Mechanistic Actions of microRNAs in Diabetic Wound Healing

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
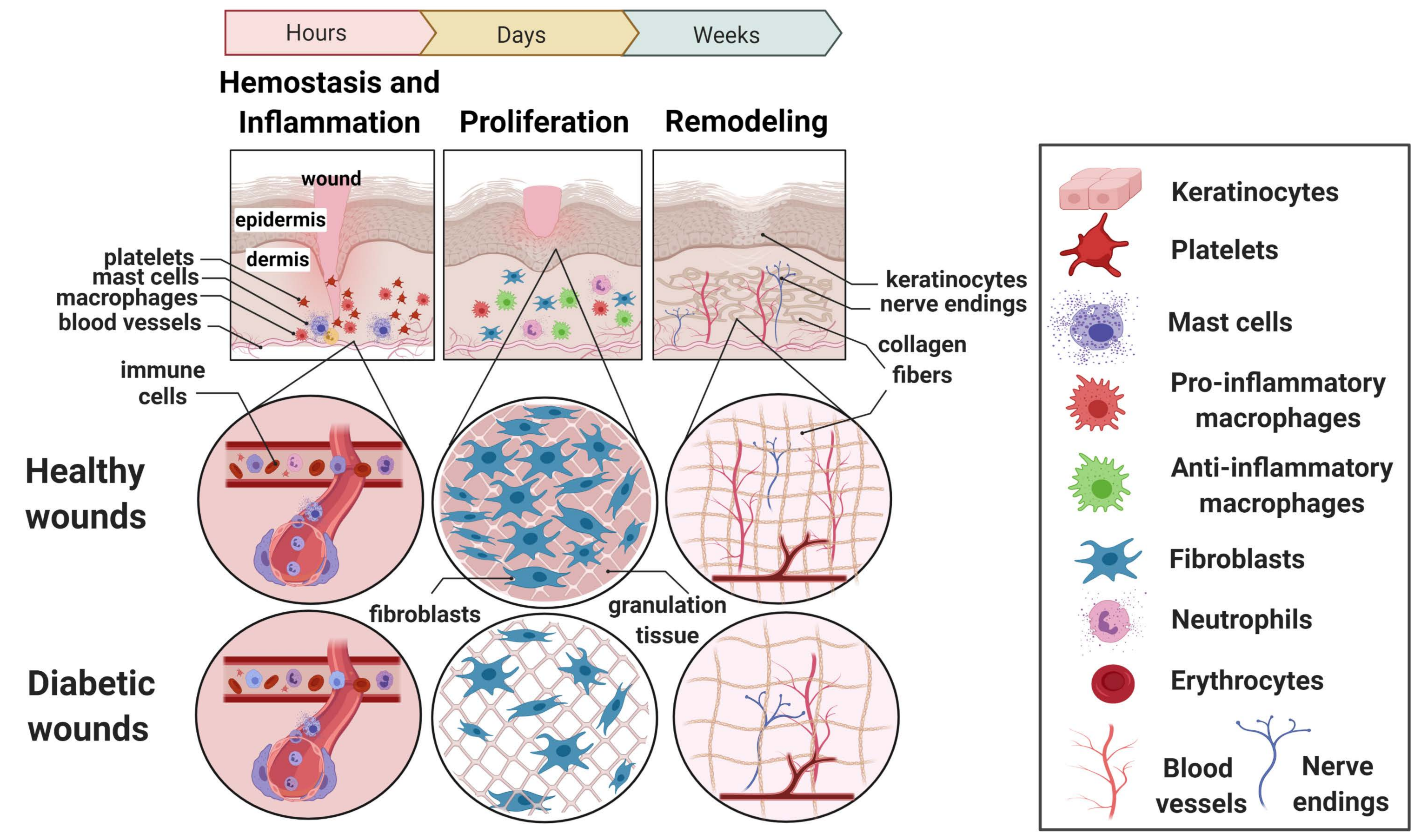
2. The Dynamic Phases of Normal Wound Healing
2.1. Hemostasis and the Inflammatory Phase
2.2. The Proliferative Phase
2.3. The Remodeling Phase
3. Impairments Observed in Wound Healing in Long-Standing Diabetes
3.1. Persistent Inflammation in the Non-Healing Wound Environment
3.2. Altered Macrophage Polarization in DFUs
3.3. Excessive Tissue Damage in Non-Healing Wounds
4. microRNAs
4.1. Biogenesis
4.2. Extracellular Roles of miRNA
5. microRNAs Altered in Diabetic Wound Healing
5.1. InflammiRs—microRNAs Having a Role in the Inflammatory Phase of Wound Healing
5.1.1. MiR-132 Up-Regulation Decreases Pro-Inflammatory Responses
5.1.2. MiR-146a Down-Regulation in Diabetic Wounds Results in Inflammation and Impaired Wound Healing
5.1.3. MiR-21 Promotes M1 Macrophage Polarization
5.1.4. MiR-155 Impairs Wound Re-Epithelization
5.1.5. MiR-223 Is a Wound Inflammation-Related miRNA
5.2. MiRNAs Affecting the Proliferative and Remodeling Phases of Diabetic Wound Healing
5.2.1. Hypoxia-Regulated miR-210 Downregulates the Growth and Differentiation of Keratinocytes
5.2.2. The miR-200 Family Is Hypoxia-Sensitive and Triggers the Angiogenic Response
5.2.3. Fibroblast Migration Is Dependent on miR-21 and miR-143/145
5.2.4. MiR-29 Family and Collagen Deposition
5.2.5. MiR-126 Has a Role in Vascular Inflammation
5.2.6. MiR-198 Restrains Cell Proliferation
5.2.7. The MiR-17∼92 Cluster Is Dysregulated in Ischemic Tissues
6. Circulating and Tissue microRNAs as Biomarkers for the Early Detection of Diabetic Foot Ulceration

{kind=link}
{kind=link}
{kind=link}
| Tissue specific microRNAs | ||||
| microRNA | Species | Expression pattern | Role in the tissue injury/repair | References |
| miR-132 | Humans Mouse | Reduced in human diabetic ulcers, upregulated in normal human wounds. Increased in diabetic mice wounds | Regulates the transition from inflammation to proliferation during wound healing | [121,124] |
| miR-146a | Human, Mouse | Increased in diabetic human skin, downregulated in mice wounds | Impairs inflammatory response observed in diabetic wounds | [136,138] |
| miR-21 | Human, Mouse | Elevated in human DFUs and rodent skin | Inhibits the inflammatory response | [151,153] |
| miR-155 | Human, Mouse | Increased in diabetic human and mice skin | Impairs wound re-epithelization | [151,178] |
| miR-223 | Human, Mouse | Highly expressed in wound skin | Controls neutrophil functions in acute inflammatory responses | [189,194] |
| miR-210 | Human, Mouse | High levels at the wound-edges | Impairs cell proliferative capacity | [127,128] |
| miR-200 | Human | Suppressed in diabetic wounds | Mediates the vascularization and cell migration | [141,142] |
| miR-126 | Human | Decreased in ulcers, increase after debridement therapy | Epithelialization and angiogenesis | [158,159] |
| miR-29 | Human, Mouse | Increased in diabetic skin, decrease with wounding | Repressor of collagen 1 and scar formation | [129,130] |
| miR-203 | Human | Increased in patients who underwent wound repairing surgery | Contributes to severity of the ulceration | [202] |
| Circulating microRNAs | ||||
| microRNA | Species | Expression pattern | Role in the tissue injury/repair | References |
| miR-217 | Rat | Increased in serum | Regulation of the VEGF pathway through HIF-1α inhibition | [203] |
| miR-145 | Human | Low in serum, higher in muscle from the amputated limbs | Fibroblast differentiation and suppression of cell growth | [184,204] |
| miR-16, -19b, -30e, -101, -144, -362, -451a, and -1260a | Human | Dysregulated in serum of patients with the Charcot foot (CF) and associated neuropathy | Monocytes differentiation | [205] |
| miR-191 and -200b | Human | Decreased in diabetes only, increased with chronic wounds and/or peripheral arterial disease (PAD) | Inflammation-mediated decrease of cellular migration and angiogenesis | [140] |
| miR-4739 | Human | Increased in plasma and serum obtained from critical limb ischemia (CLI) | Critical limb ischemia including amputation | [206,207] |
| miR-129 and-335 | Human | Skin and serum samples from patients with DFU | Impair wound healing due to elevated levels of MMP-9 together with transcription factor specificity protein 1 (Sp-1) | [208] |
6.1. Circulating miRNAs Related to Diabetic Complications and Wound Healing
6.2. MiRNAs in Diabetic Wounds as Possible Biomarkers for Wound Healing
6.3. Influence of Extracellular Vesicle miRNAs on Wound Healing
6.3.1. miRNAs in Macrophage-Derived EVs
6.3.2. Endothelial Cell Derived EVs and Their miRNAs
6.3.3. miRNAs Derived from MSC EVs Acting on Wound Healing
6.3.4. Circulating EV-Derived miRNAs and Their Actions in Wound Healing
| Phases of Wound Healing | microRNA | Action in Wound Healing | Reference |
|---|---|---|---|
 | miR-132 | Regulation of pro-inflammatory responses | [121,122,123,124] |
| miR-146a | Inflammatory responses | [136,137,138,139] | |
| miR-21 | Promotes M1 macrophage polarization | [151,152,153,154,155,156,157] | |
| miR-155 | Wound re-epithelization | [151,169,170,171,172,173,174,175,176,177,178,179] | |
| miR-223 | Inflammatory responses | [58,186,187,188,189,190,191,192,193,194] | |
 | miR-210 | Growth and differentiation of keratinocytes | [125,126,127,128] |
| miR-200 | Angiogenic response | [140,141,142,143,144] | |
| miR-126 | Re-epithelialization and angiogenesis | [158,159,160,161,162,163] | |
| miR-17~92 cluster | Migration of keratinocytes and re-epithelialization of the wound | [180,181,182,183] | |
 | miR-29 | Collagen restructuring and scar formation | [129,130,131,132,133,134,135] |
| miR-198 | Cell proliferation | [145,146,147,148,149,150] | |
| miR-21 | Migration of keratinocytes and wound re-epithelialization | [164,165,166,167,168] | |
| miR-143/ miR-145 | Cell differentiation, proliferation, angiogenesis and apoptosis | [184,185] |
7. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sorg, H.; Tilkorn, D.J.; Hager, S.; Hauser, J.; Mirastschijski, U. Skin wound healing: An update on the current knowledge and concepts. Eur. Surg. Res. 2017, 58, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Singer, A.J.; Clark, R.A.F. Mechanisms of disease: Cutaneous wound healing. N. Engl. J. Med. 1999, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Otranto, M.; Do Nascimento, A.P.; Monte-Alto-Costa, A. Insulin resistance impairs cutaneous wound healing in mice. Wound Repair Regen. 2013, 21, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Ceilley, R. Chronic wound healing: A review of current management and treatments. Adv. Ther. 2017, 34, 599–610. [Google Scholar] [CrossRef] [Green Version]
- Eleftheriadou, I.; Tentolouris, A.; Grigoropoulou, P.; Tsilingiris, D.; Anastasiou, I.; Kokkinos, A.; Perrea, D.; Katsilambros, N.; Tentolouris, N. The association of diabetic microvascular and macrovascular disease with cutaneous circulation in patients with type 2 diabetes mellitus. J. Diabetes Complicat. 2019, 33, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, S.; Schaper, N.; Apelqvist, J.; Jude, E. A review of the eurodiale studies: What lessons for diabetic foot care? Curr. Diab. Rep. 2011, 11, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Eming, S.A.; Krieg, T.; Davidson, J.M. Inflammation in wound repair: Molecular and cellular mechanisms. J. Investig. Dermatol. 2007, 127, 514–525. [Google Scholar] [CrossRef] [Green Version]
- Emanuelli, T.; Burgeiro, A.; Carvalho, E. Effects of insulin on the skin: Possible healing benefits for diabetic foot ulcers. Arch. Dermatol. Res. 2016, 308, 677–694. [Google Scholar] [CrossRef]
- Yang, G.; Li, C.; Gong, Y.; Fang, F.; Tian, H.; Li, J.; Cheng, X. Assessment of insulin resistance in subjects with normal glucose tolerance, hyperinsulinemia with normal blood glucose tolerance, impaired glucose tolerance, and newly diagnosed type 2 diabetes (prediabetes insulin resistance research). J. Diabetes Res. 2016, 2016, 11. [Google Scholar] [CrossRef] [Green Version]
- Granjon, A.; Gustin, M.P.; Rieusset, J.; Lefai, E.; Meugnier, E.; Güller, I.; Cerutti, C.; Paultre, C.; Disse, E.; Rabasa-Lhoret, R.; et al. The microRNA signature in response to insulin reveals its implication in the transcriptional action of insulin in human skeletal muscle and the role of a sterol regulatory element-binding protein-1c/myocyte enhancer factor 2C pathway. Diabetes 2009, 58, 2555–2564. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, A.; Shankar, K.; Beg, M.; Rajan, S.; Gupta, A.; Varshney, S.; Kumar, D.; Gupta, S.; Mishra, R.K.; Gaikwad, A.N. Chronic hyperinsulinemia induced miR-27b is linked to adipocyte insulin resistance by targeting insulin receptor. J. Mol. Med. 2018, 96, 315–331. [Google Scholar] [CrossRef] [PubMed]
