innate immunity. Science 2002, 297, 623–626. [Google Scholar] [CrossRef] [Green Version]
- Larsen, P.L. Aging and resistance to oxidative damage in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 1993, 90, 8905–8909. [Google Scholar] [CrossRef] [Green Version]
- Houthoofd, K.; Fidalgo, M.A.; Hoogewijs, D.; Braeckman, B.P.; Lenaerts, I.; Brys, K.; Matthijssens, F.; De Vreese, A.; Van Eygen, S.; Muñoz, M.J.; et al. Metabolism, physiology and stress defense in three aging ins/igf-1 mutants of the nematode Caenorhabditis elegans. Aging Cell 2005, 4, 87–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, Y.; Tanaka, M.; Honda, S. Modulation of longevity and diapause by redox regulation mechanisms under the insulin-like signaling control in Caenorhabditis elegans. Exp. Gerontol. 2008, 43, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Doonan, R.; McElwee, J.J.; Matthijssens, F.; Walker, G.A.; Houthoofd, K.; Back, P.; Matscheski, A.; Vanfleteren, J.R.; Gems, D. Against the oxidative damage theory of aging: Superoxide dismutases protect against oxidative stress but have little or no effect on life span in Caenorhabditis elegans. Genes Dev. 2008, 22, 3236–3241. [Google Scholar] [CrossRef] [Green Version]
- Van Raamsdonk, J.M.; Hekimi, S. Superoxide dismutase is dispensable for normal animal lifespan. Proc. Natl. Acad. Sci. USA 2012, 109, 5785–5790. [Google Scholar] [CrossRef] [Green Version]
- Dues, D.J.; Andrews, E.K.; Senchuk, M.M.; Van Raamsdonk, J.M. Resistance to stress can be experimentally dissociated from longevity. J. Gerontol. A Biol. 2018, 74, 1206–1214. [Google Scholar] [CrossRef] [Green Version]
- Erkut, C.; Vasilj, A.; Boland, S.; Habermann, B.; Shevchenko, A.; Kurzchalia, T.V. Molecular strategies of the Caenorhabditis elegans dauer larva to survive extreme desiccation. PLoS ONE 2013, 8, e82473. [Google Scholar] [CrossRef] [Green Version]
- Lant, B.; Storey, K.B. An overview of stress response and hypometabolic strategies in Caenorhabditis elegans: Conserved and contrasting signals with the mammalian system. Int. J. Biol. Sci. 2010, 6, 9–50. [Google Scholar] [CrossRef] [Green Version]
- Burnell, A.M.; Houthoofd, K.; O’Hanlon, K.; Vanfleteren, J.R. Alternate metabolism during the dauer stage of the nematode Caenorhabditis elegans. Exp. Gerontol. 2005, 40, 850–856. [Google Scholar] [CrossRef] [Green Version]
- Xie, M.; Roy, R. Amp-activated kinase regulates lipid droplet localization and stability of adipose triglyceride lipase in C. elegans dauer larvae. PLoS ONE 2015, 10, e0130480. [Google Scholar] [CrossRef] [Green Version]
- Narbonne, P.; Roy, R. Caenorhabditis elegans dauers need lkb1/ampk to ration lipid reserves and ensure long-term survival. Nature 2009, 457, 210–214. [Google Scholar] [CrossRef]
- Yen, K.; Le, T.T.; Bansal, A.; Narasimhan, S.D.; Cheng, J.-X.; Tissenbaum, H.A. A comparative study of fat storage quantitation in nematode Caenorhabditis elegans using label and label-free methods. PLoS ONE 2010, 5, e12810. [Google Scholar] [CrossRef] [PubMed]
- Frazier, H.N., III; Roth, M.B. Adaptive sugar provisioning controls survival of C. elegans embryos in adverse environments. Curr. Biol. 2009, 19, 859–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, C.L.; Van Gilst, M.R. A 13C isotope labeling strategy reveals the influence of insulin signaling on lipogenesis in C. elegans. Cell Metab. 2008, 8, 266–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shmookler Reis, R.J.; Xu, L.; Lee, H.; Chae, M.; Thaden, J.J.; Bharill, P.; Tazearslan, C.; Siegel, E.; Alla, R.; Zimniak, P.; et al. Modulation of lipid biosynthesis contributes to stress resistance and longevity of C. elegans mutants. Aging 2011, 3, 125–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, C.; Krumsiek, J.; Lehrbach, N.J.; Murfitt, S.A.; Miska, E.A.; Griffin, J.L. A study of Caenorhabditis elegans daf-2 mutants by metabolomics and differential correlation networks. Mol. Biosyst. 2013, 9, 1632–1642. [Google Scholar] [CrossRef] [PubMed]
