Pathogenic Carboxyl Ester Lipase (CEL) Variants Interact with the Normal CEL Protein in Pancreatic Cells

 , , , , and
, , , , and 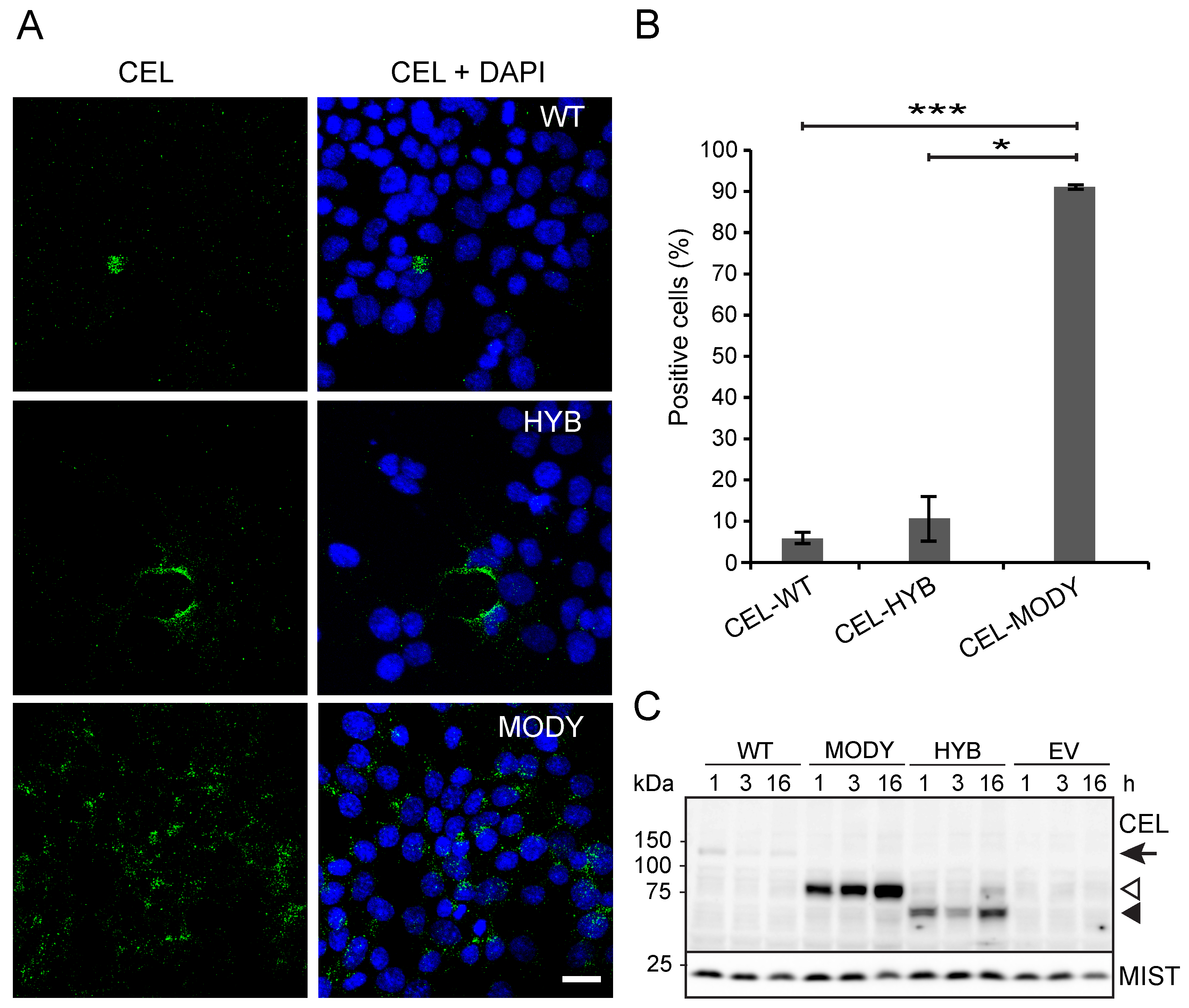{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmids
2.2. Antibodies and Reagents
2.3. Cell Culture, Transfection, and Drug Treatment
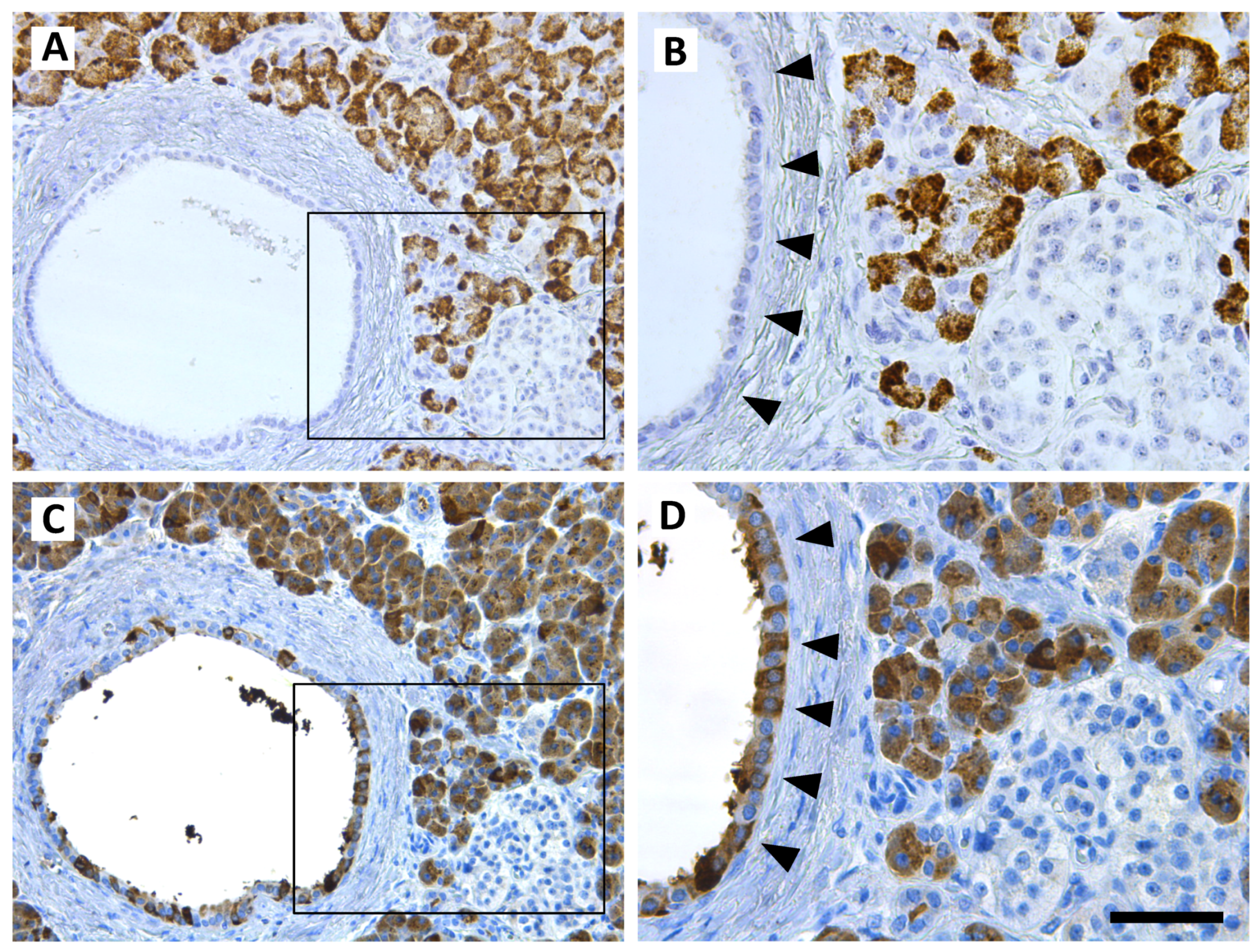
2.4. Human Pancreatic Cells and Tissue
2.5. Endocytosis
2.6. SDS-PAGE and Western Blotting
2.7. Immunofluorescent Staining and Confocal Microscopy
2.8. Apoptosis Assay
2.9. Cell Viability
2.10. Immunohistochemistry and In Situ Hybridization
2.11. Statistics
3. Results
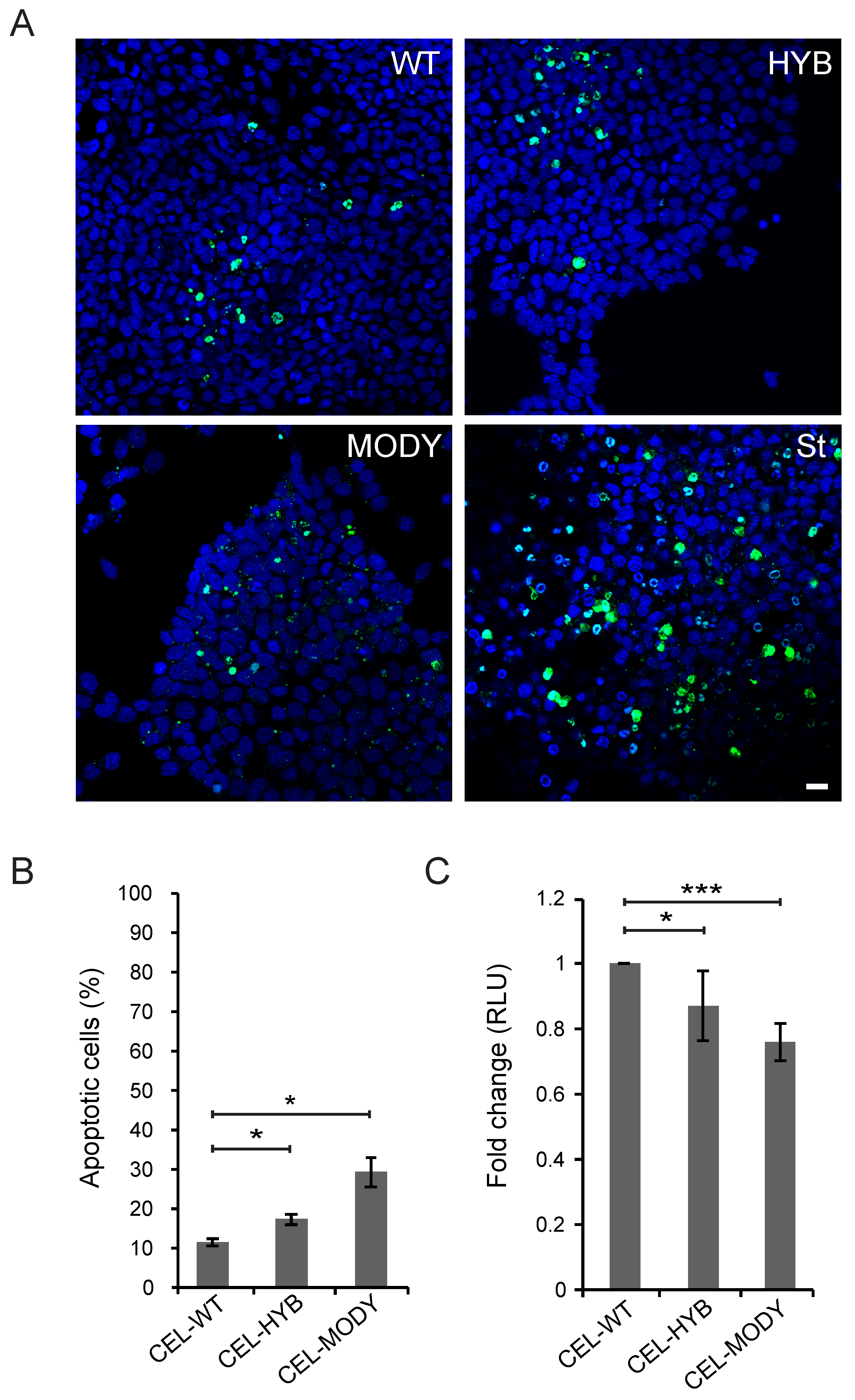
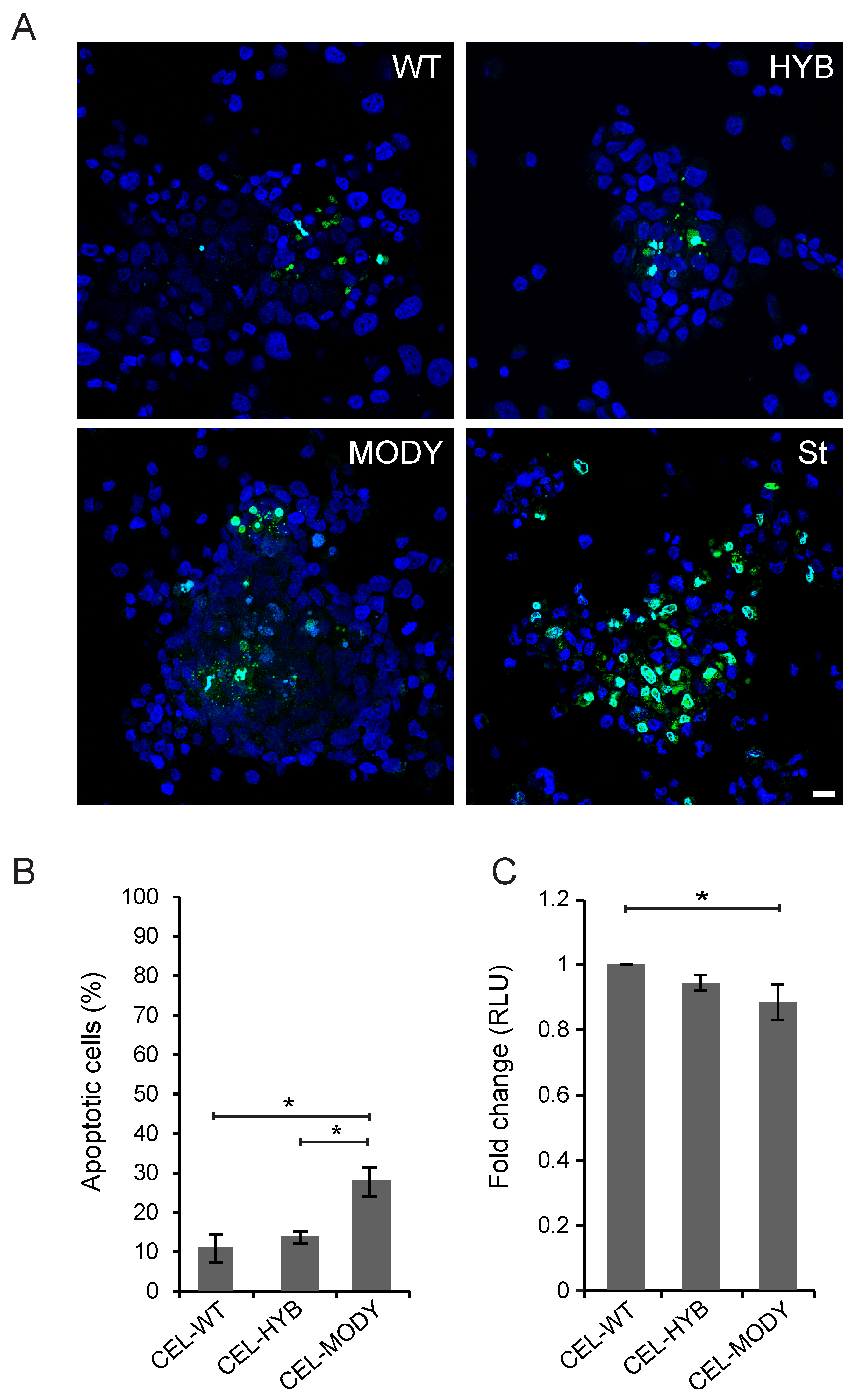
3.1. CEL-HYB Endocytosis Affects Apoptosis and Viability of Pancreatic Acinar Cells
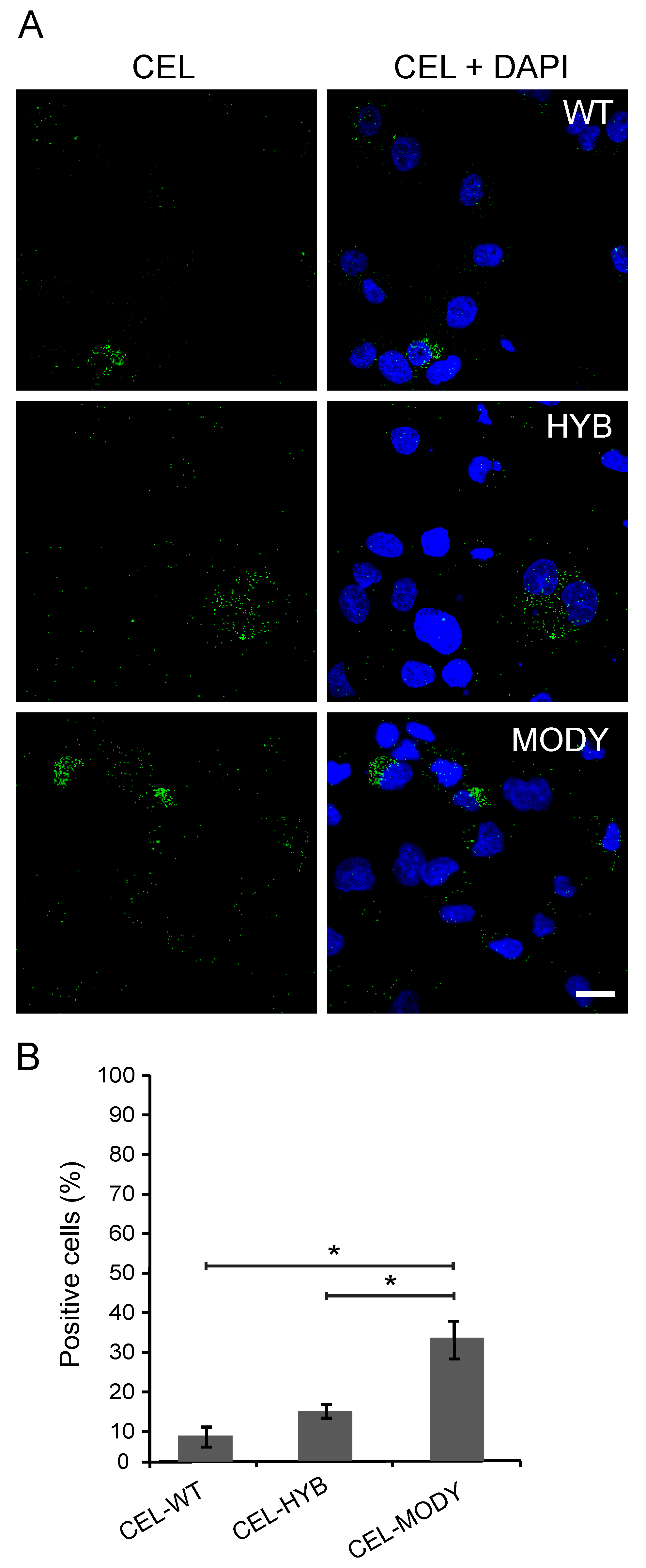
3.2. CEL Protein Variants are Endocytosed by Pancreatic Ductal Cells
3.3. CEL-MODY Can Induce Apoptosis and Reduce the Viability of Pancreatic Ductal Cells
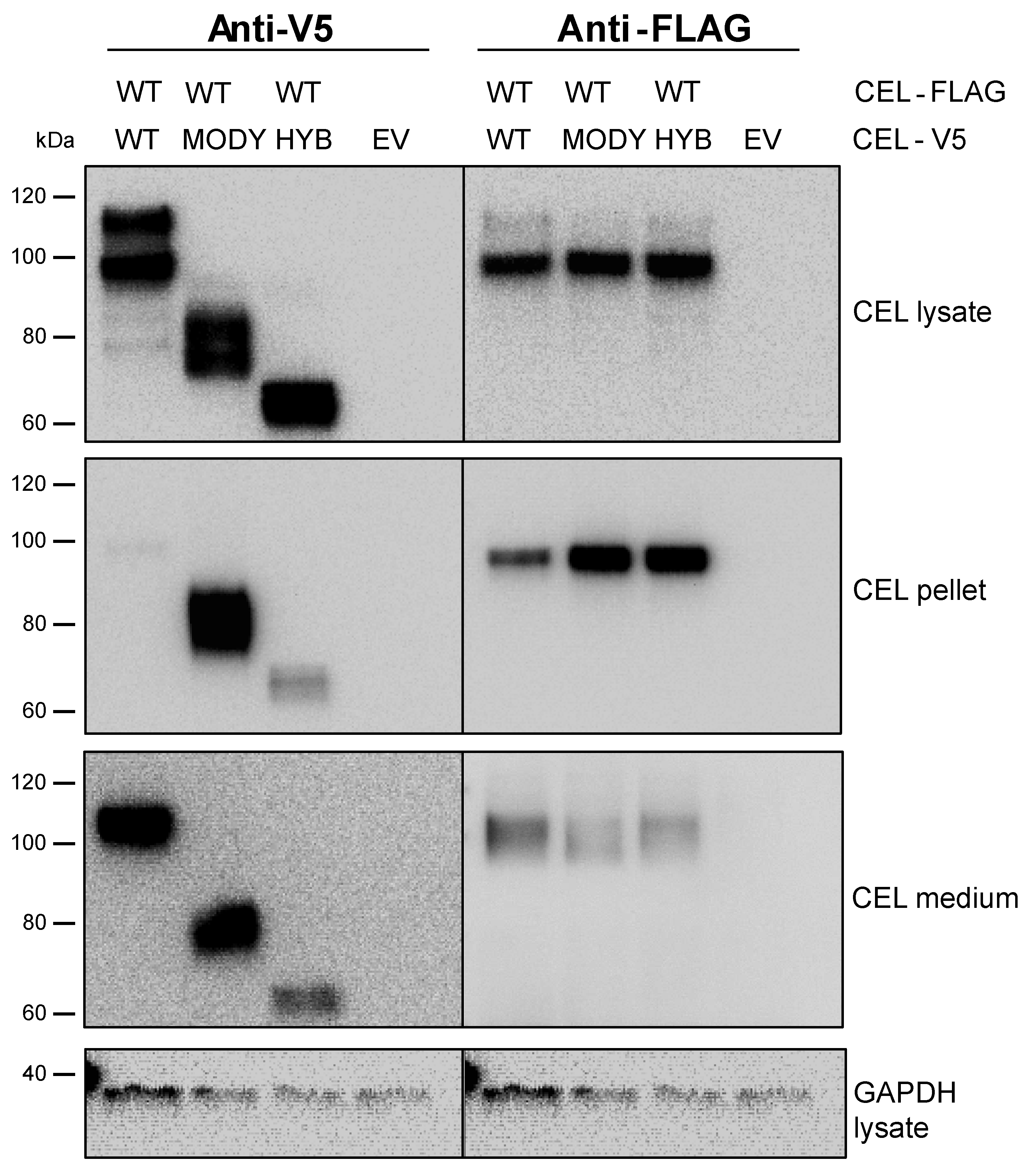
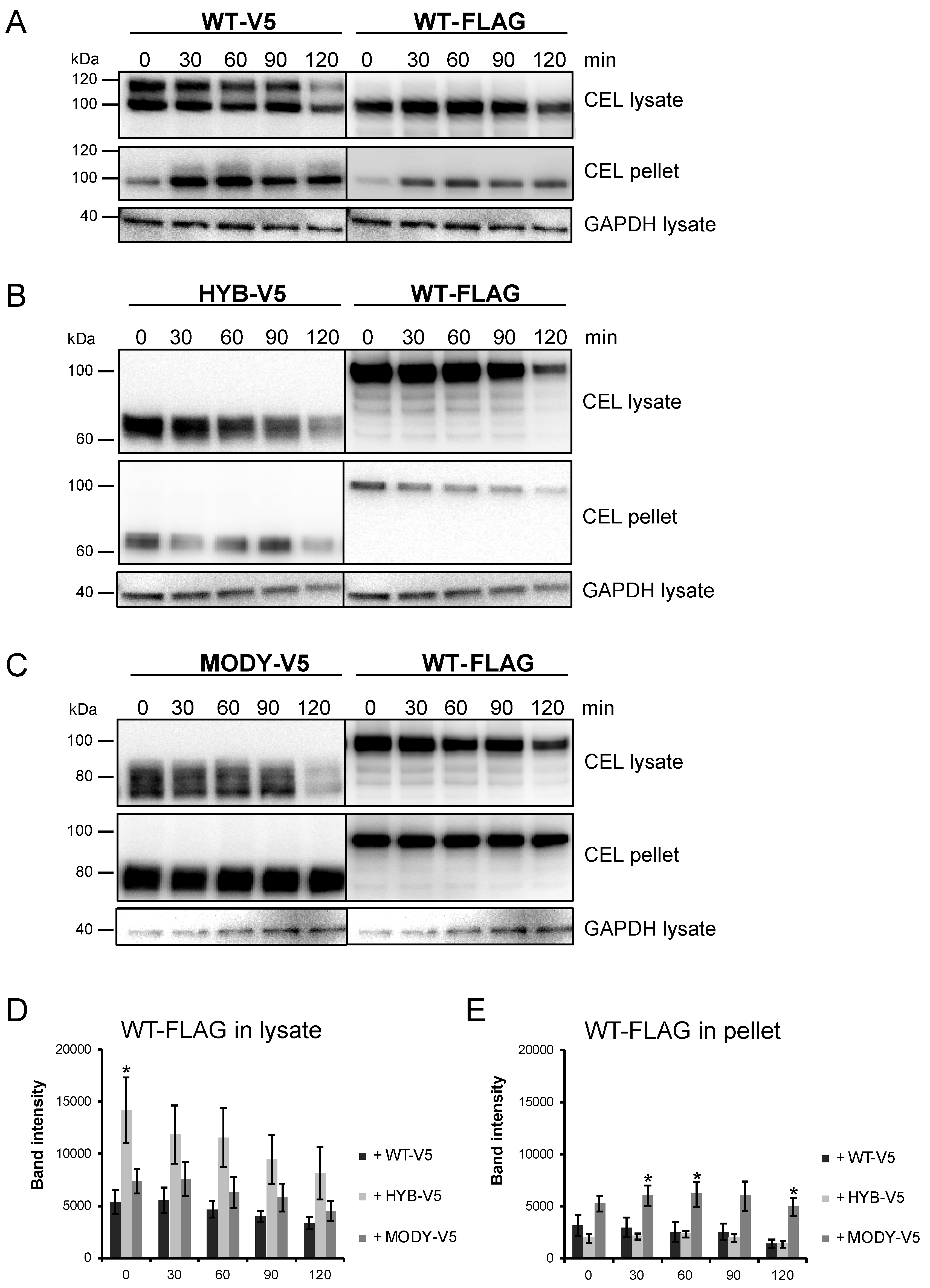
3.4. The Secretion of CEL-WT is Affected by Pathogenic CEL Variants
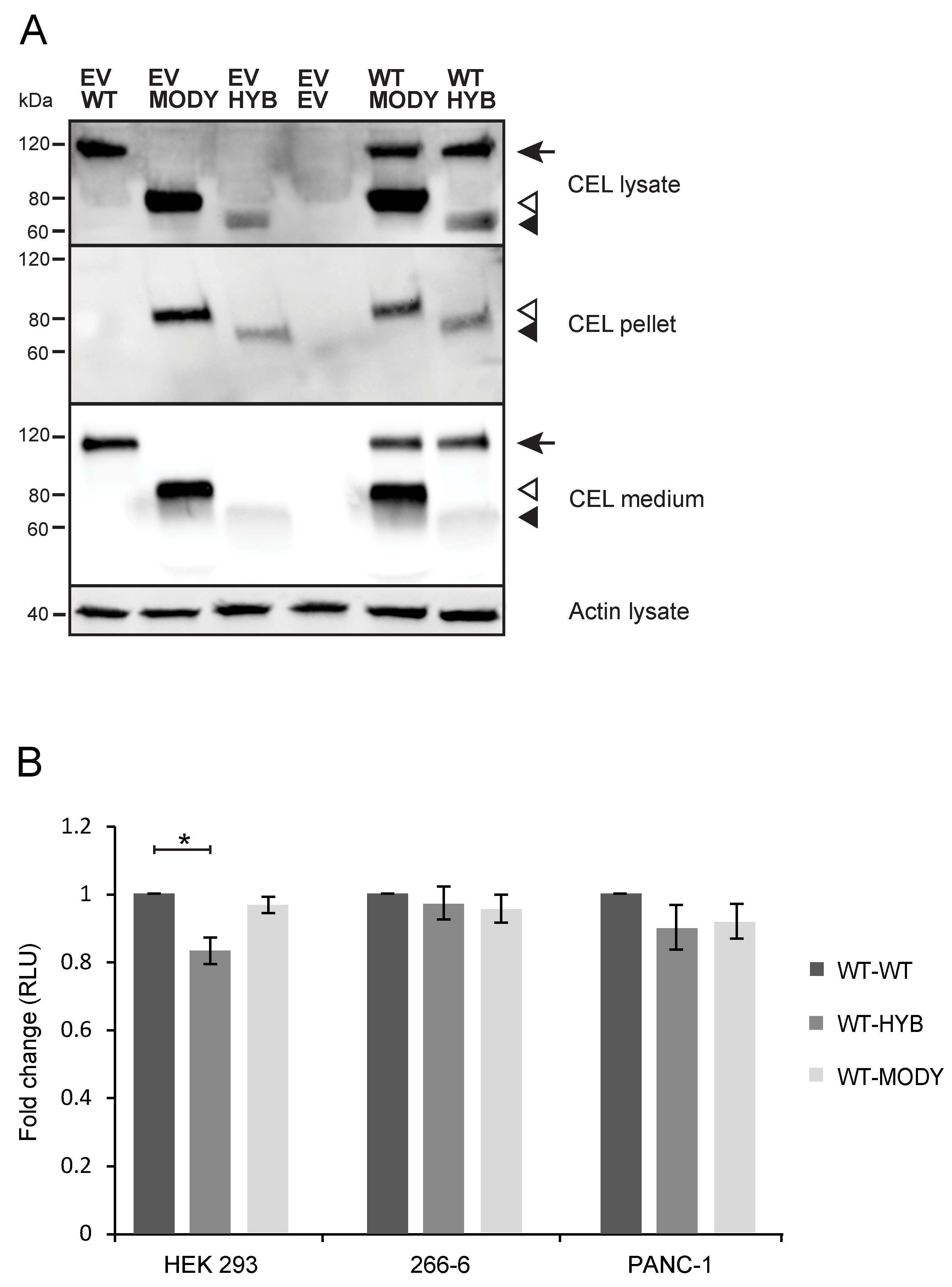
3.5. Cointernalized CEL-WT May Diminish the Toxic Effects Conferred by CEL-HYB and CEL-MODY in Pancreatic Cell Line Models
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lombardo, D.; Guy, O.; Figarella, C. Purification and characterization of a carboxyl ester hydrolase from human pancreatic juice. Biochim. Biophys. Acta 1978, 527, 142–149. [Google Scholar] [CrossRef]
- Lombardo, D.; Fauvel, J.; Guy, O. Studies on the substrate specificity of a carboxyl ester hydrolase from human pancreatic juice. I. Action on carboxyl esters, glycerides and phospholipids. Biochim. Biophys. Acta 1980, 611, 136–146. [Google Scholar] [CrossRef]