- Alejandro, E.U.; Jo, S.; Akhaphong, B.; Llacer, P.R.; Gianchandani, M.; Gregg, B.; Parlee, S.D.; MacDougald, O.A.; Bernal-Mizrachi, E. Maternal low-protein diet on the last week of pregnancy contributes to insulin resistance and β-cell dysfunction in the mouse offspring. Am. J. Physiol. Integr. Comp. Physiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.A.; Younes, S.N.; Raza, S.S.; Zarif, L.; Nisar, S.; Ahmed, I.; Mir, R.; Kumar, S.; Sharawat, S.K.; Hashem, S.; et al. Role of non-coding RNA networks in leukemia progression, metastasis and drug resistance. Mol. Cancer 2020, 19, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Tomasetti, M.; Gaetani, S.; Monaco, F.; Neuzil, J.; Santarelli, L. Epigenetic regulation of miRNA expression in malignant mesothelioma: miRNAs as biomarkers of early diagnosis and therapy. Front. Oncol. 2019, 9, 1293. [Google Scholar] [CrossRef]
- Morales, S.; Monzo, M.; Navarro, A. Epigenetic regulation mechanisms of microRNA expression. Biomol. Concepts 2017, 8, 203–212. [Google Scholar] [CrossRef]
- Falanga, V. Wound healing and its impairment in the diabetic foot. Lancet 2005, 366, 1736–1743. [Google Scholar] [CrossRef]
- Sen, C.K.; Gordillo, G.M.; Roy, S.; Kirsner, R.; Lambert, L.; Hunt, T.K.; Gottrup, F.; Gurtner, G.C.; Longaker, M.T. Human skin wounds: A major and snowballing threat to public health and the economy. Wound Repair Regen. 2009, 17, 763–771. [Google Scholar] [CrossRef] [Green Version]
- De Groot, P.G.; Urbanus, R.T.; Roest, M. Platelet interaction with the vessel wall. Handb. Exp. Pharmacol. 2012, 87–110. [Google Scholar] [CrossRef]
- Castellana, D.; Paus, R.; Perez-Moreno, M. Macrophages contribute to the cyclic activation of adult hair follicle stem cells. PLoS Biol. 2014, 12, 12. [Google Scholar] [CrossRef] [Green Version]
- Minutti, C.M.; Knipper, J.A.; Allen, J.E.; Zaiss, D.M.W. Tissue-specific contribution of macrophages to wound healing. Semin. Cell Dev. Biol. 2017, 61, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Vannella, K.M.; Wynn, T.A. Mechanisms of organ injury and repair by macrophages. Annu. Rev. Physiol. 2017, 79, 593–617. [Google Scholar] [CrossRef] [PubMed]
- Krzyszczyk, P.; Schloss, R.; Palmer, A.; Berthiaume, F. The role of macrophages in acute and chronic wound healing and interventions to promote pro-wound healing phenotypes. Front. Physiol. 2018, 9, 419. [Google Scholar] [CrossRef] [PubMed]
- Baum, C.L.; Arpey, C.J. Normal cutaneous wound healing: Clinical correlation with cellular and molecular events. Dermatol. Surg. 2005, 31, 674–686. [Google Scholar] [CrossRef]
- Seraphim, P.M.; Leal, E.C.; Moura, J.; Gonçalves, P.; Gonçalves, J.P.; Carvalho, E. Lack of lymphocytes impairs macrophage polarization and angiogenesis in diabetic wound healing. Life Sci. 2020, 254. [Google Scholar] [CrossRef] [PubMed]
- Moura, J.; Rodrigues, J.; Gonçalves, M.; Amaral, C.; Lima, M.; Carvalho, E. Impaired T-cell differentiation in diabetic foot ulceration. Cell. Mol. Immunol. 2016, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, P.-H.; Sermersheim, M.; Li, H.; Lee, P.; Steinberg, S.; Ma, J. Zinc in wound healing modulation. Nutrients 2017, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Tanno, H.; Kawakami, K.; Kanno, E.; Suzuki, A.; Takagi, N.; Yamamoto, H.; Ishii, K.; Imai, Y.; Maruyama, R.; Tachi, M. Invariant NKT cells promote skin wound healing by preventing a prolonged neutrophilic inflammatory response. Wound Repair Regen. 2017, 25, 805–815. [Google Scholar] [CrossRef]
- Nosbaum, A.; Prevel, N.; Truong, H.-A.; Mehta, P.; Ettinger, M.; Scharschmidt, T.C.; Ali, N.H.; Pauli, M.L.; Abbas, A.K.; Rosenblum, M.D. Cutting edge: Regulatory T cells facilitate cutaneous wound healing. J. Immunol. 2016, 196, 2010–2014. [Google Scholar] [CrossRef]
- Li, J.; Chen, J.; Kirsner, R. Pathophysiology of acute wound healing. Clin. Dermatol. 2007, 25, 9–18. [Google Scholar] [CrossRef]
- Cheng, F.; Shen, Y.; Mohanasundaram, P.; Lindström, M.; Ivaska, J.; Ny, T.; Erikss, J.E. Vimentin coordinates fibroblast proliferation and keratinocyte differentiation in wound healing via TGF-β-Slug signaling. Proc. Natl. Acad. Sci. USA 2016, 113, E4320–E4327. [Google Scholar] [CrossRef] [Green Version]
- Heun, Y.; Anton, M.; Pfeifer, A.; Plank, C.; Pohl, U.; Mannell, H. Lentiviral magnetic targeting of SHP-2 locally governs HIF-1a dependent wound healing angiogenesis in vivo. J. Vasc. Res. 2017, 37. [Google Scholar]
- Heun, Y.; Pogoda, K.; Anton, M.; Pircher, J.; Pfeifer, A.; Woernle, M.; Ribeiro, A.; Kameritsch, P.; Mykhaylyk, O.; Plank, C.; et al. HIF-1α dependent wound healing angiogenesis in vivo can be controlled by site-specific lentiviral magnetic targeting of SHP-2. Mol. Ther. 2017, 25, 1616–1627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singampalli, K.L.; Balaji, S.; Wang, X.; Parikh, U.M.; Kaul, A.; Gilley, J.; Birla, R.K.; Bollyky, P.L.; Keswani, S.G. The role of an IL-10/hyaluronan axis in dermal wound healing. Front. Cell Dev. Biol. 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Sindrilaru, A.; Scharffetter-Kochanek, K. Disclosure of the culprits: Macrophages—Versatile regulators of wound healing. Adv. Wound Care 2013, 2, 357–368. [Google Scholar] [CrossRef] [Green Version]
- De Wever, O.; Demetter, P.; Mareel, M.; Bracke, M. Stromal myofibroblasts are drivers of invasive cancer growth. Int. J. Cancer 2008, 2229–2238. [Google Scholar] [CrossRef]
- Rodrigues, M.; Kosaric, N.; Bonham, C.A.; Gurtner, G.C. Wound healing: A cellular perspective. Physiol. Rev. 2019, 99, 665–706. [Google Scholar] [CrossRef]
- Ogawa, R. Keloid and hypertrophic scars are the result of chronic inflammation in the reticular dermis. Int. J. Mol. Sci. 2017, 18, 606. [Google Scholar] [CrossRef] [Green Version]
- Roshan, A.; Murai, K.; Fowler, J.; Simons, B.D.; Nikolaidou-Neokosmidou, V.; Jones, P.H. Human keratinocytes have two interconvertible modes of proliferation. Nat. Cell Biol. 2016, 18, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Martin, P.; Nunan, R. Cellular and molecular mechanisms of repair in acute and chronic wound healing. Br. J. Dermatol. 2015, 173, 370–378. [Google Scholar] [CrossRef]
- Landén, N.X.; Li, D.; Ståhle, M.; Sciences, M.L.; Landén, N.X.; Li, D.; Ståhle, M. Transition from inflammation to proliferation: A critical step during wound healing. Cell. Mol. Life Sci. 2016, 73, 3861–3885. [Google Scholar] [CrossRef] [Green Version]
- Golebiewska, E.M.; Poole, A.W. Platelet secretion: From haemostasis to wound healing and beyond. Blood Rev. 2015, 29, 153–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinish, M.; Cui, W.; Stafford, E.; Bae, L.; Hawkins, H.; Cox, R.; Toliver-Kinsky, T. Dendritic cells modulate burn wound healing by enhancing early proliferation. Wound Repair Regen. 2016, 24, 6–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Wu, J.; Luo, G.; He, W. Functions of Vγ4 T cells and dendritic epidermal T cells on skin wound healing. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J. Neutrophils in tissue injury and repair. Cell Tissue Res. 2018, 371, 531–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, E.L.; Bruyninckx, W.J.; Kelly, C.J.; Glover, L.E.; McNamee, E.N.; Bowers, B.E.; Bayless, A.J.; Scully, M.; Saeedi, B.J.; Golden-Mason, L.; et al. Transmigrating neutrophils shape the mucosal microenvironment through localized oxygen depletion to influence resolution of inflammation. Immunity 2014, 40, 66–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enyedi, B.; Niethammer, P. Mechanisms of epithelial wound detection. Trends Cell Biol. 2015, 25, 398–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dees, C.; Chakraborty, D.; Distler, J.H.W. Cellular and molecular mechanisms in fibrosis. Exp. Dermatol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Koyasu, S.; Moro, K. Role of innate lymphocytes in infection and inflammation. Front. Immunol. 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Nair, M.G. Macrophages in wound healing: Activation and plasticity. Immunol. Cell Biol. 2019, 97, 258–267. [Google Scholar] [CrossRef]
