The Role of Bone Morphogenetic Protein 7 (BMP-7) in Inflammation in Heart Diseases

Abstract
:1. Introduction
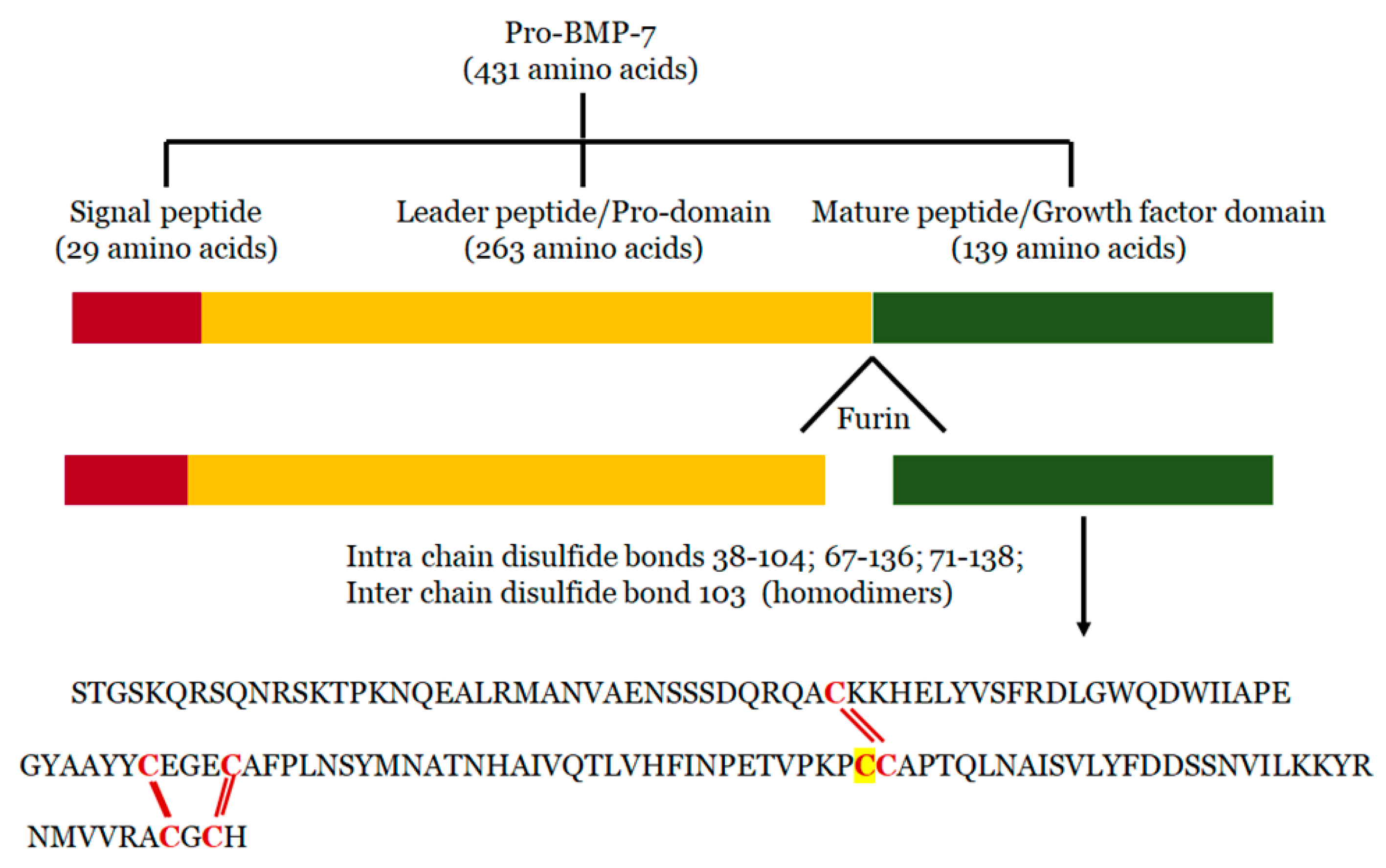
2. Structure of BMP-7
3. Mechanisms of BMP-7
4. Inhibitors of BMP-7
5. Regulators of BMP-7
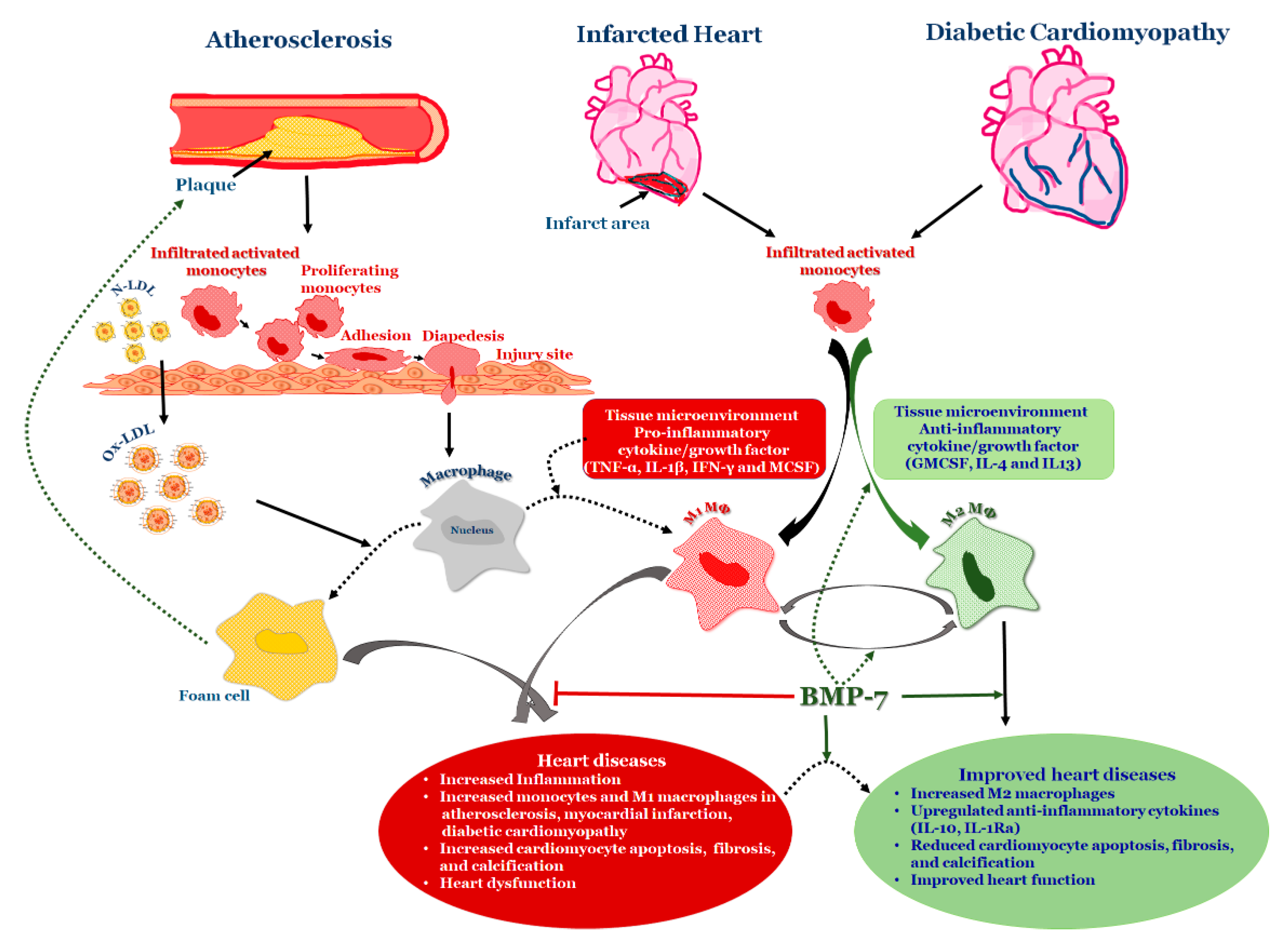
6. BMP-7 as an Anti-Inflammatory Agent in Atherosclerosis
7. BMP-7 as an Anti-Calcifying Agent
8. BMP-7 Inhibits Inflammation and Adverse Remodeling in the Infarcted Heart
9. BMP-7 Ameliorates Diabetic Cardiomyopathy
10. BMP-7 Differentiates Monocytes into M2 Macrophages in Heart Diseases
11. Conclusions and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALP | alkaline phosphatase |
| Akt | RAC-alpha serine/threonine-protein kinase |
| ATF2 | activating transcription factor 2 |
| Bcl-2 | B-cell lymphoma 2 |
| BMP | bone morphogenetic protein |
| BMPR | bone morphogenetic protein receptor |
| BRAM1 | bone morphogenetic protein receptor associated molecule 1 |
| CCR2 | C-C chemokine receptor type 2 |
| CE | cholesteryl ester |
| CTGF | connective tissue growth factor |
| CVD | cardiovascular disease |
| CX3CR1 | C-X3-C motif chemokine receptor |
| DC | dendritic cells |
| ECM | extracellular matrix |
| ERK | extracellular signal-regulated Kinase |
| GMCSF | granulocyte macrophage colony stimulating factor |
| HSC | hematopoietic stem cell |
| IFN-γ | interferon gamma |
| iNOS | inducible nitric oxide synthase |
| JNK | c-Jun-N terminal Kinase |
| MAPK | mitogen-activated protein kinases |
| MCP-1 | monocyte chemotactic protein |
| MCSF | macrophage colony stimulating factor |
| MMP | matrix metalloproteinase |
| MNC | mononuclear cells |
| MSC | mesenchymal stem cells |
| mTOR | mammalian target of rapamycin |
| MyoD | myoblast determination protein 1 |
| NFkB | nuclear factor kappa-light-chain-enhancer of activated B |
| OCN | osteocalcin |
| OPN | osteopontin |
| OSX | osterix |
| PI3 | phosphatidylinositol 3 |
| PK | protein kinase |
| PTEN | phosphatase and tensin homolog |
| RON | reactive nitrogen species |
| ROS | reactive oxygen species |
| Runx2 | run-related transcription factor 2 |
| SR | scavenger receptor |
| T2DM | type 2 diabetes |
| TAB1 | TAK1 binding protein |
| TAK1 | TGF-beta activated kinase 1 |
| TF | tissue factor |
| TGF-β1 | transforming growth factor beta1 |
| TNF-α | tumor necrosis factor alpha |
| TRAIL | TNF-related apoptosis-inducing ligand |
| VSMC | Vascular smooth muscle cells |
| XIAP | X-linked inhibitor of apoptosis protein |
References
- Mazerbourg, S.; Hsueh, A.J. Genomic analyses facilitate identification of receptors and signalling pathways for growth differentiation factor 9 and related orphan bone morphogenetic protein/growth differentiation factor ligands. Hum. Reprod. Update 2006, 12, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Von Bubnoff, A.; Cho, K.W. Intracellular BMP signaling regulation in vertebrates: Pathway or network? Dev. Biol. 2001, 239, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Bragdon, B.; Moseychuk, O.; Saldanha, S.; King, D.; Julian, J.; Nohe, A. Bone morphogenetic proteins: A critical review. Cell. Signal. 2011, 23, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Miyazono, K.; Maeda, S.; Imamura, T. BMP receptor signaling: Transcriptional targets, regulation of signals, and signaling cross-talk. Cytokine Growth Factor Rev. 2005, 16, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.T.; Xiang, L.X.; Shao, J.Z. Bone morphogenetic protein. Biochem. Biophys. Res. Commun. 2007, 362, 550–553. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Ji, X.; Harris, M.A.; Feng, J.Q.; Karsenty, G.; Celeste, A.J.; Rosen, V.; Mundy, G.R.; Harris, S.E. Differential Roles for Bone Morphogenetic Protein (BMP) Receptor Type IB and IA in Differentiation and Specification of Mesenchymal Precursor Cells to Osteoblast and Adipocyte Lineages. J. Cell Biol. 1998, 142, 295–305. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Deng, C.; Li, Y.P. TGF-beta and BMP signaling in osteoblast differentiation and bone formation. Int. J. Biol. Sci. 2012, 8, 272–288. [Google Scholar] [CrossRef] [Green Version]
- Takada, I.; Kouzmenko, A.P.; Kato, S. PPAR-gamma Signaling Crosstalk in Mesenchymal Stem Cells. PPAR Res. 2010, 2010. [Google Scholar] [CrossRef] [Green Version]
- Hata, K.; Nishimura, R.; Ikeda, F.; Yamashita, K.; Matsubara, T.; Nokubi, T.; Yoneda, T. Differential roles of Smad1 and p38 kinase in regulation of peroxisome proliferator-activating receptor gamma during bone morphogenetic protein 2-induced adipogenesis. Mol. Biol. Cell. 2003, 14, 545–555. [Google Scholar] [CrossRef] [Green Version]
- Kang, Q.; Song, W.X.; Luo, Q.; Tang, N.; Luo, J.; Luo, X.; Chen, J.; Bi, Y.; He, B.C.; Park, J.K.; et al. A comprehensive analysis of the dual roles of BMPs in regulating adipogenic and osteogenic differentiation of mesenchymal progenitor cells. Stem Cells Dev. 2009, 18, 545–559. [Google Scholar] [CrossRef] [Green Version]
- Dorman, L.J.; Tucci, M.; Benghuzzi, H. In vitro effects of bmp-2, bmp-7, and bmp-13 on proliferation and differentation of mouse mesenchymal stem cells. Biomed. Sci. Instrum. 2012, 48, 81–87. [Google Scholar] [PubMed]
- Reid, J.; Gilmour, H.M.; Holt, S. Primary non-specific ulcer of the small intestine. J. R. Coll. Surg. Edinb. 1982, 27, 228–232. [Google Scholar] [PubMed]
- Varkey, M.; Kucharski, C.; Haque, T.; Sebald, W.; Uludag, H. In vitro osteogenic response of rat bone marrow cells to bFGF and BMP-2 treatments. Clin. Orthop. Relat. Res. 2006, 443, 113–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granjeiro, J.M.; Oliveira, R.C.; Bustos-Valenzuela, J.C.; Sogayar, M.C.; Taga, R. Bone morphogenetic proteins: From structure to clinical use. Braz. J. Med. Biol. Res. Rev. Bras. Pesqui. Med. Biol. 2005, 38, 1463–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, S.W.; Tang, Y.; Li, X.; Liu, Y.; Zhang, Y.Y.; Huang, H.Y.; Xue, R.D.; Yu, H.Y.; Guo, L.; Gao, H.D.; et al. BMP4-mediated brown fat-like changes in white adipose tissue alter glucose and energy homeostasis. Proc. Natl. Acad. Sci. USA 2013, 110, E798–E807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalluri, R.; Zeisberg, M. Fibroblasts in cancer. Nat. Rev. Cancer 2006, 6, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Friedrichs, M.; Wirsdoerfer, F.; Flohe, S.B.; Schneider, S.; Wuelling, M.; Vortkamp, A. BMP signaling balances proliferation and differentiation of muscle satellite cell descendants. BMC Cell Biol. 2011, 12, 26. [Google Scholar] [CrossRef] [Green Version]
- Segklia, A.; Seuntjens, E.; Elkouris, M.; Tsalavos, S.; Stappers, E.; Mitsiadis, T.A.; Huylebroeck, D.; Remboutsika, E.; Graf, D. Bmp7 regulates the survival, proliferation, and neurogenic properties of neural progenitor cells during corticogenesis in the mouse. PLoS ONE 2012, 7, e34088. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Bradley, A. Mice deficient for BMP2 are nonviable and have defects in amnion/chorion and cardiac development. Development 1996, 122, 2977–2986. [Google Scholar]
- Winnier, G.; Blessing, M.; Labosky, P.A.; Hogan, B.L. Bone morphogenetic protein-4 is required for mesoderm formation and patterning in the mouse. Genes Dev. 1995, 9, 2105–2116. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Shi, S.; Acosta, L.; Li, W.; Lu, J.; Bao, S.; Chen, Z.; Yang, Z.; Schneider, M.D.; Chien, K.R.; et al. BMP10 is essential for maintaining cardiac growth during murine cardiogenesis. Development 2004, 131, 2219–2231. [Google Scholar] [CrossRef] [Green Version]