- Lombardo, D.; Guy, O. Studies on the substrate specificity of a carboxyl ester hydrolase from human pancreatic juice. II. Action on cholesterol esters and lipid-soluble vitamin esters. Biochim. Biophys. Acta 1980, 611, 147–155. [Google Scholar] [CrossRef]
- Hui, D.Y.; Howles, P.N. Carboxyl ester lipase: Structure-function relationship and physiological role in lipoprotein metabolism and atherosclerosis. J. Lipid. Res. 2002, 43, 2017–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolar, M.J.; Kamat, S.S.; Parsons, W.H.; Homan, E.A.; Maher, T.; Peroni, O.D.; Syed, I.; Fjeld, K.; Molven, A.; Kahn, B.B.; et al. Branched fatty acid esters of hydroxy fatty acids are preferred substrates of the MODY8 protein carboxyl ester lipase. Biochemistry 2016, 55, 4636–4641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombardo, D. Bile salt-dependent lipase: Its pathophysiological implications. Biochim. Biophys. Acta 2001, 1533, 1–28. [Google Scholar] [CrossRef]
- Bruneau, N.; Lombardo, D. Chaperone function of a Grp 94-related protein for folding and transport of the pancreatic bile salt-dependent lipase. J. Biol. Chem. 1995, 270, 13524–13533. [Google Scholar] [CrossRef] [Green Version]
- Abouakil, N.; Mas, E.; Bruneau, N.; Benajiba, A.; Lombardo, D. Bile salt-dependent lipase biosynthesis in rat pancreatic AR 4–2 J cells. Essential requirement of N-linked oligosaccharide for secretion and expression of a fully active enzyme. J. Biol. Chem. 1993, 268, 25755–25763. [Google Scholar]