- Boniakowski, A.E.; Kimball, A.S.; Jacobs, B.N.; Kunkel, S.L.; Gallagher, K.A. Macrophage-mediated inflammation in normal and diabetic wound healing. J. Immunol. 2017, 199, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Tosello-Trampont, A.; Surette, F.A.; Ewald, S.E.; Hahn, Y.S. Immunoregulatory role of NK cells in tissue inflammation and regeneration. Front. Immunol. 2017, 8, 301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, H.C.; Popi, A.F.; Bachi, A.L.L.; Nonogaki, S.; Lopes, J.D.; Mariano, M. B-1 cells modulate the kinetics of wound-healing process in mice. Immunobiology 2010, 215, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Toulon, A.; Breton, L.; Taylor, K.R.; Tenenhaus, M.; Bhavsar, D.; Lanigan, C.; Rudolph, R.; Jameson, J.; Havran, W.L. A role for human skin-resident T cells in wound healing. J. Exp. Med. 2009, 206, 743–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreau, J.M.; Rosenblum, M.D. Inflammatory T cells maintain a healing disposition. Sci. Immunol. 2019, 4. [Google Scholar] [CrossRef] [PubMed]
- Ashrafi, M.; Baguneid, M.; Bayat, A. The role of neuromediators and innervation in cutaneous wound healing. Acta Derm. Venereol. 2016, 96, 587–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal, E.C.; Carvalho, E.; Tellechea, A.; Kafanas, A.; Tecilazich, F.; Kearney, C.; Veves, A.; Kuchibhotla, S.; Auster, M.E.; Kokkotou, E.; et al. Substance P promotes wound healing in diabetes by modulating in fl ammation and macrophage phenotype. Am. J. Pathol. 2015, 185, 1638–1648. [Google Scholar] [CrossRef]
- Pereira, L.; Neves, B.M.; Moura, L.; Cruz, M.T.; Carvalho, E. Neurotensin decreases the proinflammatory status of human skin fibroblasts and increases epidermal growth factor expression. Int. J. Inflam. 2014, 2014. [Google Scholar] [CrossRef] [Green Version]
- Pradhan, L.; Cai, X.; Wu, S.; Andersen, N.D.; Martin, M.; Malek, J.; Guthrie, P.; Veves, A.; Logerfo, F.W. Gene expression of pro-inflammatory cytokines and neuropeptides in diabetic wound healing. J. Surg. Res. 2011, 167, 336–342. [Google Scholar] [CrossRef] [Green Version]
- Wojewodzka, U.; Rosinski, G.; Olszewski, W.L.; Karnafel, W.; Galkowska, H. Neurogenic factors in the impaired healing of diabetic foot ulcers. J. Surg. Res. 2006, 134, 252–258. [Google Scholar] [CrossRef]
- Amin, N.; Doupis, J. Diabetic foot disease: From the evaluation of the “foot at risk” to the novel diabetic ulcer treatment modalities. World J. Diabetes 2016, 7, 153. [Google Scholar] [CrossRef]
- Diegelmann, R.F.; Evans, M.C. Wound healing: An overview of acute, fibrotic and delayed healing. Front. Biosci. 2004, 9, 283–289. [Google Scholar] [CrossRef]
- Eming, S.A.; Martin, P.; Tomic-Canic, M. Wound repair and regeneration: Mechanisms, signaling, and translation. Sci. Transl. Med. 2014, 6. [Google Scholar] [CrossRef] [Green Version]
- Demidova-Rice, T.N.; Hamblin, M.R.; Herman, I. Acute and impaired wound healing: Pathophysiology and current methods for drug delivery, part 2: Role of growth factors in normal and pathological wound healing: Therapeutic potential and methods of delivery. Adv. Ski. Wound Care 2012, 25, 304–314. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.-J.; Velazquez, O.C. Hyperoxia, endothelial progenitor cell mobilization, and diabetic wound healing. Antioxid. Redox Signal. 2008, 10, 1869–1882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tellechea, A.; Silva, E.A.; Min, J.; Leal, E.C.; Auster, M.E.; Pradhan-Nabzdyk, L.; Shih, W.; Mooney, D.J.; Veves, A. Alginate and DNA gels are suitable delivery systems for diabetic wound healing. Int. J. Low. Extrem. Wounds 2015, 14, 146–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moura, J.; Madureira, P.; Leal, E.C.; Fonseca, A.C.; Carvalho, E. Immune aging in diabetes and its implications in wound healing. Clin. Immunol. 2019, 200, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Shannon, J.D.; Eming, S.A.; Kreft, S.; Krieger, A.; Koch, M.; Fox, J.W.; Krieg, T.; Bruckner-Tuderman, L.; Brachvogel, B. Differential proteomic analysis distinguishes tissue repair biomarker signatures in wound exudates obtained from normal healing and chronic wounds. J. Proteome Res. 2010, 9, 4758–4766. [Google Scholar] [CrossRef]
- Loots, M.A.M.; Lamme, E.N.; Zeegelaar, J.; Mekkes, J.R.; Bos, J.D.; Middelkoop, E. Differences in cellular infiltrate and extracellular matrix of chronic diabetic and venous ulcers versus acute wounds. J. Investig. Dermatol. 1998, 111, 850–857. [Google Scholar] [CrossRef] [Green Version]
- Tellechea, A.; Leal, E.C.; Kafanas, A.; Auster, M.E.; Kuchibhotla, S.; Ostrovsky, Y.; Tecilazich, F.; Baltzis, D.; Zheng, Y.; Carvalho, E.; et al. Mast cells regulate wound healing in diabetes. Diabetes 2016, 65, 2006–2019. [Google Scholar] [CrossRef] [Green Version]
- Wicks, K.; Torbica, T.; Mace, K.A. Myeloid cell dysfunction and the pathogenesis of the diabetic chronic wound. Semin. Immunol. 2014, 26, 341–353. [Google Scholar] [CrossRef]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef] [PubMed]
- Khanna, S.; Biswas, S.; Shang, Y.; Collard, E.; Azad, A.; Kauh, C.; Bhasker, V.; Gordillo, G.M.; Sen, C.K.; Roy, S. Macrophage dysfunction impairs resolution of inflammation in the wounds of diabetic mice. PLoS ONE 2010, 5, e9539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siqueira, M.F.; Li, J.; Chehab, L.; Desta, T.; Chino, T.; Krothpali, N.; Behl, Y.; Alikhani, M.; Yang, J.; Braasch, C.; et al. Impaired wound healing in mouse models of diabetes is mediated by TNF-α dysregulation and associated with enhanced activation of forkhead box O1 (FOXO1). Diabetologia 2010, 53, 378–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goren, I.; Allmann, N.; Yogev, N.; Schürmann, C.; Linke, A.; Holdener, M.; Waisman, A.; Pfeilschifter, J.; Frank, S. A transgenic mouse model of inducible macrophage depletion: Effects of diphtheria toxin-driven lysozyme m-specific cell lineage ablation on wound inflammatory, angiogenic, and contractive processes. Am. J. Pathol. 2009, 175, 132–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Momtazi, M.; Ding, J.; Kwan, P.; Anderson, C.C.; Honardoust, D.; Goekjian, S.; Tredget, E.E. Morphologic and histologic comparison of hypertrophic scar in nude mice, t-cell receptor, and recombination activating gene knockout mice. Plast. Reconstr. Surg. 2015, 136, 1192–1204. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Ding, J.; Ma, Z.; Iwashina, T.; Tredget, E.E. Systemic depletion of macrophages in the subacute phase of wound healing reduces hypertrophic scar formation. Wound Repair Regen. 2016, 24, 644–656. [Google Scholar] [CrossRef] [PubMed]
- Schett, G.; Neurath, M.F. Resolution of chronic inflammatory disease: Universal and tissue-specific concepts. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef]
- Larouche, J.; Sheoran, S.; Maruyama, K.; Martino, M.M. Immune regulation of skin wound healing: Mechanisms and novel therapeutic targets. Adv. Wound Care 2018, 7, 209–231. [Google Scholar] [CrossRef]
- Dovi, J.V.; Szpaderska, A.M.; DiPietro, L.A. Neutrophil function in the healing wound: Adding insult to injury? Thromb. Haemost. 2004, 92, 275–280. [Google Scholar] [CrossRef]
- Bourboulia, D.; Stetler-Stevenson, W.G. Matrix metalloproteinases (MMPs) and tissue inhibitors of metalloproteinases (TIMPs): Positive and negative regulators in tumor cell adhesion. Semin. Cancer Biol. 2010, 20, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Pirilä, E.; Korpi, J.T.; Korkiamäki, T.; Jahkola, T.; Gutierrez-Fernandez, A.; Lopez-Otin, C.; Saarialho-Kere, U.; Salo, T.; Sorsa, T. Collagenase-2 (MMP-8) and matrilysin-2 (MMP-26) expression in human wounds of different etiologies. Wound Repair Regen. 2007, 15, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Guo, S.; Yao, F.; Zhang, Y.; Li, T. Increased ratio of serum matrix metalloproteinase-9 against TIMP-1 predicts poor wound healing in diabetic foot ulcers. J. Diabetes Complicat. 2013, 27, 380–382. [Google Scholar] [CrossRef] [PubMed]
- Mustoe, T.A.; O’Shaughnessy, K.; Kloeters, O. Chronic Wound Pathogenesis and Current Treatment Strategies: A Unifying Hypothesis. Plast. Reconstr. Surg. 2006, 117, 35S–41S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.; Gregory, R.I. MicroRNA biogenesis pathways in cancer. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef]
- Lynn, F.C. Meta-regulation: microRNA regulation of glucose and lipid metabolism. Trends Endocrinol. Metab. 2009, 20, 452–459. [Google Scholar] [CrossRef]
- Vienberg, S.; Geiger, J.; Madsen, S.; Dalgaard, L.T. microRNAs in metabolism. Acta Physiol. 2016, 219, 346–361. [Google Scholar] [CrossRef]
- Wong, W.K.M.; Sørensen, A.E.; Joglekar, M.V.; Hardikar, A.A.; Dalgaard, L.T. Non-coding RNA in pancreas and β-cell development. Non-coding RNA 2018, 4, 41. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Wightman, B.; Ha, I.; Ruvkun, G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993, 75, 855–862. [Google Scholar] [CrossRef]