- Dudley, A.T.; Lyons, K.M.; Robertson, E.J. A requirement for bone morphogenetic protein-7 during development of the mammalian kidney and eye. Genes Dev. 1995, 9, 2795–2807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McPherron, A.C.; Lawler, A.M.; Lee, S.J. Regulation of anterior/posterior patterning of the axial skeleton by growth/differentiation factor 11. Nat. Genet. 1999, 22, 260–264. [Google Scholar] [CrossRef]
- Komatsu, Y.; Scott, G.; Nagy, A.; Kaartinen, V.; Mishina, Y. BMP type I receptor ALK2 is essential for proper patterning at late gastrulation during mouse embryogenesis. Dev. Dyn. 2007, 236, 512–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beppu, H.; Kawabata, M.; Hamamoto, T.; Chytil, A.; Minowa, O.; Noda, T.; Miyazono, K. BMP type II receptor is required for gastrulation and early development of mouse embryos. Dev. Biol. 2000, 221, 249–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishina, Y.; Suzuki, A.; Ueno, N.; Behringer, R.R. Bmpr encodes a type I bone morphogenetic protein receptor that is essential for gastrulation during mouse embryogenesis. Genes Dev. 1995, 9, 3027–3037. [Google Scholar] [CrossRef] [Green Version]
- Sirard, C.; de la Pompa, J.L.; Elia, A.; Itie, A.; Mirtsos, C.; Cheung, A.; Hahn, S.; Wakeham, A.; Schwartz, L.; Kern, S.E.; et al. The tumor suppressor gene Smad4/Dpc4 is required for gastrulation and later for anterior development of the mouse embryo. Genes Dev. 1998, 12, 107–119. [Google Scholar] [CrossRef]
- Chang, H.; Huylebroeck, D.; Verschueren, K.; Guo, Q.; Matzuk, M.M.; Zwijsen, A. Smad5 knockout mice die at mid-gestation due to multiple embryonic and extraembryonic defects. Development 1999, 126, 1631–1642. [Google Scholar]
- Lechleider, R.J.; Ryan, J.L.; Garrett, L.; Eng, C.; Deng, C.; Wynshaw-Boris, A.; Roberts, A.B. Targeted mutagenesis of Smad1 reveals an essential role in chorioallantoic fusion. Dev. Biol. 2001, 240, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Chen, H.; Zheng, D.; Kuang, C.; Fang, H.; Zou, B.; Zhu, W.; Bu, G.; Jin, T.; Wang, Z.; et al. Smad7 is required for the development and function of the heart. J. Biol. Chem. 2009, 284, 292–300. [Google Scholar] [CrossRef] [Green Version]
- Goldman, D.C.; Bailey, A.S.; Pfaffle, D.L.; Al Masri, A.; Christian, J.L.; Fleming, W.H. BMP4 regulates the hematopoietic stem cell niche. Blood 2009, 114, 4393–4401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Lu, M.F.; Schwartz, R.J.; Martin, J.F. Bmp2 is essential for cardiac cushion epithelial-mesenchymal transition and myocardial patterning. Development 2005, 132, 5601–5611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivera-Feliciano, J.; Tabin, C.J. Bmp2 instructs cardiac progenitors to form the heart-valve-inducing field. Dev. Biol. 2006, 295, 580–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimitriou, R.; Tsiridis, E.; Giannoudis, P.V. Current concepts of molecular aspects of bone healing. Injury 2005, 36, 1392–1404. [Google Scholar] [CrossRef] [PubMed]
- Carreira, A.C.; Alves, G.G.; Zambuzzi, W.F.; Sogayar, M.C.; Granjeiro, J.M. Bone Morphogenetic Proteins: Structure, biological function and therapeutic applications. Arch. Biochem. Biophys. 2014, 561, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Choe, S. BMPs and their clinical potentials. BMB Rep. 2011, 44, 619–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddi, A.H. Bone morphogenetic proteins: From basic science to clinical applications. J. Bone Jt. Surg. Am. Vol. 2001, 83. [Google Scholar] [CrossRef]
- Geesink, R.G.; Hoefnagels, N.H.; Bulstra, S.K. Osteogenic activity of OP-1 bone morphogenetic protein (BMP-7) in a human fibular defect. J. Bone Jt. Surg. British Vol. 1999, 81, 710–718. [Google Scholar] [CrossRef]
- Friedlaender, G.E.; Perry, C.R.; Cole, J.D.; Cook, S.D.; Cierny, G.; Muschler, G.F.; Zych, G.A.; Calhoun, J.H.; LaForte, A.J.; Yin, S. Osteogenic protein-1 (bone morphogenetic protein-7) in the treatment of tibial nonunions. J. Bone Jt. Surg. Am. Vol. 2001, 83, 151–158. [Google Scholar] [CrossRef]
- Cummings, S.R.; Melton, L.J. Epidemiology and outcomes of osteoporotic fractures. Lancet 2002, 359, 1761–1767. [Google Scholar] [CrossRef]
- Giannoudis, P.; Tzioupis, C.; Almalki, T.; Buckley, R. Fracture healing in osteoporotic fractures: Is it really different? A basic science perspective. Injury 2007, 38, 90–99. [Google Scholar] [CrossRef]
- Qaseem, A.; Snow, V.; Shekelle, P.; Hopkins, R., Jr.; Forciea, M.A.; Owens, D.K. Pharmacologic treatment of low bone density or osteoporosis to prevent fractures: A clinical practice guideline from the American College of Physicians. Ann. Intern. Med. 2008, 149, 404–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.; Zhao, M.; Mundy, G.R. Bone morphogenetic proteins. Growth Factors 2004, 22, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Nakase, T.; Yoshikawa, H. Potential roles of bone morphogenetic proteins (BMPs) in skeletal repair and regeneration. J. Bone Miner. Metab. 2006, 24, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Ten Dijke, P.; Yamashita, H.; Sampath, T.K.; Reddi, A.H.; Estevez, M.; Riddle, D.L.; Ichijo, H.; Heldin, C.H.; Miyazono, K. Identification of type I receptors for osteogenic protein-1 and bone morphogenetic protein-4. J. Biol. Chem. 1994, 269, 16985–16988. [Google Scholar]
- Hahn, G.V.; Cohen, R.B.; Wozney, J.M.; Levitz, C.L.; Shore, E.M.; Zasloff, M.A.; Kaplan, F.S. A bone morphogenetic protein subfamily: Chromosomal localization of human genes for BMP5, BMP6, and BMP7. Genomics 1992, 14, 759–762. [Google Scholar] [CrossRef]
- Nonner, D.; Barrett, E.F.; Kaplan, P.; Barrett, J.N. Bone morphogenetic proteins (BMP6 and BMP7) enhance the protective effect of neurotrophins on cultured septal cholinergic neurons during hypoglycemia. J. Neurochem. 2001, 77, 691–699. [Google Scholar] [CrossRef]
- Solloway, M.J.; Dudley, A.T.; Bikoff, E.K.; Lyons, K.M.; Hogan, B.L.; Robertson, E.J. Mice lacking Bmp6 function. Dev. Genet. 1998, 22, 321–339. [Google Scholar] [CrossRef]
- Zhang, Y.; Ge, G.; Greenspan, D.S. Inhibition of Bone Morphogenetic Protein 1 by Native and Altered Forms of α2-Macroglobulin. J. Biol. Chem. 2006, 281, 39096–39104. [Google Scholar] [CrossRef] [Green Version]
- Luo, G.; Hofmann, C.; Bronckers, A.; Sohocki, M.; Bradley, A.; Karsenty, G. BMP-7 is an inducer of nephrogenesis, and is also required for eye development and skeletal patterning. Genes Dev. 1995, 9, 2808–2820. [Google Scholar] [CrossRef] [Green Version]
- Bustos-Valenzuela, J.C.; Halcsik, E.; Bassi, E.J.; Demasi, M.A.; Granjeiro, J.M.; Sogayar, M.C. Expression, purification, bioactivity, and partial characterization of a recombinant human bone morphogenetic protein-7 produced in human 293T cells. Mol. Biotechnol. 2010, 46, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Swencki-Underwood, B.; Mills, J.K.; Vennarini, J.; Boakye, K.; Luo, J.; Pomerantz, S.; Cunningham, M.R.; Farrell, F.X.; Naso, M.F.; Amegadzie, B. Expression and characterization of a human BMP-7 variant with improved biochemical properties. Protein Expr. Purif. 2008, 57, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, N.W. Cystine knots. Curr. Opin. Struct. Biol. 1995, 5, 391–395. [Google Scholar] [CrossRef]