- Bruneau, N.; Nganga, A.; Fisher, E.A.; Lombardo, D. O-Glycosylation of C-terminal tandem-repeated sequences regulates the secretion of rat pancreatic bile salt-dependent lipase. J. Biol. Chem. 1997, 272, 27353–27361. [Google Scholar] [CrossRef] [Green Version]
- Rogers, S.; Wells, R.; Rechsteiner, M. Amino acid sequences common to rapidly degraded proteins: The PEST hypothesis. Science 1986, 234, 364–368. [Google Scholar] [CrossRef]
- Loomes, K.M.; Senior, H.E.; West, P.M.; Roberton, A.M. Functional protective role for mucin glycosylated repetitive domains. Eur. J. Biochem. 1999, 266, 105–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, B.B.; Fjeld, K.; El Jellas, K.; Gravdal, A.; Dalva, M.; Tjora, E.; Ræder, H.; Kulkarni, R.N.; Johansson, S.; Njølstad, P.R.; et al. The role of the carboxyl ester lipase (CEL) gene in pancreatic disease. Pancreatology 2018, 18, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, S.; Nakamura, Y.; Saito, S. Characterization of a VNTR polymorphism in the coding region of the CEL gene. J. Hum. Genet. 2002, 47, 213–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindquist, S.; Blackberg, L.; Hernell, O. Human bile salt-stimulated lipase has a high frequency of size variation due to a hypervariable region in exon 11. Eur. J. Biochem. 2002, 269, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Torsvik, J.; Johansson, S.; Johansen, A.; Ek, J.; Minton, J.; Ræder, H.; Ellard, S.; Hattersley, A.; Pedersen, O.; Hansen, T.; et al. Mutations in the VNTR of the carboxyl-ester lipase gene (CEL) are a rare cause of monogenic diabetes. Hum. Genet. 