- Lagos-Quintana, M.; Rauhut, R.; Lendeckel, W.; Tuschl, T. Identification of novel genes coding for small expressed RNAs. Science 2001, 294, 853–858. [Google Scholar] [CrossRef] [Green Version]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2018, 47, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Huang, S.; Ding, J.; Zhao, Y.; Liang, L.; Liu, T.; Zhan, R.; He, X. Multiple microRNAs modulate p21Cip1/Waf1 expression by directly targeting its 3′ untranslated region. Oncogene 2010, 29, 2302–2308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA genes are transcribed by RNA polymerase II. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef]
- Lund, E.; Güttinger, S.; Calado, A.; Dahlberg, J.E.; Kutay, U. Nuclear export of MicroRNA precursors. Science 2004, 303, 95–98. [Google Scholar] [CrossRef] [Green Version]
- Bohnsack, M.T.; Czaplinski, K.; Görlich, D. Exportin 5 is a RanGTP-dependent dsRNA-binding protein that mediates nuclear export of pre-miRNAs. RNA 2004, 10, 185–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chendrimada, T.P.; Gregory, R.I.; Kumaraswamy, E.; Norman, J.; Cooch, N.; Nishikura, K.; Shiekhattar, R. TRBP recruits the Dicer complex to Ago2 for microRNA processing and gene silencing. Nature 2005, 436, 740–744. [Google Scholar] [CrossRef]
- Lee, Y.; Hur, I.; Park, S.Y.; Kim, Y.K.; Mi, R.S.; Kim, V.N. The role of PACT in the RNA silencing pathway. EMBO J. 2006, 25, 522–532. [Google Scholar] [CrossRef]
- Meijer, H.A.; Smith, E.M.; Bushell, M. Regulation of miRNA strand selection: Follow the leader? Biochem. Soc. Trans. 2014, 42, 1135–1140. [Google Scholar] [CrossRef]
- Chiang, H.R.; Schoenfeld, L.W.; Ruby, J.G.; Auyeung, V.C.; Spies, N.; Baek, D.; Johnston, W.K.; Russ, C.; Luo, S.; Babiarz, J.E.; et al. Mammalian microRNAs: Experimental evaluation of novel and previously annotated genes. Genes Dev. 2010, 24, 992–1009. [Google Scholar] [CrossRef] [Green Version]
- Huntzinger, E.; Izaurralde, E. Gene silencing by microRNAs: Contributions of translational repression and mRNA decay. Nat. Rev. Genet. 2011, 12, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Fan, J.; Belasco, J.G. microRNAs direct rapid deadenylation of mRNA. Proc. Natl. Acad. Sci. USA 2006, 103, 4034–4039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, H.W.; Wentzel, E.A.; Mendell, J.T. A hexanucleotide element directs microRNA nuclear import. Science 2007, 315, 97–100. [Google Scholar] [CrossRef]
- Rissland, O.S.; Hong, S.J.; Bartel, D.P. MicroRNA Destabilization enables dynamic regulation of the miR-16 family in response to cell-cycle changes. Mol. Cell 2011, 43, 993–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avraham, R.; Sas-Chen, A.; Manor, O.; Steinfeld, I.; Shalgi, R.; Tarcic, G.; Bossel, N.; Zeisel, A.; Amit, I.; Zwang, Y.; et al. EGF decreases the abundance of microRNAs that restrain oncogenic transcription factors. Sci. Signal. 2010, 3. [Google Scholar] [CrossRef] [Green Version]
- Krol, J.; Busskamp, V.; Markiewicz, I.; Stadler, M.B.; Ribi, S.; Richter, J.; Duebel, J.; Bicker, S.; Fehling, H.J.; Schübeler, D.; et al. Characterizing light-regulated retinal microRNAs reveals rapid turnover as a common property of neuronal microRNAs. Cell 2010, 141, 618–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ameres, S.L.; Horwich, M.D.; Hung, J.H.; Xu, J.; Ghildiyal, M.; Weng, Z.; Zamore, P.D. Target RNA-directed trimming and tailing of small silencing RNAs. Science 2010, 328, 1534–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bail, S.; Swerdel, M.; Liu, H.; Jiao, X.; Goff, L.A.; Hart, R.P.; Kiledjian, M. Differential regulation of microRNA stability. RNA 2010, 16, 1032–1039. [Google Scholar] [CrossRef] [Green Version]
- Das, S.K.; Sokhi, U.K.; Bhutia, S.K.; Azab, B.; Su, Z.Z.; Sarkar, D.; Fisher, P.B. Human polynucleotide phosphorylase selectively and preferentially degrades microRNA-221 in human melanoma cells. Proc. Natl. Acad. Sci. USA 2010, 107, 11948–11953. [Google Scholar] [CrossRef] [Green Version]
- Katoh, T.; Sakaguchi, Y.; Miyauchi, K.; Suzuki, T.; Suzuki, T.; Kashiwabara, S.I.; Baba, T. Selective stabilization of mammalian microRNAs by 3’ adenylation mediated by the cytoplasmic poly(A) polymerase GLD-2. Genes Dev. 2009, 23, 433–438. [Google Scholar] [CrossRef] [Green Version]
- Boele, J.; Persson, H.; Shin, J.W.; Ishizu, Y.; Newie, I.S.; Søkilde, R.; Hawkins, S.M.; Coarfa, C.; Ikeda, K.; Takayama, K.I.; et al. PAPD5-mediated 3′ adenylation and subsequent degradation of miR-21 is disrupted in proliferative disease. Proc. Natl. Acad. Sci. USA 2014, 111, 11467–11472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, G.; Cetin, S.; Messmer, M.; Chane-Woon-Ming, B.; Terenzi, O.; Chicher, J.; Kuhn, L.; Hammann, P.; Pfeffer, S. Identification of factors involved in target RNA-directed microRNA degradation. Nucleic Acids Res. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Van Niel, G.; Porto-Carreiro, I.; Simoes, S.; Raposo, G. Exosomes: A common pathway for a specialized function. J. Biochem. 2006, 140, 13–21. [Google Scholar] [CrossRef]
- Wollert, T.; Hurley, J.H. Molecular mechanism of multivesicular body biogenesis by ESCRT complexes. Nature 2010, 464, 864–869. [Google Scholar] [CrossRef] [Green Version]
- Baietti, M.F.; Zhang, Z.; Mortier, E.; Melchior, A.; Degeest, G.; Geeraerts, A.; Ivarsson, Y.; Depoortere, F.; Coomans, C.; Vermeiren, E.; et al. Syndecan-syntenin-ALIX regulates the biogenesis of exosomes. Nat. Cell Biol. 2012, 14, 677–685. [Google Scholar] [CrossRef]
- Ambros, V.; Lee, R.C. Identification of microRNAs and other tiny noncoding RNAs by cDNA CLoning. RNA Interf. Ed. Modif. 2009, 265, 131–158. [Google Scholar] [CrossRef]
- He, T.; Zhang, Y.; Liu, Y.; Wang, H.; Zhang, W.; Liu, J.; Li, N.; Li, Y.; Wang, L.; Xie, S.; et al. MicroRNA-494 targets PTEN and suppresses PI3K/AKT pathway to alleviate hypertrophic scar formation. J. Mol. Histol. 2019. [Google Scholar] [CrossRef]
- Wu, X.; Li, J.; Yang, X.; Bai, X.; Shi, J.; Gao, J.; Li, Y.; Han, S.; Zhang, Y.; Han, F.; et al. miR-155 inhibits the formation of hypertrophic scar fibroblasts by targeting HIF-1α via PI3K/AKT pathway. J. Mol. Histol. 2018. [Google Scholar] [CrossRef]
- Li, D.; Wang, A.; Liu, X.; Meisgen, F.; Grünler, J.; Botusan, I.R.; Narayanan, S.; Erikci, E.; Li, X.; Blomqvist, L.; et al. MicroRNA-132 enhances transition from inflammation to proliferation during wound healing. J. Clin. Investig. 2015, 125, 3008–3026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Essandoh, K.; Li, Y.; Fan, G. MiRNA-mediated macrophage polarization and its potential role in the regulation of inflammatory response. Shock 2016, 46, 122–131. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Jing, Z.; Cheng, G. microRNAs: New regulators of toll-like receptor signalling pathways. Biomed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Li, D.; Wang, A.; Chu, T.; Lohcharoenkal, W.; Zheng, X.; Grünler, J.; Narayanan, S.; Eliasson, S.; Herter, E.K.; et al. MicroRNA-132 with Therapeutic Potential in Chronic Wounds. J. Investig. Dermatol. 2017, 137, 2630–2638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannakakis, A.; Sandaltzopoulos, R.; Greshock, J.; Liang, S.; Huang, J.; Hasegawa, K.; Li, C.; O’Brien-Jenkins, A.; Katsaros, D.; Weber, B.L.; et al. miR-210 links hypoxia with cell cycle regulation and is deleted in human epithelial ovarian cancer. Cancer Biol. Ther. 2008. [Google Scholar] [CrossRef] [Green Version]
- Biswas, S.; Roy, S.; Banerjee, J.; Hussain, S.R.A.; Khanna, S.; Meenakshisundaram, G.; Kuppusamy, P.; Friedman, A.; Sen, C.K. Hypoxia inducible microRNA 210 attenuates keratinocyte proliferation and impairs closure in a murine model of ischemic wounds. Proc. Natl. Acad. Sci. USA 2010, 107, 6976–6981. [Google Scholar] [CrossRef] [Green Version]
- Ghatak, S.; Li, J.; Chan, Y.C.; Gnyawali, S.C.; Steen, E.; Yung, B.C.; Khanna, S.; Roy, S.; Lee, R.J.; Sen, C.K. AntihypoxamiR functionalized gramicidin lipid nanoparticles rescue against ischemic memory improving cutaneous wound healing. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 1827–1831. [Google Scholar] [CrossRef] [Green Version]
- Essandoh, K.; Li, Y.; Huo, J.; Fan, G.C. Downregulation of miR-210 promotes sulfur mustard-induced skin wound healing. Int. J. Clin. Exp. Med. 2019, 12, 5279–5287. [Google Scholar]
- Harmanci, D.; Erkan, E.P.; Kocak, A.; Akdogan, G.G. Role of the microRNA-29 family in fibrotic skin diseases. Biomed. Rep. 2017, 6, 599–604. [Google Scholar] [CrossRef] [Green Version]