- Sieber, C.; Kopf, J.; Hiepen, C.; Knaus, P. Recent advances in BMP receptor signaling. Cytokine Growth Factor Rev. 2009, 20, 343–355. [Google Scholar] [CrossRef]
- Little, S.C.; Mullins, M.C. Bone morphogenetic protein heterodimers assemble heteromeric type I receptor complexes to pattern the dorsoventral axis. Nat. Cell. Biol. 2009, 11, 637–643. [Google Scholar] [CrossRef] [Green Version]
- Aono, A.; Hazama, M.; Notoya, K.; Taketomi, S.; Yamasaki, H.; Tsukuda, R.; Sasaki, S.; Fujisawa, Y. Potent ectopic bone-inducing activity of bone morphogenetic protein-4/7 heterodimer. Biochem. Biophys. Res. Commun. 1995, 210, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Hazama, M.; Aono, A.; Ueno, N.; Fujisawa, Y. Efficient expression of a heterodimer of bone morphogenetic protein subunits using a baculovirus expression system. Biochem. Biophys. Res. Commun. 1995, 209, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Israel, D.I.; Nove, J.; Kerns, K.M.; Kaufman, R.J.; Rosen, V.; Cox, K.A.; Wozney, J.M. Heterodimeric bone morphogenetic proteins show enhanced activity in vitro and in vivo. Growth Factors 1996, 13, 291–300. [Google Scholar] [CrossRef]
- Dimitriou, R.; Dahabreh, Z.; Katsoulis, E.; Matthews, S.J.; Branfoot, T.; Giannoudis, P.V. Application of recombinant BMP-7 on persistent upper and lower limb non-unions. Injury 2005, 36, 51–59. [Google Scholar] [CrossRef]
- Nishimatsu, S.; Thomsen, G.H. Ventral mesoderm induction and patterning by bone morphogenetic protein heterodimers in Xenopus embryos. Mech. Dev. 1998, 74, 75–88. [Google Scholar] [CrossRef]
- Schmid, B.; Furthauer, M.; Connors, S.A.; Trout, J.; Thisse, B.; Thisse, C.; Mullins, M.C. Equivalent genetic roles for bmp7/snailhouse and bmp2b/swirl in dorsoventral pattern formation. Development 2000, 127, 957–967. [Google Scholar]
- Suzuki, A.; Kaneko, E.; Maeda, J.; Ueno, N. Mesoderm induction by BMP-4 and -7 heterodimers. Biochem. Biophys. Res. Commun. 1997, 232, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-S.; Neugebauer, J.; McKnite, A.; Tilak, A.; Christian, J.L. BMP7 functions predominantly as a heterodimer with BMP2 or BMP4 during mammalian embryogenesis. BioRxiv 2019, 686758. [Google Scholar] [CrossRef] [PubMed]
- Vaccaro, A.R.; Whang, P.G.; Patel, T.; Phillips, F.M.; Anderson, D.G.; Albert, T.J.; Hilibrand, A.S.; Brower, R.S.; Kurd, M.F.; Appannagari, A.; et al. The safety and efficacy of OP-1 (rhBMP-7) as a replacement for iliac crest autograft for posterolateral lumbar arthrodesis: Minimum 4-year follow-up of a pilot study. Spine J. 2008, 8, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Boon, M.R.; van der Horst, G.; van der Pluijm, G.; Tamsma, J.T.; Smit, J.W.; Rensen, P.C. Bone morphogenetic protein 7: A broad-spectrum growth factor with multiple target therapeutic potency. Cytokine Growth Factor Rev. 2011, 22, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Carreira, A.C.; Lojudice, F.H.; Halcsik, E.; Navarro, R.D.; Sogayar, M.C.; Granjeiro, J.M. Bone morphogenetic proteins: Facts, challenges, and future perspectives. J. Dent. Res. 2014, 93, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Rocher, C.; Singla, R.; Singal, P.K.; Parthasarathy, S.; Singla, D.K. Bone morphogenetic protein 7 polarizes THP-1 cells into M2 macrophages. Can. J. Physiol. Pharmacol. 2012, 90, 947–951. [Google Scholar] [CrossRef]
- Cecchi, S.; Bennet, S.J.; Arora, M. Bone morphogenetic protein-7: Review of signalling and efficacy in fracture healing. J. Orthop. Translat. 2015, 4, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Singla, D.K.; Singla, R.; Wang, J. BMP-7 Treatment Increases M2 Macrophage Differentiation and Reduces Inflammation and Plaque Formation in Apo E-/- Mice. PLoS ONE 2016, 11, e0147897. [Google Scholar] [CrossRef]
- Rocher, C.; Singla, D.K. SMAD-PI3K-Akt-mTOR pathway mediates BMP-7 polarization of monocytes into M2 macrophages. PLoS ONE 2013, 8, e84009. [Google Scholar] [CrossRef] [Green Version]
- Shoulders, H.; Garner, K.H.; Singla, D.K. Macrophage depletion by clodronate attenuates bone morphogenetic protein-7 induced M2 macrophage differentiation and improved systolic blood velocity in atherosclerosis. Transl. Res. 2019, 203, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, T.; Singh, R.R.; Gupta, S.; Surolia, A. Bone morphogenetic protein-7 (BMP-7) augments insulin sensitivity in mice with type II diabetes mellitus by potentiating PI3K/AKT pathway. Biofactors 2017, 43, 195–209. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, B.L.; Imamura, T.; Okadome, T.; Cox, G.N.; Yamashita, H.; ten Dijke, P.; Heldin, C.H.; Miyazono, K. Cloning and characterization of a human type II receptor for bone morphogenetic proteins. Proc. Natl. Acad. Sci. USA 1995, 92, 7632–7636. [Google Scholar] [CrossRef] [Green Version]
- Miyazono, K.; Kamiya, Y.; Morikawa, M. Bone morphogenetic protein receptors and signal transduction. J. Biochem. 2010, 147, 35–51. [Google Scholar] [CrossRef] [Green Version]
- Ulsamer, A.; Ortuno, M.J.; Ruiz, S.; Susperregui, A.R.; Osses, N.; Rosa, J.L.; Ventura, F. BMP-2 induces Osterix expression through up-regulation of Dlx5 and its phosphorylation by p38. J. Biol. Chem. 2008, 283, 3816–3826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franceschi, R.T.; Xiao, G. Regulation of the osteoblast-specific transcription factor, Runx2: Responsiveness to multiple signal transduction pathways. J. Cell. Biochem. 2003, 88, 446–454. [Google Scholar] [CrossRef]
- Lee, K.S.; Hong, S.H.; Bae, S.C. Both the Smad and p38 MAPK pathways play a crucial role in Runx2 expression following induction by transforming growth factor-beta and bone morphogenetic protein. Oncogene 2002, 21, 7156–7163. [Google Scholar] [CrossRef] [Green Version]
- Morrell, N.W.; Bloch, D.B.; ten Dijke, P.; Goumans, M.J.; Hata, A.; Smith, J.; Yu, P.B.; Bloch, K.D. Targeting BMP signalling in cardiovascular disease and anaemia. Nat. Rev. Cardiol. 2016, 13, 106–120. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.H.; Derynck, R. Specificity and versatility in tgf-beta signaling through Smads. Annu. Rev. Cell. Dev. Biol. 2005, 21, 659–693. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Chen, G.; Li, Y.P. TGF-beta and BMP signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease. Bone Res. 2016, 4, 16009. [Google Scholar] [CrossRef]
- Macias-Silva, M.; Hoodless, P.A.; Tang, S.J.; Buchwald, M.; Wrana, J.L. Specific activation of Smad1 signaling pathways by the BMP7 type I receptor, ALK2. J. Biol. Chem. 1998, 273, 25628–25636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, R.; Khanna, A. Role of smad- and wnt-dependent pathways in embryonic cardiac development. Stem Cells Dev. 2006, 15, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Ebisawa, T.; Tada, K.; Kitajima, I.; Tojo, K.; Sampath, T.K.; Kawabata, M.; Miyazono, K.; Imamura, T. Characterization of bone morphogenetic protein-6 signaling pathways in osteoblast differentiation. J. Cell Sci. 1999, 112, 3519–3527. [Google Scholar] [PubMed]