2010, 127, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Ragvin, A.; Fjeld, K.; Weiss, F.U.; Torsvik, J.; Aghdassi, A.; Mayerle, J.; Simon, P.; Njølstad, P.R.; Lerch, M.M.; Johansson, S.; et al. The number of tandem repeats in the carboxyl-ester lipase (CEL) gene as a risk factor in alcoholic and idiopathic chronic pancreatitis. Pancreatology 2013, 13, 29–32. [Google Scholar] [CrossRef]
- Dalva, M.; El Jellas, K.; Steine, S.J.; Johansson, B.B.; Ringdal, M.; Torsvik, J.; Immervoll, H.; Hoem, D.; Laemmerhirt, F.; Simon, P.; et al. Copy number variants and VNTR length polymorphisms of the carboxyl-ester lipase (CEL) gene as risk factors in pancreatic cancer. Pancreatology 2016, 17, 83–88. [Google Scholar] [CrossRef]
- Fjeld, K.; Beer, S.; Johnstone, M.; Zimmer, C.; Mössner, J.; Ruffert, C.; Krehan, M.; Zapf, C.; Njølstad, P.R.; Johansson, S.; et al. Length of variable numbers of tandem repeats in the carboxyl ester lipase (CEL) gene may confer susceptibility to alcoholic liver cirrhosis but not alcoholic chronic pancreatitis. PLoS ONE 2016, 11, e0165567. [Google Scholar] [CrossRef] [Green Version]
- Ræder, H.; Johansson, S.; Holm, P.I.; Haldorsen, I.S.; Mas, E.; Sbarra, V.; Nermoen, I.; Eide, S.Å.; Grevle, L.; Bjørkhaug, L.; et al. Mutations in the CEL VNTR cause a syndrome of diabetes and pancreatic exocrine dysfunction. Nat. Gen. 2006, 38, 54–62. [Google Scholar] [CrossRef]
- Waldron, R.T.; Su, H.Y.; Piplani, H.; Capri, J.; Cohn, W.; Whitelegge, J.P.; Faull, K.F.; Sakkiah, S.; Abrol, R.; Yang, W.; et al. Ethanol Induced Disordering of Pancreatic Acinar Cell Endoplasmic Reticulum: An ER Stress/Defective Unfolded Protein Response Model. Cell Mol. Gastroenterol. Hepatol. 2018, 5, 479–497. [Google Scholar] [CrossRef] [Green Version]