- Gallant-Behm, C.L.; Piper, J.; Lynch, J.M.; Seto, A.G.; Hong, S.J.; Mustoe, T.A.; Maari, C.; Pestano, L.A.; Dalby, C.M.; Jackson, A.L.; et al. A MicroRNA-29 Mimic (Remlarsen) represses extracellular matrix expression and fibroplasia in the skin. J. Investig. Dermatol. 2019, 139, 1073–1081. [Google Scholar] [CrossRef]
- Cheng, J.; Yu, H.; Deng, S.; Shen, G. MicroRNA profiling in mid- and late-gestational fetal skin: Implication for scarless wound healing. Tohoku J. Exp. Med. 2010, 221, 203–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Rooij, E.; Sutherland, L.B.; Thatcher, J.E.; DiMaio, J.M.; Naseem, R.H.; Marshall, W.S.; Hill, J.A.; Olson, E.N. Dysregulation of microRNAs after myocardial infarction reveals a role of miR-29 in cardiac fibrosis. Proc. Natl. Acad. Sci. USA 2008, 105, 13027–13032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulholland, E.J.; Dunne, N.; McCarthy, H.O. MicroRNA as therapeutic targets for chronic wound healing. Mol. Ther. Nucleic Acids 2017, 8, 46–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zgheib, C.; Hodges, M.; Hu, J.; Beason, D.P.; Soslowsky, L.J.; Liechty, K.W.; Xu, J. Mechanisms of mesenchymal stem cell correction of the impaired biomechanical properties of diabetic skin: The role of miR-29a. Wound Repair Regen. 2016, 24, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Shilo, S.; Roy, S.; Khanna, S.; Sen, C.K. MicroRNA in cutaneous wound healing: A new paradigm. DNA Cell Biol. 2007, 26, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wu, W.; Zhang, L.; Dorset-Martin, W.; Morris, M.W.; Mitchell, M.E.; Liechty, K.W. The role of MicroRNA-146a in the pathogenesis of the diabetic wound-healing impairment: Correction with mesenchymal stem cell treatment. Diabetes 2012, 61, 2906–2912. [Google Scholar] [CrossRef] [Green Version]
- Meyer, S.E. MicroRNA immunomodulating therapeutics. Blood 2020, 135, 155–156. [Google Scholar] [CrossRef] [Green Version]
- Quinn, E.M.; Wang, J.H.; O’Callaghan, G.; Redmond, H.P. MicroRNA-146a Is upregulated by and negatively regulates TLR2 signaling. PLoS ONE 2013, 8, e62232. [Google Scholar] [CrossRef] [Green Version]
- Zgheib, C.; Hilton, S.A.; Dewberry, L.C.; Hodges, M.M.; Ghatak, S.; Xu, J.; Singh, S.; Roy, S.; Sen, C.K.; Seal, S.; et al. Use of cerium oxide nanoparticles conjugated with MicroRNA-146a to correct the diabetic wound healing impairment. J. Am. Coll. Surg. 2019, 228, 107–115. [Google Scholar] [CrossRef]
- Dangwal, S.; Stratmann, B.; Bang, C.; Lorenzen, J.M.; Kumarswamy, R.; Fiedler, J.; Falk, C.S.; Scholz, C.J.; Thum, T.; Tschoepe, D. Impairment of wound healing in patients with type 2 diabetes mellitus influences circulating MicroRNA patterns via inflammatory cytokines. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 1480–1488. [Google Scholar] [CrossRef] [Green Version]
- Chan, Y.C.; Khanna, S.; Roy, S.; Sen, C.K. MiR-200b targets Ets-1 and is down-regulated by hypoxia to induce angiogenic response of endothelial cells. J. Biol. Chem. 2011, 286, 2047–2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, R.C.; Pastar, I.; Ojeh, N.; Chen, V.; Liu, S.; Garzon, K.I.; Tomic-Canic, M. Epithelial-mesenchymal transition in tissue repair and fibrosis. Cell Tissue Res. 2016, 365, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Gregory, P.A.; Bert, A.G.; Paterson, E.L.; Barry, S.C.; Tsykin, A.; Farshid, G.; Vadas, M.A.; Khew-Goodall, Y.; Goodall, G.J. The miR-200 family and miR-205 regulate epithelial to mesenchymal transition by targeting ZEB1 and SIP1. Nat. Cell Biol. 2008, 10, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Aunin, E.; Broadley, D.; Ahmed, M.I.; Mardaryev, A.N.; Botchkareva, N.V. Exploring a role for regulatory miRNAs in wound healing during ageing:involvement of miR-200c in wound repair. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sylva, M.; Moorman, A.F.M.; Van den Hoff, M.J.B. Follistatin-like 1 in vertebrate development. Birth Defects Res. Part C Embryo Today Rev. 2013, 99, 61–69. [Google Scholar] [CrossRef]
- Sundaram, G.M.; Common, J.E.A.; Gopal, F.E.; Srikanta, S.; Lakshman, K.; Lunny, D.P.; Lim, T.C.; Tanavde, V.; Lane, E.B.; Sampath, P. ‘See-saw’ expression of microrna-198 and fstl1 from a single transcript in wound healing. Nature 2013, 495, 103–106. [Google Scholar] [CrossRef]
- Marin-Muller, C.; Li, D.; Bharadwaj, U.; Li, M.; Chen, C.; Hodges, S.E.; Fisher, W.E.; Mo, Q.; Hung, M.C.; Yao, Q. A tumorigenic factor interactome connected through tumor suppressor MicroRNA-198 in human pancreatic cancer. Clin. Cancer Res. 2013, 19, 5901–5913. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.; Li, R.; Ding, K.; Lobie, P.E.; Zhu, T. MiR-198 inhibits migration and invasion of hepatocellular carcinoma cells by targeting the HGF/c-MET pathway. FEBS Lett. 2011. [Google Scholar] [CrossRef] [Green Version]
- Lulla, R.R.; Costa, F.F.; Bischof, J.M.; Chou, P.M.; Bonaldo, M.D.F.; Vanin, E.F.; Soares, M.B. Identification of differentially expressed microRNAs in osteosarcoma. Sarcoma 2011, 2011, 732690. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Dan, G.; Shangguan, T.; Hao, H.; Tang, R.; Peng, K.; Zhao, J.; Sun, H.; Zou, Z. miR-198 represses the proliferation of HaCaT cells by targeting cyclin D2. Int. J. Mol. Sci. 2015, 16, 17018–17025. [Google Scholar] [CrossRef] [Green Version]
- Moura, J.; Sørensen, A.; Leal, E.C.; Svendsen, R.; Carvalho, L.; Willemoes, R.J.; Jørgensen, P.T.; Jenssen, H.; Wengel, J.; Dalgaard, L.T.; et al. microRNA-155 inhibition restores Fibroblast Growth Factor 7 expression in diabetic skin and decreases wound inflammation. Sci. Rep. 2019, 9, 5836. [Google Scholar] [CrossRef]
- Grieco, G.E.; Brusco, N.; Licata, G.; Nigi, L.; Formichi, C.; Dotta, F.; Sebastiani, G. Targeting microRNAs as a therapeutic strategy to reduce oxidative stress in diabetes. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, L.; Stone, R.C.; Stojadinovic, O.; Ramirez, H.; Pastar, I.; Maione, A.G.; Smith, A.; Yanez, V.; Veves, A.; Kirsner, R.S.; et al. Integrative analysis of miRNA and mRNA paired expression profiling of primary fibroblast derived from diabetic foot ulcers reveals multiple impaired cellular functions. Wound Repair Regen. 2016, 24, 943–953. [Google Scholar] [CrossRef] [PubMed]
- Curtale, G.; Rubino, M.; Locati, M. microRNAs as molecular switches in macrophage activation. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Liechty, C.; Hu, J.; Zhang, L.; Liechty, K.W.; Xu, J. Role of microRNA-21 and its underlying mechanisms in inflammatory responses in diabetic wounds. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Li, A.; Deng, J.; Yang, Y.; Dang, L.; Ye, Y.; Li, Y.; Zhang, W. MiR-21 attenuates lipopolysaccharide-induced lipid accumulation and inflammatory response: Potential role in cerebrovascular disease. Lipids Health Dis. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, B.; Cheng, Y.; Ma, L.; Zhang, C. MiR-21 regulates biological behavior through the PTEN/PI-3 K/Akt signaling pathway in human colorectal cancer cells. Int. J. Oncol. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Aurora, A.B.; Johnson, B.A.; Qi, X.; McAnally, J.; Hill, J.A.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. The endothelial-specific microrna mir-126 governs vascular integrity and angiogenesis. Dev. Cell 2008, 15, 261–271. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Sun, X.J.; Chen, J.; Hu, Z.W.; Wang, L.; Gu, D.M.; Wang, A. ping Increasing the miR-126 expression in the peripheral blood of patients with diabetic foot ulcers treated with maggot debridement therapy. J. Diabetes Complicat. 2017, 31, 241–244. [Google Scholar] [CrossRef]
- Zhang, D.; Li, Z.; Wang, Z.; Zeng, F.; Xiao, W.; Yu, A. MicroRNA-126: A promising biomarker for angiogenesis of diabetic wounds treated with negative pressure wound therapy. Diabetes Metab. Syndr. Obes. Targets Ther. 2019. [Google Scholar] [CrossRef] [Green Version]
- Harris, T.A.; Yamakuchi, M.; Ferlito, M.; Mendell, J.T.; Lowenstein, C.J. MicroRNA-126 regulates endothelial expression of vascular cell adhesion molecule 1. Proc. Natl. Acad. Sci. USA 2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milstone, D.S.; Ilyama, M.; Chen, M.; O’Donnell, P.; Davis, V.M.; Plutzky, J.; Brown, J.D.; Haldar, S.M.; Siu, A.; Lau, A.C.; et al. Differential role of an NF-κB transcriptional response element in endothelial versus intimal cell VCAM-1 expression. Circ. Res. 2015, 117, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Ke, Q.F.; Tao, S.C.; Guo, S.C.; Rui, B.Y.; Guo, Y.P. Fabrication of hydroxyapatite/chitosan composite hydrogels loaded with exosomes derived from miR-126-3p overexpressed synovial mesenchymal stem cells for diabetic chronic wound healing. J. Mater. Chem. B 2016, 4, 6830–6841. [Google Scholar] [CrossRef] [PubMed]