- Lavery, K.; Hawley, S.; Swain, P.; Rooney, R.; Falb, D.; Alaoui-Ismaili, M.H. New insights into BMP-7 mediated osteoblastic differentiation of primary human mesenchymal stem cells. Bone 2009, 45, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Yeh, L.C.; Tsai, A.D.; Lee, J.C. Osteogenic protein-1 (OP-1, BMP-7) induces osteoblastic cell differentiation of the pluripotent mesenchymal cell line C2C12. J. Cell Biochem. 2002, 87, 292–304. [Google Scholar] [CrossRef]
- Zhang, J.; Li, L. BMP signaling and stem cell regulation. Dev. Biol. 2005, 284, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Herpin, A.; Cunningham, C. Cross-talk between the bone morphogenetic protein pathway and other major signaling pathways results in tightly regulated cell-specific outcomes. FEBS J. 2007, 274, 2977–2985. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Nagai, S.; Ninomiya-Tsuji, J.; Nishita, M.; Tamai, K.; Irie, K.; Ueno, N.; Nishida, E.; Shibuya, H.; Matsumoto, K. XIAP, a cellular member of the inhibitor of apoptosis protein family, links the receptors to TAB1-TAK1 in the BMP signaling pathway. EMBO J. 1999, 18, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.C.; Yang, S.T.; Lin, C.Y.; Hsu, C.J.; Tsai, C.H.; Su, J.L.; Tang, C.H. BMP-7 enhances cell migration and αvβ3 integrin expression via a c-Src-dependent pathway in human chondrosarcoma cells. PLoS ONE 2014, 9, e112636. [Google Scholar] [CrossRef]
- Yeh, C.H.; Chang, C.K.; Cheng, M.F.; Lin, H.J.; Cheng, J.T. The antioxidative effect of bone morphogenetic protein-7 against high glucose-induced oxidative stress in mesangial cells. Biochem. Biophys. Res. Commun. 2009, 382, 292–297. [Google Scholar] [CrossRef]
- Yeh, L.C.; Ma, X.; Matheny, R.W.; Adamo, M.L.; Lee, J.C. Protein kinase D mediates the synergistic effects of BMP-7 and IGF-I on osteoblastic cell differentiation. Growth Factors 2010, 28, 318–328. [Google Scholar] [CrossRef]
- Klatte-Schulz, F.; Giese, G.; Differ, C.; Minkwitz, S.; Ruschke, K.; Puts, R.; Knaus, P.; Wildemann, B. An investigation of BMP-7 mediated alterations to BMP signalling components in human tenocyte-like cells. Sci. Rep. 2016, 6, 29703. [Google Scholar] [CrossRef]
- Hu, M.C.; Wasserman, D.; Hartwig, S.; Rosenblum, N.D. p38MAPK acts in the BMP7-dependent stimulatory pathway during epithelial cell morphogenesis and is regulated by Smad1. J. Biol. Chem. 2004, 279, 12051–12059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blank, U.; Brown, A.; Adams, D.C.; Karolak, M.J.; Oxburgh, L. BMP7 promotes proliferation of nephron progenitor cells via a JNK-dependent mechanism. Development 2009, 136, 3557–3566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sovershaev, T.A.; Egorina, E.M.; Unruh, D.; Bogdanov, V.Y.; Hansen, J.B.; Sovershaev, M.A. BMP-7 induces TF expression in human monocytes by increasing F3 transcriptional activity. Thromb. Res. 2015, 135, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Kayamori, T.; Murayama, C.; Miyamoto, A. Bone morphogenetic protein (BMP)-4 and BMP-7 suppress granulosa cell apoptosis via different pathways: BMP-4 via PI3K/PDK-1/Akt and BMP-7 via PI3K/PDK-1/PKC. Biochem. Biophys. Res. Commun. 2012, 417, 869–873. [Google Scholar] [CrossRef]
- Weichhart, T.; Saemann, M.D. The PI3K/Akt/mTOR pathway in innate immune cells: Emerging therapeutic applications. Ann. Rheum. Dis. 2008, 67, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Rauh, M.J.; Ho, V.; Pereira, C.; Sham, A.; Sly, L.M.; Lam, V.; Huxham, L.; Minchinton, A.I.; Mui, A.; Krystal, G. SHIP represses the generation of alternatively activated macrophages. Immunity 2005, 23, 361–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gazzerro, E.; Gangji, V.; Canalis, E. Bone morphogenetic proteins induce the expression of noggin, which limits their activity in cultured rat osteoblasts. J. Clin. Investig. 1998, 102, 2106–2114. [Google Scholar] [CrossRef]
- Dionne, M.S.; Skarnes, W.C.; Harland, R.M. Mutation and analysis of Dan, the founding member of the Dan family of transforming growth factor beta antagonists. Mol. Cell. Biol. 2001, 21, 636–643. [Google Scholar] [CrossRef] [Green Version]
- Ideno, H.; Takanabe, R.; Shimada, A.; Imaizumi, K.; Araki, R.; Abe, M.; Nifuji, A. Protein related to DAN and cerberus (PRDC) inhibits osteoblastic differentiation and its suppression promotes osteogenesis in vitro. Exp. Cell. Res. 2009, 315, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Topol, L.Z.; Bardot, B.; Zhang, Q.; Resau, J.; Huillard, E.; Marx, M.; Calothy, G.; Blair, D.G. Biosynthesis, post-translation modification, and functional characterization of Drm/Gremlin. J. Biol. Chem. 2000, 275, 8785–8793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudo, S.; Avsian-Kretchmer, O.; Wang, L.S.; Hsueh, A.J. Protein related to DAN and cerberus is a bone morphogenetic protein antagonist that participates in ovarian paracrine regulation. J. Biol. Chem. 2004, 279, 23134–23141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balemans, W.; Van Hul, W. Extracellular regulation of BMP signaling in vertebrates: A cocktail of modulators. Dev. Biol. 2002, 250, 231–250. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Akhtar, N.; Jamil, H.M.; Banik, R.S.; Asaduzzaman, S.M. TGF-beta/BMP signaling and other molecular events: Regulation of osteoblastogenesis and bone formation. Bone Res. 2015, 3, 15005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, S.; ten Dijke, P. Negative regulation of TGF-beta receptor/Smad signal transduction. Curr. Opin. Cell Biol. 2007, 19, 176–184. [Google Scholar] [CrossRef]
- Knockaert, M.; Sapkota, G.; Alarcon, C.; Massague, J.; Brivanlou, A.H. Unique players in the BMP pathway: Small C-terminal domain phosphatases dephosphorylate Smad1 to attenuate BMP signaling. Proc. Natl. Acad. Sci. USA 2006, 103, 11940–11945. [Google Scholar] [CrossRef] [Green Version]
- Heldin, C.H.; Moustakas, A. Role of Smads in TGFbeta signaling. Cell Tissue Res. 2012, 347, 21–36. [Google Scholar] [CrossRef]
- Larman, B.W.; Karolak, M.J.; Adams, D.C.; Oxburgh, L. Chordin-like 1 and twisted gastrulation 1 regulate BMP signaling following kidney injury. J. Am. Soc. Nephrol. 2009, 20, 1020–1031. [Google Scholar] [CrossRef] [Green Version]
- Matzuk, M.M.; Lu, N.; Vogel, H.; Sellheyer, K.; Roop, D.R.; Bradley, A. Multiple defects and perinatal death in mice deficient in follistatin. Nature 1995, 374, 360–363. [Google Scholar] [CrossRef]
- Amthor, H.; Christ, B.; Rashid-Doubell, F.; Kemp, C.F.; Lang, E.; Patel, K. Follistatin regulates bone morphogenetic protein-7 (BMP-7) activity to stimulate embryonic muscle growth. Dev. Biol. 2002, 243, 115–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Lu, Y.; Han, R.; Yue, Q.; Song, X.; Wang, F.; Wu, R.; Hou, F.; Yang, L.; Xu, L.; et al. Gremlin2 Regulates the Differentiation and Function of Cardiac Progenitor Cells via the Notch Signaling Pathway. Cell. Physiol. Biochem. 2018, 47, 579–589. [Google Scholar] [CrossRef]
- Sasai, Y.; Lu, B.; Steinbeisser, H.; De Robertis, E.M. Regulation of neural induction by the Chd and Bmp-4 antagonistic patterning signals in Xenopus. Nature 1995, 377, 757. [Google Scholar] [CrossRef] [PubMed]
- Bachiller, D.; Klingensmith, J.; Kemp, C.; Belo, J.A.; Anderson, R.M.; May, S.R.; McMahon, J.A.; McMahon, A.P.; Harland, R.M.; Rossant, J.; et al. The organizer factors Chordin and Noggin are required for mouse forebrain development. Nature 2000, 403, 658–661. [Google Scholar] [CrossRef] [PubMed]