- Johansson, B.B.; Torsvik, J.; Bjørkhaug, L.; Vesterhus, M.; Ragvin, A.; Tjora, E.; Fjeld, K.; Hoem, D.; Johansson, S.; Ræder, H.; et al. Diabetes and pancreatic exocrine dysfunction due to mutations in the carboxyl ester lipase gene-maturity onset diabetes of the young (CEL-MODY): A protein misfolding disease. J. Biol. Chem. 2011, 286, 34593–34605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torsvik, J.; Johansson, B.B.; Dalva, M.; Marie, M.; Fjeld, K.; Johansson, S.; Bjørkøy, G.; Saraste, J.; Njølstad, P.R.; Molven, A. Endocytosis of secreted carboxyl ester lipase in a syndrome of diabetes and pancreatic exocrine dysfunction. J. Biol. Chem. 2014, 289, 29097–29111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fjeld, K.; Weiss, F.U.; Lasher, D.; Rosendahl, J.; Chen, J.M.; Johansson, B.B.; Kirsten, H.; Ruffert, C.; Masson, E.; Steine, S.J.; et al. A recombined allele of the lipase gene CEL and its pseudogene CELP confers susceptibility to chronic pancreatitis. Nat. Genet. 2015, 47, 518–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, M.; McFadden, B.; Valiente, L.; Omori, K.; Bilbao, S.; Juan, J.; Rawson, J.; Oancea, A.R.; Scott, S.; Nair, I.; et al. Human Pancreatic Islets Isolated From Donors With Elevated HbA1c Levels: Islet Yield and Graft Efficacy. Cell Transplant. 2015, 24, 1879–1886. [Google Scholar] [CrossRef] [Green Version]
- Lugea, A.; Waldron, R.T.; Mareninova, O.A.; Shalbueva, N.; Deng, N.; Su, H.Y.; Thomas, D.D.; Jones, E.K.; Messenger, S.W.; Yang, J.; et al. Human Pancreatic Acinar Cells: Proteomic Characterization, Physiologic Responses, and Organellar Disorders in ex Vivo Pancreatitis. Am. J. Pathol. 2017, 187, 2726–2743. [Google Scholar] [CrossRef] [Green Version]