- Madhyastha, R.; Madhyastha, H.; Nakajima, Y.; Omura, S.; Maruyama, M. MicroRNA signature in diabetic wound healing: Promotive role of miR-21 in fibroblast migration. Int. Wound J. 2012, 9, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Nylander, E.; Coates, P.J.; Nylander, K. Effect of narrow-band ultraviolet B phototherapy on p63 and MicroRNA (miR-21 and miR-125b) expression in psoriatic epidermis. Acta Derm. Venereol. 2011, 91, 392–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.X.; Wang, J.; Guo, S.L.; Fan, K.J.; Li, J.; Wang, Y.L.; Teng, Y.; Yang, X.X. miR-21 promotes keratinocyte migration and re-epithelialization during wound Healing. Int. J. Biol. Sci. 2011, 7, 685–690. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.Y.; Kim, H.; Kwak, G.; Jo, S.D.; Cho, D.; Yang, Y.; Kwon, I.C.; Jeong, J.H.; Kim, S.H. Development of microRNA-21 mimic nanocarriers for the treatment of cutaneous wounds. Theranostics 2020, 10, 3240–3253. [Google Scholar] [CrossRef]
- Wang, T.; Feng, Y.; Sun, H.; Zhang, L.; Hao, L.; Shi, C.; Wang, J.; Li, R.; Ran, X.; Su, Y.; et al. MiR-21 regulates skin wound healing by targeting multiple aspects of the healing process. Am. J. Pathol. 2012, 181, 1911–1920. [Google Scholar] [CrossRef]
- Yang, L.; Zheng, Z.; Zhou, Q.; Bai, X.; Fan, L.; Yang, C.; Su, L.; Hu, D. miR-155 promotes cutaneous wound healing through enhanced keratinocytes migration by MMP-2. J. Mol. Histol. 2017, 48, 147–155. [Google Scholar] [CrossRef]
- Elgzyri, T.; Larsson, J.; Nyberg, P.; Thörne, J.; Eriksson, K.F.; Apelqvist, J. Reconstructive vascular surgery and the extent of tissue damage due to diabetic foot ulcers relates to risk of new ulceration in patients with PAD. J. Wound Care 2015, 24, 590–597. [Google Scholar] [CrossRef]
- Contreras, J.; Rao, D.S. microRNAs in inflammation and immune responses. Leukemia 2012, 26, 404–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, R.X.; Liu, R.F.; Zhao, X.C.; Jia, Y.R.; An, P.; Hao, Z.P.; Li, J.Q.; Li, X.H.; Yin, G.H.; Zhang, K.M. Increased miR-155-5p expression in dermal mesenchymal stem cells of psoriatic patients: Comparing the microRNA expression profile by microarray. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef]
- Qayum, A.A.; Paranjape, A.; Abebayehu, D.; Kolawole, E.M.; Haque, T.T.; McLeod, J.J.A.; Spence, A.J.; Caslin, H.L.; Taruselli, M.T.; Chumanevich, A.P.; et al. IL-10–induced miR-155 targets SOCS1 to enhance IgE-mediated mast cell function. J. Immunol. 2016, 196, 4457–4467. [Google Scholar] [CrossRef] [Green Version]
- Šahmatova, L.; Tankov, S.; Prans, E.; Aab, A.; Hermann, H.; Reemann, P.; Pihlap, M.; Karelson, M.; Abram, K.; Kisand, K.; et al. MicroRNA-155 is dysregulated in the skin of patients with vitiligo and inhibits melanogenesis-associated genes in melanocytes and keratinocytes. Acta Derm. Venereol. 2016, 96, 742–747. [Google Scholar] [CrossRef]
- Zhang, Y.; Mei, H.; Chang, X.; Chen, F.; Zhu, Y.; Han, X. Adipocyte-derived microvesicles from obese mice induce M 1 macrophage phenotype through secreted miR-155. J. Mol. Cell Biol. 2017, 8, 505–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Cui, Y.; Li, B.; Luo, X.; Li, B.; Tang, Y. miR-155 regulates high glucose-induced cardiac fibrosis via the TGF-β signaling pathway. Mol. Biosyst. 2017, 13, 215–224. [Google Scholar] [CrossRef] [PubMed]
- van Solingen, C.; Araldi, E.; Chamorro-jorganes, A.; Fernández-Hernando, C.; Suárez, Y.; Van Solingen, C.; Araldi, E.; Chamorro-jorganes, A.; Fern, C. Improved repair of dermal wounds in mice lacking microRNA-155. J. Cell. Mol. Med. 2014, 18, 1104–1112. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Kang, Y.; Sun, X.; Ni, P.; Wu, M.; Lu, S. MicroRNA-155 inhibition promoted wound healing in diabetic rats. Int. J. Low Extrem. Wounds 2017, 16, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, E.; Li, X.; Zhang, M.; Tang, Z.; He, L.; Lv, K. miR-155 contributes to Df1-induced asthma by increasing the proliferative response of Th cells via CTLA-4 downregulation. Cell. Immunol. 2017, 314, 1–9. [Google Scholar] [CrossRef]
- Landskroner-Eiger, S.; Qiu, C.; Perrotta, P.; Siragusa, M.; Lee, M.Y.; Ulrich, V.; Luciano, A.K.; Zhuang, Z.W.; Corti, F.; Simons, M.; et al. Endothelial miR-17~92 cluster negatively regulates arteriogenesis via miRNA-19 repression of WNT signaling. Proc. Natl. Acad. Sci. USA 2015, 112, 12812–12817. [Google Scholar] [CrossRef] [Green Version]
- Hinkel, R.; Penzkofer, D.; Zühlke, S.; Fischer, A.; Husada, W.; Xu, Q.F.; Baloch, E.; Van Rooij, E.; Zeiher, A.M.; Kupatt, C.; et al. Inhibition of microRNA~92a protects against ischemia/reperfusion injury in a large-animal model. Circulation 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, T.; Schäfer, F.; Müller, P.; Eming, S.A.; Heckel, A.; Dimmeler, S. Light-inducible antimiR~92a as a therapeutic strategy to promote skin repair in healing-impaired diabetic mice. Nat. Commun. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallant-Behm, C.L.; Piper, J.; Dickinson, B.A.; Dalby, C.M.; Pestano, L.A.; Jackson, A.L. A synthetic microRNA~92a inhibitor (MRG-110) accelerates angiogenesis and wound healing in diabetic and nondiabetic wounds. Wound Repair Regen. 2018, 26, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Davis-Dusenbery, B.N.; Chan, M.C.; Reno, K.E.; Weisman, A.S.; Layne, M.D.; Lagna, G.; Hata, A. Down-regulation of Krüppel-like Factor-4 (KLF4) by MicroRNA-143/145 Is Critical for Modulation of Vascular Smooth Muscle Cell Phenotype by Transforming Growth Factor-β and Bone Morphogenetic Protein 4. J. Biol. Chem. 2011, 286, 28097–28110. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Hu, C.; Cheng, J.; Chen, B.; Ke, Q.; Lv, Z.; Wu, J.; Zhou, Y. MicroRNA-145 suppresses hepatocellular carcinoma by targeting IRS1 and its downstream Akt signaling. Biochem. Biophys. Res. Commun. 2014. [Google Scholar] [CrossRef]
- Fernando, N.; Wong, J.H.C.; Das, S.; Dietrich, C.; Aggio-Bruce, R.; Cioanca, A.V.; Wooff, Y.; Chu-Tan, J.A.; Schumann, U.; Ngo, C.; et al. MicroRNA-223 regulates retinal function and inflammation in the healthy and degenerating retina. Front. Cell Dev. Biol. 2020, 8. [Google Scholar] [CrossRef]
- Dorhoi, A.; Iannaccone, M.; Farinacci, M.; Faé, K.C.; Schreiber, J.; Moura-Alves, P.; Nouailles, G.; Mollenkopf, H.J.; Oberbeck-Müller, D.; Jörg, S.; et al. MicroRNA-223 controls susceptibility to tuberculosis by regulating lung neutrophil recruitment. J. Clin. Investig. 2013, 123, 4836–4848. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Feng, D.; Li, M.; Gao, Y.; Ramirez, T.; Cao, H.; Kim, S.-J.J.; Yang, Y.; Cai, Y.; Ju, C.; et al. Hepatic mitochondrial DNA/Toll-like receptor 9/MicroRNA-223 forms a negative feedback loop to limit neutrophil overactivation and acetaminophen hepatotoxicity in mice. Hepatology 2017, 66, 220–234. [Google Scholar] [CrossRef] [Green Version]
- Kerckhove, M.; Tanaka, K.K.; Umehara, T.; Okamoto, M.; Kanematsu, S.; Hayashi, H.; Yano, H.; Nishiura, S.; Tooyama, S.; Matsubayashi, Y.; et al. Targeting miR-223 in neutrophils enhances the clearance of Staphylococcus aureus in infected wounds. EMBO Mol. Med. 2018, 10. [Google Scholar] [CrossRef]
- Ji, Q.; Xu, X.; Song, Q.; Xu, Y.; Tai, Y.; Goodman, S.B.; Bi, W.; Xu, M.; Jiao, S.; Maloney, W.J.; et al. miR-223-3p inhibits human osteosarcoma metastasis and progression by directly targeting CDH6. Mol. Ther. 2018, 26, 1299–1312. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Pal, A.S.; Hsu, A.Y.H.; Gurol, T.; Zhu, X.; Wirbisky-Hershberger, S.E.; Freeman, J.L.; Kasinski, A.L.; Deng, Q. MicroRNA-223 suppresses the canonical nf-κb pathway in basal keratinocytes to dampen neutrophilic inflammation. Cell Rep. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Morgan, M.J.; Choksi, S.; Zhang, Y.; Kim, Y.S.; Liu, Z.G. microRNAs modulate the noncanonical transcription factor NF-κB pathway by regulating expression of the kinase IKKα during macrophage differentiation. Nat. Immunol. 2010, 11, 799–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsui, S.; Ogata, Y. Effects of miR-223 on expression of IL-1β and IL-6 in human gingival fibroblasts. J. Oral Sci. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, C.P.; Leelahavanichkul, A. Over-expression of miR-223 induces M2 macrophage through glycolysis alteration and attenuates LPS-induced sepsis mouse model, the cell-based therapy in sepsis. PLoS ONE 2020. [Google Scholar] [CrossRef] [PubMed]
- Rousselle, P.; Braye, F.; Dayan, G. Re-epithelialization of adult skin wounds: Cellular mechanisms and therapeutic strategies. Adv. Drug Deliv. Rev. 2019, 146, 344–365. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Yang, W.; Shi, J.; Zhou, Y.; Yang, J.; Cui, Q.; Zhou, Y. Identification and Analysis of Human Sex-biased microRNAs. Genom. Proteom. Bioinforma 2018. [Google Scholar] [CrossRef] [PubMed]