- Wijgerde, M.; Karp, S.; McMahon, J.; McMahon, A.P. Noggin antagonism of BMP4 signaling controls development of the axial skeleton in the mouse. Dev. Biol. 2005, 286, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Coffinier, C.; Tran, U.; Larrain, J.; De Robertis, E.M. Neuralin-1 is a novel Chordin-related molecule expressed in the mouse neural plate. Mech. Dev. 2001, 100, 119–122. [Google Scholar] [CrossRef]
- Nakayama, N.; Han, C.E.; Scully, S.; Nishinakamura, R.; He, C.; Zeni, L.; Yamane, H.; Chang, D.; Yu, D.; Yokota, T.; et al. A novel chordin-like protein inhibitor for bone morphogenetic proteins expressed preferentially in mesenchymal cell lineages. Dev. Biol. 2001, 232, 372–387. [Google Scholar] [CrossRef] [Green Version]
- Sakuta, H.; Suzuki, R.; Takahashi, H.; Kato, A.; Shintani, T.; Iemura, S.; Yamamoto, T.S.; Ueno, N.; Noda, M. Ventroptin: A BMP-4 antagonist expressed in a double-gradient pattern in the retina. Science 2001, 293, 111–115. [Google Scholar] [CrossRef]
- Ikeya, M.; Kawada, M.; Kiyonari, H.; Sasai, N.; Nakao, K.; Furuta, Y.; Sasai, Y. Essential pro-Bmp roles of crossveinless 2 in mouse organogenesis. Development 2006, 133, 4463–4473. [Google Scholar] [CrossRef] [Green Version]
- Rentzsch, F.; Zhang, J.; Kramer, C.; Sebald, W.; Hammerschmidt, M. Crossveinless 2 is an essential positive feedback regulator of Bmp signaling during zebrafish gastrulation. Development 2006, 133, 801–811. [Google Scholar] [CrossRef] [Green Version]
- Zakin, L.; Metzinger, C.A.; Chang, E.Y.; Coffinier, C.; De Robertis, E.M. Development of the vertebral morphogenetic field in the mouse: Interactions between Crossveinless-2 and Twisted Gastrulation. Dev. Biol. 2008, 323, 6–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, Y.; Jumabay, M.; Ly, A.; Radparvar, M.; Wang, A.H.; Abdmaulen, R.; Bostrom, K.I. Crossveinless 2 regulates bone morphogenetic protein 9 in human and mouse vascular endothelium. Blood 2012, 119, 5037–5047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichert, S.; Randall, R.A.; Hill, C.S. A BMP regulatory network controls ectodermal cell fate decisions at the neural plate border. Development 2013, 140, 4435–4444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyer, L.; Wu, Y.; Moser, M.; Patterson, C. BMPER-induced BMP signaling promotes coronary artery remodeling. Dev. Biol. 2014, 386, 385–394. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Patel, S.R.; Cheng, X.; Cho, E.A.; Levitan, I.; Ullenbruch, M.; Phan, S.H.; Park, J.M.; Dressler, G.R. Kielin/chordin-like protein, a novel enhancer of BMP signaling, attenuates renal fibrotic disease. Nat. Med. 2005, 11, 387–393. [Google Scholar] [CrossRef]
- Soofi, A.; Zhang, P.; Dressler, G.R. Kielin/chordin-like protein attenuates both acute and chronic renal injury. J. Am. Soc. Nephrol. 2013, 24, 897–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivkovic, S.; Yoon, B.S.; Popoff, S.N.; Safadi, F.F.; Libuda, D.E.; Stephenson, R.C.; Daluiski, A.; Lyons, K.M. Connective tissue growth factor coordinates chondrogenesis and angiogenesis during skeletal development. Development 2003, 130, 2779–2791. [Google Scholar] [CrossRef] [Green Version]
- Maeda, A.; Nishida, T.; Aoyama, E.; Kubota, S.; Lyons, K.M.; Kuboki, T.; Takigawa, M. CCN family 2/connective tissue growth factor modulates BMP signalling as a signal conductor, which action regulates the proliferation and differentiation of chondrocytes. J. Biochem. 2009, 145, 207–216. [Google Scholar] [CrossRef] [Green Version]
- Mundy, C.; Gannon, M.; Popoff, S.N. Connective tissue growth factor (CTGF/CCN2) negatively regulates BMP-2 induced osteoblast differentiation and signaling. J. Cell. Physiol. 2014, 229, 672–681. [Google Scholar] [CrossRef] [Green Version]
- Glass, C.K.; Witztum, J.L. Atherosclerosis. the road ahead. Cell 2001, 104, 503–516. [Google Scholar] [CrossRef] [Green Version]
- Libby, P. Inflammation in atherosclerosis. Nature 2002, 420, 868–874. [Google Scholar] [CrossRef] [PubMed]
- Hansson, G.K. Inflammation, atherosclerosis, and coronary artery disease. N. Engl. J. Med. 2005, 352, 1685–1695. [Google Scholar] [CrossRef] [Green Version]
- Bentzon, J.F.; Otsuka, F.; Virmani, R.; Falk, E. Mechanisms of plaque formation and rupture. Circ. Res. 2014, 114, 1852–1866. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathy, S. Modified Lipoproteins in the Pathogenesis of Atherosclerosis; RG Landes Co: Austin, TX, USA, 1994. [Google Scholar]
- Steinberg, D. Thematic review series: The pathogenesis of atherosclerosis: An interpretive history of the cholesterol controversy, part III: Mechanistically defining the role of hyperlipidemia. J. Lipid. Res. 2005, 46, 2037–2051. [Google Scholar] [CrossRef] [Green Version]
- Galkina, E.; Ley, K. Leukocyte influx in atherosclerosis. Curr. Drug Targets 2007, 8, 1239–1248. [Google Scholar] [CrossRef]
- Weber, C.; Zernecke, A.; Libby, P. The multifaceted contributions of leukocyte subsets to atherosclerosis: Lessons from mouse models. Nat. Rev. Immunol. 2008, 8, 802–815. [Google Scholar] [CrossRef]
- Libby, P.; Lichtman, A.H.; Hansson, G.K. Immune effector mechanisms implicated in atherosclerosis: From mice to humans. Immunity 2013, 38, 1092–1104. [Google Scholar] [CrossRef] [Green Version]
- Steven, S.; Frenis, K.; Oelze, M.; Kalinovic, S.; Kuntic, M.; Bayo Jimenez, M.T.; Vujacic-Mirski, K.; Helmstädter, J.; Kröller-Schön, S.; Münzel, T.; et al. Vascular Inflammation and Oxidative Stress: Major Triggers for Cardiovascular Disease. Oxidative Med. Cell. Longev. 2019, 2019, 26. [Google Scholar] [CrossRef] [Green Version]
- Steinberg, D.; Witztum, J.L. Lipoproteins and atherogenesis. Current concepts. Jama 1990, 264, 3047–3052. [Google Scholar] [CrossRef]
- Ross, R. The pathogenesis of atherosclerosis: A perspective for the 1990s. Nature 1993, 362, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Swirski, F.K.; Pittet, M.J.; Kircher, M.F.; Aikawa, E.; Jaffer, F.A.; Libby, P.; Weissleder, R. Monocyte accumulation in mouse atherogenesis is progressive and proportional to extent of disease. Proc. Natl. Acad. Sci. USA 2006, 103, 10340–10345. [Google Scholar] [CrossRef] [Green Version]
- Hahn, C.; Schwartz, M.A. Mechanotransduction in vascular physiology and atherogenesis. Na. Rev. Mol. Cell Biol. 2009, 10, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Bi, Y.; Chen, J.; Hu, F.; Liu, J.; Li, M.; Zhao, L. M2 Macrophages as a Potential Target for Antiatherosclerosis Treatment. Neural Plast. 2019, 2019, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Tits, L.J.; Stienstra, R.; van Lent, P.L.; Netea, M.G.; Joosten, L.A.; Stalenhoef, A.F. Oxidized LDL enhances pro-inflammatory responses of alternatively activated M2 macrophages: A crucial role for Kruppel-like factor 2. Atherosclerosis 2011, 214, 345–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinon, F. Signaling by ROS drives inflammasome activation. Eur. J. Immunol. 2010, 40, 616–619. [Google Scholar] [CrossRef] [PubMed]
- Chinetti-Gbaguidi, G.; Baron, M.; Bouhlel, M.A.; Vanhoutte, J.; Copin, C.; Sebti, Y.; Derudas, B.; Mayi, T.; Bories, G.; Tailleux, A.; et al. Human atherosclerotic plaque alternative macrophages display low cholesterol handling but high phagocytosis because of distinct activities of the PPARgamma and LXRalpha pathways. Circ. Res. 2011, 108, 985–995. [Google Scholar] [CrossRef] [PubMed]