- El Jellas, K.; Hoem, D.; Hagen, K.G.; Kalvenes, M.B.; Aziz, S.; Steine, S.J.; Immervoll, H.; Johansson, S.; Molven, A. Associations between ABO blood groups and pancreatic ductal adenocarcinoma: Influence on resection status and survival. Cancer Med. 2017, 7, 1531–1540. [Google Scholar] [CrossRef] [Green Version]
- El Jellas, K.; Johansson, B.B.; Fjeld, K.; Antonopoulos, A.; Immervoll, H.; Choi, M.H.; Hoem, D.; Lowe, M.E.; Lombardo, D.; Njølstad, P.R.; et al. The mucinous domain of pancreatic carboxyl-ester lipase (CEL) contains core 1/core 2 O-glycans that can be modified by ABO blood group determinants. J. Biol. Chem. 2018, 293, 19476–19491. [Google Scholar] [CrossRef] [Green Version]
- Kayatekin, C.; Amasino, A.; Gaglia, G.; Flannick, J.; Bonner, J.M.; Fanning, S.; Narayan, P.; Barrasa, M.I.; Pincus, D.; Landgraf, D.; et al. Translocon Declogger Ste24 Protects against IAPP Oligomer-Induced Proteotoxicity. Cell 2018, 173, 62–73 e69. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, A.; Morales-Scheihing, D.; Salvadores, N.; Moreno-Gonzalez, I.; Gonzalez, C.; Taylor-Presse, K.; Mendez, N.; Shahnawaz, M.; Gaber, A.O.; Sabek, O.M.; et al. Induction of IAPP amyloid deposition and associated diabetic abnormalities by a prion-like mechanism. J. Exp. Med. 2017, 214, 2591–2610. [Google Scholar] [CrossRef]
- Lee, H.J.; Suk, J.E.; Bae, E.J.; Lee, J.H.; Paik, S.R.; Lee, S.J. Assembly-dependent endocytosis and clearance of extracellular alpha-synuclein. Int J. Biochem. Cell. Biol. 2008, 40, 1835–1849. [Google Scholar] [CrossRef]