- Noren Hooten, N.; Abdelmohsen, K.; Gorospe, M.; Ejiogu, N.; Zonderman, A.B.; Evans, M.K. microRNA expression patterns reveal differential expression of target genes with age. PLoS ONE 2010. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Yuan, Y.; Cho, J.H.; McClarty, S.; Baxter, D.; Galas, D.J. Comparing the MicroRNA spectrum between serum and plasma. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Vasu, S.; Kumano, K.; Darden, C.M.; Rahman, I.; Lawrence, M.C.; Naziruddin, B. MicroRNA signatures as future biomarkers for diagnosis of diabetes states. Cells 2019. [Google Scholar] [CrossRef] [Green Version]
- Mestdagh, P.; Hartmann, N.; Baeriswyl, L.; Andreasen, D.; Bernard, N.; Chen, C.; Cheo, D.; D’Andrade, P.; DeMayo, M.; Dennis, L.; et al. Evaluation of quantitative miRNA expression platforms in the microRNA quality control (miRQC) study. Nat. Methods 2014. [Google Scholar] [CrossRef]
- MaClellan, S.A.; Macaulay, C.; Lam, S.; Garnis, C. Pre-profiling factors influencing serum microRNA levels. BMC Clin. Pathol. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Xu, Y.; Shu, B.; Wang, P.; Tang, J.; Chen, L.; Qi, S.; Liu, X.; Xie, J. Quantification of the differential expression levels of microRNA-203 in different degrees of diabetic foot. Int. J. Clin. Exp. Pathol. 2015, 8, 13416–13420. [Google Scholar]
- Lin, C.J.; Lan, Y.M.; Ou, M.Q.; Ji, L.Q.; Lin, S.D. Expression of miR-217 and HIF-1α/VEGF pathway in patients with diabetic foot ulcer and its effect on angiogenesis of diabetic foot ulcer rats. J. Endocrinol. Investig. 2019, 42, 1307–1317. [Google Scholar] [CrossRef] [PubMed]
- La Rocca, G.; Badin, M.; Shi, B.; Xu, S.Q.; Deangelis, T.; Sepp-Lorenzino, L.; Baserga, R. Mechanism of growth inhibition by microRNA 145: The role of the IGF-I receptor signaling pathway. J. Cell. Physiol. 2009. [Google Scholar] [CrossRef] [PubMed]
- Pasquier, J.; Ramachandran, V.; Abu-Qaoud, M.R.; Thomas, B.; Benurwar, M.J.; Chidiac, O.; Hoarau-Véchot, J.; Robay, A.; Fakhro, K.; Menzies, R.A.; et al. Differentially expressed circulating microRNAs in the development of acute diabetic Charcot foot. Epigenomics 2018, 10, 1267–1278. [Google Scholar] [CrossRef] [Green Version]
- Li, J.Y.; Cheng, B.; Wang, X.F.; Wang, Z.J.; Zhang, H.M.; Liu, S.F.; Chen, L.S.; Huang, W.J.; Liu, J.; Deng, A.P. Circulating MicroRNA-4739 may be a potential biomarker of critical limb ischemia in patients with diabetes. BioMed Res. Int. 2018, 2018. [Google Scholar] [CrossRef] [Green Version]
- Spinetti, G.; Fortunato, O.; Caporali, A.; Shantikumar, S.; Marchetti, M.; Meloni, M.; Descamps, B.; Floris, I.; Sangalli, E.; Vono, R.; et al. MicroRNA-15a and MicroRNA-16 impair human circulating proangiogenic cell functions and are increased in the proangiogenic cells and serum of patients with critical limb ischemia. Circ. Res. 2013, 112, 335–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witherel, C.E.; Abebayehu, D.; Barker, T.H.; Spiller, K.L. Macrophage and fibroblast interactions in biomaterial-mediated fibrosis. Adv. Healthc. Mater. 2019, 8, e1801451. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Ren, Y.; Liu, P.; Ren, Y.; Wang, D. Expression of TGF-β1 and miRNA-145 in patients with diabetic foot ulcers. Exp. Ther. Med. 2016, 11, 2011–2014. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Yang, C.; Wang, X.Y.; Zhou, L.Y.; Lao, G.J.; Liu, D.; Wang, C.; Hu, M.D.; Zeng, T.T.; Yan, L.; et al. MicroRNA-129 and -335 promote diabetic wound healing by inhibiting Sp1-mediated MMP-9 expression. Diabetes 2018, 67, 1627–1638. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, H.A.; Liang, L.; Pastar, I.; Rosa, A.M.; Stojadinovic, O.; Zwick, T.G.; Kirsner, R.S.; Maione, A.G.; Garlick, J.A.; Tomic-Canic, M. Comparative genomic, microRNA, and tissue analyses reveal subtle differences between non-diabetic and diabetic foot skin. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Than, U.T.T.; Guanzon, D.; Broadbent, J.A.; Leavesley, D.I.; Salomon, C.; Parker, T.J. Differential expression of keratinocyte-derived extracellular vesicle mirnas discriminate exosomes from apoptotic bodies and microvesicles. Front. Endocrinol. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, E.W.; Seo, M.K.; Woo, E.Y.; Kim, S.H.; Park, E.J.; Kim, S. Exosomes from human adipose-derived stem cells promote proliferation and migration of skin fibroblasts. Exp. Dermatol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Geiger, A.; Walker, A.; Nissen, E. Human fibrocyte-derived exosomes accelerate wound healing in genetically diabetic mice. Biochem. Biophys. Res. Commun. 2015. [Google Scholar] [CrossRef]
- Zhao, B.; Zhang, Y.; Han, S.; Zhang, W.; Zhou, Q.; Guan, H.; Liu, J.; Shi, J.; Su, L.; Hu, D. Exosomes derived from human amniotic epithelial cells accelerate wound healing and inhibit scar formation. J. Mol. Histol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jiang, C.; Zhao, J. Human endothelial progenitor cells-derived exosomes accelerate cutaneous wound healing in diabetic rats by promoting endothelial function. J. Diabetes Complicat. 2016. [Google Scholar] [CrossRef]
- Freeman, D.W.; Noren Hooten, N.; Eitan, E.; Green, J.; Mode, N.A.; Bodogai, M.; Zhang, Y.; Lehrmann, E.; Zonderman, A.B.; Biragyn, A.; et al. Altered extracellular vesicle concentration, cargo, and function in diabetes. Diabetes 2018. [Google Scholar] [CrossRef] [Green Version]
- Leszczynska, A.; Kulkarni, M.; Ljubimov, A.V.; Saghizadeh, M. Exosomes from normal and diabetic human corneolimbal keratocytes differentially regulate migration, proliferation and marker expression of limbal epithelial cells. Sci. Rep. 2018. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Zhang, C.; Liu, L.; Xi, A.; Chen, B.; Li, Y.; Du, J. Macrophage-Derived mir-155-containing exosomes suppress fibroblast proliferation and promote fibroblast inflammation during cardiac injury. Mol. Ther. 2017. [Google Scholar] [CrossRef] [Green Version]
- Henriques-Antunes, H.; Cardoso, R.M.S.; Zonari, A.; Correia, J.; Leal, E.C.; Jiménez-Balsa, A.; Lino, M.M.; Barradas, A.; Kostic, I.; Gomes, C.; et al. The kinetics of small extracellular vesicle delivery impacts skin tissue regeneration. ACS Nano 2019. [Google Scholar] [CrossRef]
- Jansen, F.; Yang, X.; Hoelscher, M.; Cattelan, A.; Schmitz, T.; Proebsting, S.; Wenzel, D.; Vosen, S.; Franklin, B.S.; Fleischmann, B.K.; et al. Endothelial microparticle-mediated transfer of microRNA-126 promotes vascular endothelial cell repair via spred1 and is abrogated in glucose-damaged endothelial microparticles. Circulation 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Bai, S.; Cao, Y.; Liu, L.; Fang, Y.; Du, J.; Luo, L.; Chen, M.; Shen, B.; Zhang, Q. miRNA-221-3p in endothelial progenitor cell-derived exosomes accelerates skin wound healing in diabetic mice. Diabetes Metab. Syndr. Obes. Targets Ther. 2020. [Google Scholar] [CrossRef] [Green Version]
- Shyu, K.G.; Wang, B.W.; Pan, C.M.; Fang, W.J.; Lin, C.M. Hyperbaric oxygen boosts long noncoding RNA MALAT1 exosome secretion to suppress microRNA-92a expression in therapeutic angiogenesis. Int. J. Cardiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Zhu, C.; Jia, J.; Hao, X.Y.; Yu, X.Y.; Liu, X.Y.; Shu, M.G. ADSC-Exos containing MALAT1 promotes wound healing by targeting miR-124 through activating Wnt/β-catenin pathway. Biosci. Rep. 2020. [Google Scholar] [CrossRef] [PubMed]
- Abdulle, L.E.; Hao, J.L.; Pant, O.P.; Liu, X.F.; Zhou, D.D.; Gao, Y.; Suwal, A.; Lu, C.W. MALAT1 as a diagnostic and therapeutic target in diabetes-related complications: A promising long-noncoding RNA. Int. J. Med. Sci. 2019, 16, 548–555. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Luan, S.; Chen, J.; Zhou, Y.; Wang, T.; Li, Z.; Fu, Y.; Zhai, A.; Bi, C. The MSC-derived exosomal lncRNA H19 promotes wound healing in diabetic foot ulcers by upregulating PTEN via MicroRNA-152-3p. Mol. Ther. Nucleic Acids 2020. [Google Scholar] [CrossRef] [PubMed]
- Ti, D.; Hao, H.; Tong, C.; Liu, J.; Dong, L.; Zheng, J.; Zhao, Y.; Liu, H.; Fu, X.; Han, W. LPS-preconditioned mesenchymal stromal cells modify macrophage polarization for resolution of chronic inflammation via exosome-shuttled let-7b. J. Transl. Med. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Zhang, L.; Wang, S.; Han, Q.; Zhao, R.C. Exosomes secreted by mesenchymal stem cells promote endothelial cell angiogenesis by transferring miR-125a. J. Cell Sci. 2016. [Google Scholar] [CrossRef] [Green Version]