- Abdelbaky, A.; Corsini, E.; Figueroa, A.L.; Fontanez, S.; Subramanian, S.; Ferencik, M.; Brady, T.J.; Hoffmann, U.; Tawakol, A. Focal arterial inflammation precedes subsequent calcification in the same location: A longitudinal FDG-PET/CT study. Circ. Cardiovasc. Imaging 2013, 6, 747–754. [Google Scholar] [CrossRef] [Green Version]
- Bobryshev, Y.V. Transdifferentiation of smooth muscle cells into chondrocytes in atherosclerotic arteries in situ: Implications for diffuse intimal calcification. J. Pathol. 2005, 205, 641–650. [Google Scholar] [CrossRef]
- Lanzer, P.; Boehm, M.; Sorribas, V.; Thiriet, M.; Janzen, J.; Zeller, T.; St Hilaire, C.; Shanahan, C. Medial vascular calcification revisited: Review and perspectives. Eur. Heart. J. 2014, 35, 1515–1525. [Google Scholar] [CrossRef]
- Hruska, K.A.; Mathew, S.; Saab, G. Bone morphogenetic proteins in vascular calcification. Circ. Res. 2005, 97, 105–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riad, A.; Narasimhulu, C.A.; Deme, P.; Parthasarathy, S. A Novel Mechanism for Atherosclerotic Calcification: Potential Resolution of the Oxidation Paradox. Antioxid. Redox Signal. 2018, 29, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Nakaoka, T.; Gonda, K.; Ogita, T.; Otawara-Hamamoto, Y.; Okabe, F.; Kira, Y.; Harii, K.; Miyazono, K.; Takuwa, Y.; Fujita, T. Inhibition of rat vascular smooth muscle proliferation in vitro and in vivo by bone morphogenetic protein-2. J. Clin. Investig. 1997, 100, 2824–2832. [Google Scholar] [CrossRef] [PubMed]
- Wong, G.A.; Tang, V.; El-Sabeawy, F.; Weiss, R.H. BMP-2 inhibits proliferation of human aortic smooth muscle cells via p21Cip1/Waf1. Am. J. Physiol. Endocrinol. Metab. 2003, 284, E972–E979. [Google Scholar] [CrossRef]
- Matsushita, H.; Morishita, R.; Kida, I.; Aoki, M.; Hayashi, S.; Tomita, N.; Yamamoto, K.; Moriguchi, A.; Noda, A.; Kaneda, Y.; et al. Inhibition of growth of human vascular smooth muscle cells by overexpression of p21 gene through induction of apoptosis. Hypertension 1998, 31, 493–498. [Google Scholar] [CrossRef] [Green Version]
- Dorai, H.; Vukicevic, S.; Sampath, T.K. Bone morphogenetic protein-7 (osteogenic protein-1) inhibits smooth muscle cell proliferation and stimulates the expression of markers that are characteristic of SMC phenotype in vitro. J. Cell. Physiol. 2000, 184, 37–45. [Google Scholar] [CrossRef]
- Dorai, H.; Sampath, T.K. Bone morphogenetic protein-7 modulates genes that maintain the vascular smooth muscle cell phenotype in culture. J. Bone Jt. Surg. Am. Vol. 2001, 83, 70–78. [Google Scholar] [CrossRef]
- Li, X.; Yang, H.Y.; Giachelli, C.M. BMP-2 promotes phosphate uptake, phenotypic modulation, and calcification of human vascular smooth muscle cells. Atherosclerosis 2008, 199, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Tobin, J.F.; Celeste, A.J. Bone morphogenetic proteins and growth differentiation factors as drug targets in cardiovascular and metabolic disease. Drug. Discov. Today 2006, 11, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Vattikuti, R.; Towler, D.A. Osteogenic regulation of vascular calcification: An early perspective. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E686–E696. [Google Scholar] [CrossRef]
- Johnson, R.C.; Leopold, J.A.; Loscalzo, J. Vascular calcification: Pathobiological mechanisms and clinical implications. Circ. Res. 2006, 99, 1044–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, M.R.; Lund, R.J.; Hruska, K.A. BMP-7 is an efficacious treatment of vascular calcification in a murine model of atherosclerosis and chronic renal failure. J. Am. Soc. Nephrol. 2003, 14, 1559–1567. [Google Scholar] [CrossRef] [Green Version]
- Morrell, N.W.; Yang, X.; Upton, P.D.; Jourdan, K.B.; Morgan, N.; Sheares, K.K.; Trembath, R.C. Altered growth responses of pulmonary artery smooth muscle cells from patients with primary pulmonary hypertension to transforming growth factor-beta(1) and bone morphogenetic proteins. Circulation 2001, 104, 790–795. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.H.; Jin, J.S.; Yi, D.W.; Son, S.M. Bone morphogenetic protein-7 inhibits vascular calcification induced by high vitamin D in mice. Tohoku J. Exp. Med. 2010, 221, 299–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.; Liu, M.; Sun, R.; Zheng, Y.; Zhang, P. Myocardial Infarction: Symptoms and Treatments. Cell Biochem. Biophys. 2015, 72, 865–867. [Google Scholar] [CrossRef]
- Swynghedauw, B. Molecular mechanisms of myocardial remodeling. Physiol. Rev. 1999, 79, 215–262. [Google Scholar] [CrossRef]
- Holmes, J.W.; Borg, T.K.; Covell, J.W. Structure and mechanics of healing myocardial infarcts. Annu. Rev. Biomed. Eng. 2005, 7, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Jugdutt, B.I. Remodeling of the myocardium and potential targets in the collagen degradation and synthesis pathways. Curr. Drug Targets Cardiovasc. Haematol. Disord. 2003, 3, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Jugdutt, B.I. Ventricular remodeling after infarction and the extracellular collagen matrix: When is enough enough? Circulation 2003, 108, 1395–1403. [Google Scholar] [CrossRef] [Green Version]
- Jugdutt, B.I.; Menon, V.; Kumar, D.; Idikio, H. Vascular remodeling during healing after myocardial infarction in the dog model: Effects of reperfusion, amlodipine and enalapril. J. Am. Coll. Cardiol. 2002, 39, 1538–1545. [Google Scholar] [CrossRef] [Green Version]
- Alibhai, F.J.; Tsimakouridze, E.V.; Chinnappareddy, N.; Wright, D.C.; Billia, F.; O’Sullivan, M.L.; Pyle, W.G.; Sole, M.J.; Martino, T.A. Short-term disruption of diurnal rhythms after murine myocardial infarction adversely affects long-term myocardial structure and function. Circ. Res. 2014, 114, 1713–1722. [Google Scholar] [CrossRef] [Green Version]
- Nahrendorf, M.; Swirski, F.K.; Aikawa, E.; Stangenberg, L.; Wurdinger, T.; Figueiredo, J.L.; Libby, P.; Weissleder, R.; Pittet, M.J. The healing myocardium sequentially mobilizes two monocyte subsets with divergent and complementary functions. J. Exp. Med. 2007, 204, 3037–3047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dewald, O.; Zymek, P.; Winkelmann, K.; Koerting, A.; Ren, G.; Abou-Khamis, T.; Michael, L.H.; Rollins, B.J.; Entman, M.L.; Frangogiannis, N.G. CCL2/Monocyte Chemoattractant Protein-1 regulates inflammatory responses critical to healing myocardial infarcts. Circ. Res. 2005, 96, 881–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zouggari, Y.; Ait-Oufella, H.; Bonnin, P.; Simon, T.; Sage, A.P.; Guerin, C.; Vilar, J.; Caligiuri, G.; Tsiantoulas, D.; Laurans, L.; et al. B lymphocytes trigger monocyte mobilization and impair heart function after acute myocardial infarction. Nat. Med. 2013, 19, 1273–1280. [Google Scholar] [CrossRef]
- Hilgendorf, I.; Gerhardt, L.M.; Tan, T.C.; Winter, C.; Holderried, T.A.; Chousterman, B.G.; Iwamoto, Y.; Liao, R.; Zirlik, A.; Scherer-Crosbie, M.; et al. Ly-6Chigh monocytes depend on Nr4a1 to balance both inflammatory and reparative phases in the infarcted myocardium. Circ. Res. 2014, 114, 1611–1622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frangogiannis, N.G.; Smith, C.W.; Entman, M.L. The inflammatory response in myocardial infarction. Cardiovasc. Res. 2002, 53, 31–47. [Google Scholar] [CrossRef]