- Xiao, X.; Jones, G.; Sevilla, W.A.; Stolz, D.B.; Magee, K.E.; Haughney, M.; Mukherjee, A.; Wang, Y.; Lowe, M.E. A carboxyl ester lipase (CEL) mutant causes chronic pancreatitis by forming intracellular aggregates that activate apoptosis. J. Biol. Chem. 2016, 291, 23224–23236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marciniak, S.J.; Ron, D. Endoplasmic reticulum stress signaling in disease. Physiol. Rev. 2006, 86, 1133–1149. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.S.; Li, W.M.; Chen, Y.N.; Zhao, Q.; Chen, Q.F. Endoplasmic reticulum stress is activated in acute pancreatitis. J. Dig. Dis. 2016, 17, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Molven, A.; Fjeld, K.; Lowe, M.E. Lipase genetic variants in chronic pancreatitis: When the end is wrong, all’s not well. Gastroenterology 2016, 150, 1515–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid, S.L.; Sorkin, A.; Zerial, M. Endocytosis: Past, present, and future. Cold Spring Harb. Perspect. Biol. 2014, 6, a022509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawakami, S.; Hashida, M. Glycosylation-mediated targeting of carriers. J. Control. Release 2014, 190, 542–555. [Google Scholar] [CrossRef] [Green Version]
- El Jellas, K.; Haslam, S.M.; Choi, M.H.; Dell, A.; Johansson, B.B.; Fjeld, K.; Molven, A. Altered O- and N-linked glycosylation profiles in carboxyl ester lipase (CEL) protein variants involved in chronic pancreatitis and MODY8 syndrome. Pancreatology 2018, 18, S119. [Google Scholar] [CrossRef]
- Ræder, H.; Haldorsen, I.S.; Ersland, L.; Grüner, R.; Taxt, T.; Søvik, O.; Molven, A.; Njølstad, P.R. Pancreatic lipomatosis is a structural marker in nondiabetic children with mutations in carboxyl-ester lipase. Diabetes 2007, 56, 444–449. [Google Scholar] [CrossRef] [Green Version]