- Possik, E.; Ajisebutu, A.; Manteghi, S.; Gingras, M.-C.; Vijayaraghavan, T.; Flamand, M.; Coull, B.; Schmeisser, K.; Duchaine, T.; Van Steensel, M.; et al. Flcn and ampk confer resistance to hyperosmotic stress via remodeling of glycogen stores. PLoS Genet. 2015, 11, e1005520. [Google Scholar] [CrossRef] [Green Version]
- LaMacchia, J.C.; Frazier, H.N., 3rd; Roth, M.B. Glycogen fuels survival during hyposmotic-anoxic stress in Caenorhabditis elegans. Genetics 2015, 201, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Gusarov, I.; Pani, B.; Gautier, L.; Smolentseva, O.; Eremina, S.; Shamovsky, I.; Katkova-Zhukotskaya, O.; Mironov, A.; Nudler, E. Glycogen controls Caenorhabditis elegans lifespan and resistance to oxidative stress. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef]
- Seo, Y.; Kingsley, S.; Walker, G.; Mondoux, M.A.; Tissenbaum, H.A. Metabolic shift from glycogen to trehalose promotes lifespan and healthspan in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2018, 115, E2791. [Google Scholar] [CrossRef] [Green Version]
- Clegg, J.S.; Evans, D.R. The physiology of blood trehalose and its function during flight in the blowfly. J. Exp. Biol. 1961, 38, 771–792. [Google Scholar]
- Erkut, C.; Penkov, S.; Khesbak, H.; Vorkel, D.; Verbavatz, J.-M.; Fahmy, K.; Kurzchalia, T.V. Trehalose renders the dauer larva of Caenorhabditis elegans resistant to extreme desiccation. Curr. Biol. 2011, 21, 1331–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapia, H.; Young, L.; Fox, D.; Bertozzi, C.R.; Koshland, D. Increasing intracellular trehalose is sufficient to confer desiccation tolerance to Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2015, 112, 6122–6127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, B.; Haltiwanger, R.S.; Futcher, B. Synergy between trehalose and hsp104 for thermotolerance in Saccharomyces cerevisiae. Genetics 1996, 144, 923–933. [Google Scholar] [PubMed]
- Page-Sharp, M.; Behm, C.A.; Smith, G.D. Involvement of the compatible solutes trehalose and sucrose in the response to salt stress of a cyanobacterial scytonema species isolated from desert soils. Biochim. Biophys. Acta 1999, 1472, 519–528. [Google Scholar] [CrossRef]
- Pellerone, F.I.; Archer, S.K.; Behm, C.A.; Grant, W.N.; Lacey, M.J.; Somerville, A.C. Trehalose metabolism genes in Caenorhabditis elegans and filarial nematodes. Int. J. Parasitol. 2003, 33, 1195–1206. [Google Scholar] [CrossRef]
- Goyal, K.; Browne, J.A.; Burnell, A.M.; Tunnacliffe, A. Dehydration-induced tps gene transcripts from an anhydrobiotic nematode contain novel spliced leaders and encode atypical gt-20 family proteins. Biochimie 2005, 87, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Honda, Y.; Tanaka, M.; Honda, S. Trehalose extends longevity in the nematode Caenorhabditis elegans. Aging Cell 2010, 9, 558–569. [Google Scholar] [CrossRef]
- Erkut, C.; Gade, V.R.; Laxman, S.; Kurzchalia, T.V. The glyoxylate shunt is essential for desiccation tolerance in C. elegans and budding yeast. eLife 2016, 5, e13614. [Google Scholar] [CrossRef]
- Liu, F.; Thatcher, J.D.; Barral, J.M.; Epstein, H.F. Bifunctional glyoxylate cycle protein of Caenorhabditis elegans: A developmentally regulated protein of intestine and muscle. Dev. Biol. 1995, 169, 399–414. [Google Scholar] [CrossRef] [Green Version]
- Braeckman, B.P.; Houthoofd, K.; Vanfleteren, J.R. Intermediary Metabolism. Available online: http://www.wormbook.org/chapters/www_intermetabolism/intermetabolism.html (accessed on 31 December 2019).