- Fadok, V.A.; Bratton, D.L.; Konowal, A.; Freed, P.W.; Westcott, J.Y.; Henson, P.M. Macrophages that have ingested apoptotic cells in vitro inhibit proinflammatory cytokine production through autocrine/paracrine mechanisms involving TGF-beta, PGE2, and PAF. J. Clin. Investig. 1998, 101, 890–898. [Google Scholar] [CrossRef] [Green Version]
- Freire-de-Lima, C.G.; Xiao, Y.Q.; Gardai, S.J.; Bratton, D.L.; Schiemann, W.P.; Henson, P.M. Apoptotic cells, through transforming growth factor-beta, coordinately induce anti-inflammatory and suppress pro-inflammatory eicosanoid and NO synthesis in murine macrophages. J. Biol. Chem. 2006, 281, 38376–38384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korns, D.; Frasch, S.C.; Fernandez-Boyanapalli, R.; Henson, P.M.; Bratton, D.L. Modulation of macrophage efferocytosis in inflammation. Front. Immunol. 2011, 2, 57. [Google Scholar] [CrossRef] [Green Version]
- Narula, J.; Haider, N.; Virmani, R.; DiSalvo, T.G.; Kolodgie, F.D.; Hajjar, R.J.; Schmidt, U.; Semigran, M.J.; Dec, G.W.; Khaw, B.A. Apoptosis in myocytes in end-stage heart failure. N. Engl. J. Med. 1996, 335, 1182–1189. [Google Scholar] [CrossRef]
- Sharov, V.G.; Sabbah, H.N.; Shimoyama, H.; Goussev, A.V.; Lesch, M.; Goldstein, S. Evidence of cardiocyte apoptosis in myocardium of dogs with chronic heart failure. Am. J. Pathol. 1996, 148, 141–149. [Google Scholar]
- Intengan, H.D.; Schiffrin, E.L. Vascular remodeling in hypertension: Roles of apoptosis, inflammation, and fibrosis. Hypertension 2001, 38, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Buja, L.M.; Vela, D. Cardiomyocyte death and renewal in the normal and diseased heart. Cardiovasc. Pathol. 2008, 17, 349–374. [Google Scholar] [CrossRef] [PubMed]
- Talman, V.; Ruskoaho, H. Cardiac fibrosis in myocardial infarction-from repair and remodeling to regeneration. Cell Tissue Res. 2016, 365, 563–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barin, J.G.; Rose, N.R.; Ciháková, D. Macrophage diversity in cardiac inflammation: A review. Immunobiology 2012, 217, 468–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, F.; Li, Y.; Jia, L.; Han, Y.; Cheng, J.; Li, H.; Qi, Y.; Du, J. Macrophage-stimulated cardiac fibroblast production of IL-6 is essential for TGF beta/Smad activation and cardiac fibrosis induced by angiotensin II. PLoS ONE 2012, 7, e35144. [Google Scholar] [CrossRef] [Green Version]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoyagi, T.; Matsui, T. The Cardiomyocyte as a Source of Cytokines in Cardiac Injury. J. Cell Sci. Ther. 2011, 2012, 003. [Google Scholar] [CrossRef] [Green Version]
- Han, G.; Li, F.; Singh, T.P.; Wolf, P.; Wang, X.-J. The pro-inflammatory role of TGFβ1: A paradox? Int. J. Biol. Sci. 2012, 8, 228–235. [Google Scholar] [CrossRef] [Green Version]
- Sanjabi, S.; Zenewicz, L.A.; Kamanaka, M.; Flavell, R.A. Anti-inflammatory and pro-inflammatory roles of TGF-beta, IL-10, and IL-22 in immunity and autoimmunity. Curr. Opin. Pharmacol. 2009, 9, 447–453. [Google Scholar] [CrossRef] [Green Version]
- Ramesh, S.; Wildey, G.M.; Howe, P.H. Transforming growth factor beta (TGFbeta)-induced apoptosis: The rise & fall of Bim. Cell Cycle 2009, 8, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Cucoranu, I.; Clempus, R.; Dikalova, A.; Phelan, P.J.; Ariyan, S.; Dikalov, S.; Sorescu, D. NAD(P)H oxidase 4 mediates transforming growth factor-beta1-induced differentiation of cardiac fibroblasts into myofibroblasts. Circ. Res. 2005, 97, 900–907. [Google Scholar] [CrossRef] [Green Version]
- Hao, J.; Ju, H.; Zhao, S.; Junaid, A.; Scammell-La Fleur, T.; Dixon, I.M. Elevation of expression of Smads 2, 3, and 4, decorin and TGF-beta in the chronic phase of myocardial infarct scar healing. J. Mol. Cell. Cardiol. 1999, 31, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Li, T.S.; Hayashi, M.; Ito, H.; Furutani, A.; Murata, T.; Matsuzaki, M.; Hamano, K. Regeneration of infarcted myocardium by intramyocardial implantation of ex vivo transforming growth factor-beta-preprogrammed bone marrow stem cells. Circulation 2005, 111, 2438–2445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.Z.; Fan, T.T.; Gao, F.; Fu, J.; Liu, Q. Exogenous cytochrome c inhibits the expression of transforming growth factor-beta1 in a mouse model of sepsis-induced myocardial dysfunction via the SMAD1/5/8 signaling pathway. Mol. Med. Rep. 2015, 12, 2189–2196. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Wang, B.; Jones, S.C.; Jassal, D.S.; Dixon, I.M. Interaction between angiotensin II and Smad proteins in fibroblasts in failing heart and in vitro. Am. J. Physiol. Heart Circ. Physiol. 2000, 279, 3020–3030. [Google Scholar] [CrossRef]
- Schneiders, D.; Heger, J.; Best, P.; Michael Piper, H.; Taimor, G. SMAD proteins are involved in apoptosis induction in ventricular cardiomyocytes. Cardiovasc. Res. 2005, 67, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Heger, J.; Warga, B.; Meyering, B.; Abdallah, Y.; Schluter, K.D.; Piper, H.M.; Euler, G. TGFbeta receptor activation enhances cardiac apoptosis via SMAD activation and concomitant NO release. J. Cell. Physiol. 2011, 226, 2683–2690. [Google Scholar] [CrossRef]
- Gabriel, V.A. Transforming growth factor-beta and angiotensin in fibrosis and burn injuries. J. Burn Care Res. 2009, 30, 471–481. [Google Scholar] [CrossRef]
- Rosenkranz, S.; Flesch, M.; Amann, K.; Haeuseler, C.; Kilter, H.; Seeland, U.; Schluter, K.D.; Bohm, M. Alterations of beta-adrenergic signaling and cardiac hypertrophy in transgenic mice overexpressing TGF-beta(1). Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H1253–H1262. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Vaughan, D.E. PAI-1 in tissue fibrosis. J. Cell. Physiol. 2012, 227, 493–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derynck, R.; Zhang, Y.E. Smad-dependent and Smad-independent pathways in TGF-beta family signalling. Nature 2003, 425, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Engel, M.E.; McDonnell, M.A.; Law, B.K.; Moses, H.L. Interdependent SMAD and JNK signaling in transforming growth factor-beta-mediated transcription. J. Biol. Chem. 1999, 274, 37413–37420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Hebert, M.C.; Zhang, Y.E. TGF-beta receptor-activated p38 MAP kinase mediates Smad-independent TGF-beta responses. EMBO J. 2002, 21, 3749–3759. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, H.; Abdollah, S.; Qiu, Y.; Cai, J.; Xu, Y.Y.; Grinnell, B.W.; Richardson, M.A.; Topper, J.N.; Gimbrone, M.A., Jr.; Wrana, J.L.; et al. The MAD-related protein Smad7 associates with the TGFbeta receptor and functions as an antagonist of TGFbeta signaling. Cell 1997, 89, 1165–1173. [Google Scholar] [CrossRef] [Green Version]
- Walton, K.L.; Johnson, K.E.; Harrison, C.A. Targeting TGF-beta Mediated SMAD Signaling for the Prevention of Fibrosis. Front. Pharmacol. 2017, 8, 461. [Google Scholar] [CrossRef] [Green Version]
- Zeisberg, M.; Bottiglio, C.; Kumar, N.; Maeshima, Y.; Strutz, F.; Muller, G.A.; Kalluri, R. Bone morphogenic protein-7 inhibits progression of chronic renal fibrosis associated with two genetic mouse models. Am. J. Physiol. Renal. Physiol. 2003, 285, 1060–1067. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, K.; Iimuro, Y.; Otogawa, K.; Saika, S.; Inagaki, Y.; Nakajima, Y.; Kawada, N.; Fujimoto, J.; Friedman, S.L.; Ikeda, K. Adenovirus-mediated expression of BMP-7 suppresses the development of liver fibrosis in rats. Gut 2007, 56, 706–714. [Google Scholar] [CrossRef]