- Xueya, Z.; Yamei, L.; Sha, C.; Dan, C.; Hong, S.; Xingyu, Y.; Weiwei, C. Exosomal encapsulation of miR-125a-5p inhibited trophoblast cell migration and proliferation by regulating the expression of VEGFA in preeclampsia. Biochem. Biophys. Res. Commun. 2020. [Google Scholar] [CrossRef]
- Shi, R.; Jin, Y.; Hu, W.; Lian, W.; Cao, C.; Han, S.; Zhao, S.; Yuan, H.; Yang, X.; Shi, J.; et al. Exosomes derived from mmu_circ_0000250-modified adipose-derived mesenchymal stem cells promote wound healing in diabetic mice by inducing miR-128-3p/SIRT1-mediated autophagy. Am. J. Physiol. Cell Physiol. 2020. [Google Scholar] [CrossRef]
- Xiong, Y.; Chen, L.; Yan, C.; Zhou, W.; Endo, Y.; Liu, J.; Hu, L.; Hu, Y.; Mi, B.; Liu, G. Circulating Exosomal miR-20b-5p inhibition restores wnt9b signaling and reverses diabetes-associated impaired wound healing. Small 2020. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; Chen, L.; Yu, T.; Yan, C.; Zhou, W.; Cao, F.; You, X.; Zhang, Y.; Sun, Y.; Liu, J.; et al. Inhibition of circulating exosomal microRNA-15a-3p accelerates diabetic wound repair. Aging 2020. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Luo, L.; Bai, X.; Shen, K.; Liu, K.; Wang, J.; Hu, D. Highly-expressed micoRNA-21 in adipose derived stem cell exosomes can enhance the migration and proliferation of the HaCaT cells by increasing the MMP-9 expression through the PI3K/AKT pathway. Arch. Biochem. Biophys. 2020. [Google Scholar] [CrossRef]
- Hu, Y.; Rao, S.S.; Wang, Z.X.; Cao, J.; Tan, Y.J.; Luo, J.; Li, H.M.; Zhang, W.S.; Chen, C.Y.; Xie, H. Exosomes from human umbilical cord blood accelerate cutaneous wound healing through miR-21-3p-mediated promotion of angiogenesis and fibroblast function. Theranostics 2018. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhao, H.; Chen, W.; Huang, P.; Bi, J. Human keratinocyte-derived microvesicle miRNA-21 promotes skin wound healing in diabetic rats through facilitating fibroblast function and angiogenesis. Int. J. Biochem. Cell Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.; Deng, J.; Chen, Y.; Wang, Y.; Liu, B.; Liu, J. Engineered human adipose stem-cell-derived exosomes loaded with miR-21-5p to promote diabetic cutaneous wound healing. Mol. Pharm. 2020. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Wang, B.; Jiang, C.; Li, R.; Zhao, J. Endothelial progenitor cell-derived exosomes facilitate vascular endothelial cell repair through shuttling miR-21-5p to modulate Thrombospondin-1 expression. Clin. Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Zhang, T.; Wei, Y.; Cai, W.J.; Feng, G.; Chang, X.Y.; Sun, K. Epigenetic regulation of microRNA-375 and its role as DNA epigenetic marker of type 2 diabetes mellitus in Chinese Han population. Int. J. Clin. Exp. Pathol. 2017, 10, 11986–11994. [Google Scholar] [PubMed]
- Sookoian, S.; Gianotti, T.F.; Burgueño, A.L.; Pirola, C.J. Insulin resistance and epigenetic regulation: Insights from human studies and prospects for future research. Biomol. Concepts 2011, 2, 445–457. [Google Scholar] [CrossRef]
- Van der Kwast, R.V.C.T.; Parma, L.; van der Bent, M.L.; van Ingen, E.; Baganha, F.; Peters, H.A.B.; Goossens, E.A.C.; Simons, K.H.; Palmen, M.; de Vries, M.R.; et al. Adenosine-to-inosine editing of vasoactive microRNAs alters their targetome and function in ischemia. Mol. Ther. Nucleic Acids 2020, 21, 932–953. [Google Scholar] [CrossRef]



| Cell Type | Function | References |
|---|---|---|
Keratinocytes | Under normal conditions, the main function of keratinocytes (KCs) is to form the protective barrier of the skin. As a response to the injury, keratinocytes secrete a vast yield of soluble fibrinogenic and angiogenic growth factors such as TGFα, TGFβ, VEGF, EGF, and KGF that prompt the regeneration in bordering tissue, during the wound healing process. | [38,39] |
Platelets | Platelets are one of the first cell types to respond to injury. They are a large source of growth factors like platelet-derived growth factor (PDGF) and transforming growth factor beta (TGF-β) in the early wound. | [40,41] |
Dendritic cells | Dendritic cells (DCs) are antigen-presenting cells also known as messengers between the innate and the adaptive mammalian immune response during each phase of wound healing. | [42,43] |
Neutrophils | Neutrophils are in the first line of response of the innate immune response, producing an immediate and potent response against invading and harmful agents. Neutrophils are highly motile and abundantly recruited and amplified in response to cytokines released from damaged and necrotic cells after tissue injury. | [44,45] |
Endothelial cells | Endothelial cells (ECs) respond to the wound healing signals released by keratinocytes and fibroblasts, initiating angiogenesis, where they have a special role in augmenting the growth and survival of newly formed tissue. | [46] |
Myofibroblasts and fibroblasts | Myofibroblasts are typically activated fibroblasts that contribute to wound healing by generating extracellular matrix. They also propagate a contractile strength to the wound edges during wound contraction. Myofibroblasts differentiation is frequently induced by endothelin-1, TGFβ, and cellular fibronectin. Fibroblasts are one of the key players in the wound contraction related events like resolving the fibrin clot, and establishing the extra cellular matrix (ECM) and collagen deposition. | [47] |
Lymphocytes | Lymphocytes play an important regulatory role in wound healing and scar formation. | [48] |
Macrophages | Two fractions present in the tissues. Tissue resident macrophages are constitutively present, while the recruited macrophages appear at the wound site and play an important role in clearing the matrix, cell debris and microorganisms. They coordinate tissue repair. | [49,50] |
NK-cells | Natural killer (NK) cells regulate the inflammatory phase in wound repair, as well as the later stages of wound healing (re-epithelialization, angiogenesis, granulation tissue formation, and the remodeling phase). | [51] |
B-cells | B cells are part of the humoral branch of the immune system. One of the studies shows that the infiltration of B-cells into the wound is mediated through interleukin 10 (IL-10). | [52] |
T-cells | T-cells localized at the epidermis and may play regulatory roles in skin tissue homeostasis and repair. | [53,54] |
Cutaneous innervation | Skin physiological and pathophysiological properties rely on the sensory neuronal network (consisting of various epidermal and dermal receptors; and sensory autonomic nerve fibers). Sensory innervation coordinates skin responses by releasing different kinds of neuropeptides, which in turn activate skin cells during the immune response. | [55,56,57,58] |
| Wound Healing | ||||||||
|---|---|---|---|---|---|---|---|---|
| Inflammation | Proliferation | Remodeling | ||||||
| microRNA | Target | Reference | microRNA | Target | Reference | microRNA | Target | Reference |
| miR-132 | NFκB, TNFα, STAT3, IRAK4 | [121,122,123,124] | miR-210 | E2F3 | [125,126,127,128] | miR-29 | COL 1-3, TGF1β, SMAD | [129,130,131,132,133,134,135] |
| miR-146a | TRAF6, IRAK1, IRAK2 | [136,137,138,139] | miR-200 | MMP2, VEGF2 ZEB1 SIP1 | [140,141,142,143,144] | miR-198 | FSTL1, CCND2 | [145,146,147,148,149,150] |
| miR-21 | NOX2, IL1β, iNOS, IL6, TNFα, PTEN | [151,152,153,154,155,156,157] | miR-126 | SPRED1, PIK3R2, VCAM1 | [158,159,160,161,162,163] | miR-21 | TGF1β | [164,165,166,167,168] |
| miR-155 | TLA4, SOCS1, SHIP, FGF7 | [151,169,170,171,172,173,174,175,176,177,178,179] | miR-17~92 | TGF1β, SMAD1, ITGA5 | [180,181,182,183] | miR-143/miR-145 | IRS1, PDGFD, αSMA | [184,185 |
| miR-223 | MKP5, IKKα | [58,186,187,188,189,190,191,192,193,194] | ||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petkovic, M.; Sørensen, A.E.; Leal, E.C.; Carvalho, E.; Dalgaard, L.T. Mechanistic Actions of microRNAs in Diabetic Wound Healing. Cells 2020, 9, 2228. https://doi.org/10.3390/cells9102228
Petkovic M, Sørensen AE, Leal EC, Carvalho E, Dalgaard LT. Mechanistic Actions of microRNAs in Diabetic Wound Healing. Cells. 2020; 9(10):2228. https://doi.org/10.3390/cells9102228
Chicago/Turabian StylePetkovic, Marija, Anja Elaine Sørensen, Ermelindo Carreira Leal, Eugenia Carvalho, and Louise Torp Dalgaard. 2020. "Mechanistic Actions of microRNAs in Diabetic Wound Healing" Cells 9, no. 10: 2228. https://doi.org/10.3390/cells9102228
APA StylePetkovic, M., Sørensen, A. E., Leal, E. C., Carvalho, E., & Dalgaard, L. T. (2020). Mechanistic Actions of microRNAs in Diabetic Wound Healing. Cells, 9(10), 2228. https://doi.org/10.3390/cells9102228