- Ræder, H.; McAllister, F.E.; Tjora, E.; Bhatt, S.; Haldorsen, I.; Hu, J.; Willems, S.M.; Vesterhus, M.; El Ouaamari, A.; Liu, M.; et al. Carboxyl-ester lipase maturity-onset diabetes of the young is associated with development of pancreatic cysts and upregulated MAPK signaling in secretin-stimulated duodenal fluid. Diabetes 2014, 63, 259–269. [Google Scholar] [CrossRef] [Green Version]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dalva, M.; Lavik, I.K.; El Jellas, K.; Gravdal, A.; Lugea, A.; Pandol, S.J.; Njølstad, P.R.; Waldron, R.T.; Fjeld, K.; Johansson, B.B.; et al. Pathogenic Carboxyl Ester Lipase (CEL) Variants Interact with the Normal CEL Protein in Pancreatic Cells. Cells 2020, 9, 244. https://doi.org/10.3390/cells9010244
Dalva M, Lavik IK, El Jellas K, Gravdal A, Lugea A, Pandol SJ, Njølstad PR, Waldron RT, Fjeld K, Johansson BB, et al. Pathogenic Carboxyl Ester Lipase (CEL) Variants Interact with the Normal CEL Protein in Pancreatic Cells. Cells. 2020; 9(1):244. https://doi.org/10.3390/cells9010244
Chicago/Turabian StyleDalva, Monica, Ida K. Lavik, Khadija El Jellas, Anny Gravdal, Aurelia Lugea, Stephen J. Pandol, Pål R. Njølstad, Richard T. Waldron, Karianne Fjeld, Bente B. Johansson, and et al. 2020. "Pathogenic Carboxyl Ester Lipase (CEL) Variants Interact with the Normal CEL Protein in Pancreatic Cells" Cells 9, no. 1: 244. https://doi.org/10.3390/cells9010244
APA StyleDalva, M., Lavik, I. K., El Jellas, K., Gravdal, A., Lugea, A., Pandol, S. J., Njølstad, P. R., Waldron, R. T., Fjeld, K., Johansson, B. B., & Molven, A. (2020). Pathogenic Carboxyl Ester Lipase (CEL) Variants Interact with the Normal CEL Protein in Pancreatic Cells. Cells, 9(1), 244. https://doi.org/10.3390/cells9010244