- Shen, E.-Z.; Song, C.-Q.; Lin, Y.; Zhang, W.-H.; Su, P.-F.; Liu, W.-Y.; Zhang, P.; Xu, J.; Lin, N.; Zhan, C.; et al. Mitoflash frequency in early adulthood predicts lifespan in Caenorhabditis elegans. Nature 2014, 508, 128–132. [Google Scholar] [CrossRef]
- Holt, S.J.; Riddle, D.L. Sage surveys C. elegans carbohydrate metabolism: Evidence for an anaerobic shift in the long-lived dauer larva. Mech. Ageing Dev. 2003, 124, 779–800. [Google Scholar] [CrossRef]
- Yilmaz, L.S.; Walhout, A.J. A Caenorhabditis elegans genome-scale metabolic network model. Cell Syst. 2016, 2, 297–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakrabortee, S.; Boschetti, C.; Walton, L.J.; Sarkar, S.; Rubinsztein, D.C.; Tunnacliffe, A. Hydrophilic protein associated with desiccation tolerance exhibits broad protein stabilization function. Proc. Natl. Acad. Sci. USA 2007, 104, 18073–18078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, S.; Davies, J.E.; Huang, Z.; Tunnacliffe, A.; Rubinsztein, D.C. Trehalose, a novel mtor-independent autophagy enhancer, accelerates the clearance of mutant huntingtin and α-synuclein. J. Biol. Chem. 2007, 282, 5641–5652. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.M.; Campbell, L.T.; Lodge, J.K. Cryptococcus neoformans, a fungus under stress. Curr. Opin. Microbiol. 2007, 10, 320–325. [Google Scholar] [CrossRef] [Green Version]
- Challabathula, D.; Bartels, D. Desiccation tolerance in resurrection plants: New insights from transcriptome, proteome and metabolome analysis. Front. Plant. Sci. 2013, 4, 482. [Google Scholar]
- Alpert, P. Constraints of tolerance: Why are desiccation-tolerant organisms so small or rare? J. Exp. Biol. 2006, 209, 1575–1584. [Google Scholar] [CrossRef] [Green Version]
- Martins, R.; Lithgow, G.J.; Link, W. Long live foxo: Unraveling the role of foxo proteins in aging and longevity. Aging Cell 2016, 15, 196–207. [Google Scholar] [CrossRef]
- Stefanetti, R.J.; Voisin, S.; Russell, A.; Lamon, S. Recent advances in understanding the role of foxo3. F1000Res 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Morris, B.J.; Willcox, D.C.; Donlon, T.A.; Willcox, B.J. Foxo3: A major gene for human longevity—A mini-review. Gerontology 2015, 61, 515–525. [Google Scholar] [CrossRef] [Green Version]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zečić, A.; Braeckman, B.P. DAF-16/FoxO in Caenorhabditis elegans and Its Role in Metabolic Remodeling. Cells 2020, 9, 109. https://doi.org/10.3390/cells9010109
Zečić A, Braeckman BP. DAF-16/FoxO in Caenorhabditis elegans and Its Role in Metabolic Remodeling. Cells. 2020; 9(1):109. https://doi.org/10.3390/cells9010109
Chicago/Turabian StyleZečić, Aleksandra, and Bart P. Braeckman. 2020. "DAF-16/FoxO in Caenorhabditis elegans and Its Role in Metabolic Remodeling" Cells 9, no. 1: 109. https://doi.org/10.3390/cells9010109
APA StyleZečić, A., & Braeckman, B. P. (2020). DAF-16/FoxO in Caenorhabditis elegans and Its Role in Metabolic Remodeling. Cells, 9(1), 109. https://doi.org/10.3390/cells9010109