- Liang, D.; Wang, Y.; Zhu, Z.; Yang, G.; An, G.; Li, X.; Niu, P.; Chen, L.; Tian, L. BMP-7 attenuated silica-induced pulmonary fibrosis through modulation of the balance between TGF-beta/Smad and BMP-7/Smad signaling pathway. Chem. Biol. Interact. 2016, 243, 72–81. [Google Scholar] [CrossRef]
- Vukicevic, S.; Basic, V.; Rogic, D.; Basic, N.; Shih, M.S.; Shepard, A.; Jin, D.; Dattatreyamurty, B.; Jones, W.; Dorai, H.; et al. Osteogenic protein-1 (bone morphogenetic protein-7) reduces severity of injury after ischemic acute renal failure in rat. J. Clin. Investig. 1998, 102, 202–214. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Cheng, X.; Lu, J.; Li, X. Exogenous BMP-7 Facilitates the Recovery of Cardiac Function after Acute Myocardial Infarction through Counteracting TGF-beta1 Signaling Pathway. Tohoku J. Exp. Med. 2018, 244, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Zheng, S.; Yang, J.; Wu, Y.; Ren, Y.; Kong, X.; Li, W.; Xuan, J. Suppression of TGF-beta1/Smad signaling pathway by sesamin contributes to the attenuation of myocardial fibrosis in spontaneously hypertensive rats. PLoS ONE 2015, 10, e0121312. [Google Scholar] [CrossRef] [Green Version]
- Merino, D.; Villar, A.V.; Garcia, R.; Tramullas, M.; Ruiz, L.; Ribas, C.; Cabezudo, S.; Nistal, J.F.; Hurle, M.A. BMP-7 attenuates left ventricular remodelling under pressure overload and facilitates reverse remodelling and functional recovery. Cardiovasc. Res. 2016, 110, 331–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmet, P.Z. Diabetes and its drivers: The largest epidemic in human history? Clin. Diabetes Endocrinol. 2017, 3, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maahs, D.M.; West, N.A.; Lawrence, J.M.; Mayer-Davis, E.J. Epidemiology of type 1 diabetes. Endocrinol. Metab. Clin. North Am. 2010, 39, 481–497. [Google Scholar] [CrossRef] [Green Version]
- Nathan, D.M.; Davidson, M.B.; DeFronzo, R.A.; Heine, R.J.; Henry, R.R.; Pratley, R.; Zinman, B. Impaired Fasting Glucose and Impaired Glucose Tolerance. Implic. Care 2007, 30, 753–759. [Google Scholar] [CrossRef] [Green Version]
- Aragno, M.; Mastrocola, R.; Catalano, M.G.; Brignardello, E.; Danni, O.; Boccuzzi, G. Oxidative stress impairs skeletal muscle repair in diabetic rats. Diabetes 2004, 53, 1082–1088. [Google Scholar] [CrossRef] [Green Version]
- Bloomgarden, Z.T. Diabetic nephropathy. Diabetes Care 2005, 28, 745–751. [Google Scholar] [CrossRef] [Green Version]
- Bloomgarden, Z.T. Diabetic retinopathy and diabetic neuropathy. Diabetes Care 2007, 30, 760–765. [Google Scholar] [CrossRef] [Green Version]
- D’Souza, A.; Howarth, F.C.; Yanni, J.; Dobryznski, H.; Boyett, M.R.; Adeghate, E.; Bidasee, K.R.; Singh, J. Left ventricle structural remodelling in the prediabetic Goto-Kakizaki rat. Exp. Physiol. 2011, 96, 875–888. [Google Scholar] [CrossRef]
- Oberley, L.W. Free radicals and diabetes. Free Radic. Biol. Med. 1988, 5, 113–124. [Google Scholar] [CrossRef]
- Wolff, S.P.; Jiang, Z.Y.; Hunt, J.V. Protein glycation and oxidative stress in diabetes mellitus and ageing. Free Radic. Biol. Med. 1991, 10, 339–352. [Google Scholar] [CrossRef]
- Baynes, J.W.; Thorpe, S.R. Role of oxidative stress in diabetic complications: A new perspective on an old paradigm. Diabetes 1999, 48, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Baynes, J.W. Role of oxidative stress in development of complications in diabetes. Diabetes 1991, 40, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Pickup, J.C.; Crook, M.A. Is type II diabetes mellitus a disease of the innate immune system? Diabetologia 1998, 41, 1241–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, S.; Martin, S.; Koenig, W.; Hanifi-Moghaddam, P.; Rathmann, W.; Haastert, B.; Giani, G.; Illig, T.; Thorand, B.; Kolb, H. Impaired glucose tolerance is associated with increased serum concentrations of interleukin 6 and co-regulated acute-phase proteins but not TNF-alpha or its receptors. Diabetologia 2002, 45, 805–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickup, J.C.; Chusney, G.D.; Thomas, S.M.; Burt, D. Plasma interleukin-6, tumour necrosis factor alpha and blood cytokine production in type 2 diabetes. Life Sci. 2000, 67, 291–300. [Google Scholar] [CrossRef]
- Rehman, K.; Akash, M.S.H.; Liaqat, A.; Kamal, S.; Qadir, M.I.; Rasul, A. Role of Interleukin-6 in Development of Insulin Resistance and Type 2 Diabetes Mellitus. Crit. Rev. Eukaryot. Gene Expr. 2017, 27, 229–236. [Google Scholar] [CrossRef]
- Dhingra, S.; Bagchi, A.K.; Ludke, A.L.; Sharma, A.K.; Singal, P.K. Akt regulates IL-10 mediated suppression of TNFα-induced cardiomyocyte apoptosis by upregulating Stat3 phosphorylation. PLoS ONE 2011, 6, e25009. [Google Scholar] [CrossRef] [Green Version]
- Chowdhry, M.F.; Vohra, H.A.; Galinanes, M. Diabetes increases apoptosis and necrosis in both ischemic and nonischemic human myocardium: Role of caspases and poly-adenosine diphosphate-ribose polymerase. J. Thorac Cardiovasc. Surg. 2007, 134, 124–131.e3. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zhang, T.; Dai, H.; Liu, G.; Wang, H.; Sun, Y.; Zhang, Y.; Ge, Z. Involvement of endoplasmic reticulum stress in myocardial apoptosis of streptozocin-induced diabetic rats. J. Clin. Biochem. Nutr. 2007, 41, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Ares-Carrasco, S.; Picatoste, B.; Benito-Martin, A.; Zubiri, I.; Sanz, A.B.; Sanchez-Nino, M.D.; Ortiz, A.; Egido, J.; Tunon, J.; Lorenzo, O. Myocardial fibrosis and apoptosis, but not inflammation, are present in long-term experimental diabetes. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H2109–H2119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izumi, M.; Fujio, Y.; Kunisada, K.; Negoro, S.; Tone, E.; Funamoto, M.; Osugi, T.; Oshima, Y.; Nakaoka, Y.; Kishimoto, T.; et al. Bone morphogenetic protein-2 inhibits serum deprivation-induced apoptosis of neonatal cardiac myocytes through activation of the Smad1 pathway. J. Biol. Chem. 2001, 276, 31133–31141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urbina, P.; Singla, D.K. BMP-7 attenuates adverse cardiac remodeling mediated through M2 macrophages in prediabetic cardiomyopathy. Am. J. Physiol. Heart Circ. Physiol. 2014, 307, H762–H772. [Google Scholar] [CrossRef] [Green Version]
- Kurlawalla-Martinez, C.; Stiles, B.; Wang, Y.; Devaskar, S.U.; Kahn, B.B.; Wu, H. Insulin hypersensitivity and resistance to streptozotocin-induced diabetes in mice lacking PTEN in adipose tissue. Mol. Cell. Biol. 2005, 25, 2498–2510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moroni, F.; Ammirati, E.; Norata, G.D.; Magnoni, M.; Camici, P.G. The Role of Monocytes and Macrophages in Human Atherosclerosis, Plaque Neoangiogenesis, and Atherothrombosis. Mediators Inflamm. 2019, 2019, 11. [Google Scholar] [CrossRef] [Green Version]
- Giannarelli, C.; Fernandez, D.M. Manipulating Macrophage Polarization to Fix the Broken Heart: Challenges and Hopes. J. Am. Coll. Cardiol. 2018, 72, 905–907. [Google Scholar] [CrossRef]
- Nikiforov, N.G.; Wetzker, R.; Kubekina, M.V.; Petukhova, A.V.; Kirichenko, T.V.; Orekhov, A.N. Trained Circulating Monocytes in Atherosclerosis: Ex Vivo Model Approach. Front. Pharmacol. 2019, 10, 725. [Google Scholar] [CrossRef]
- Woollard, K.J.; Geissmann, F. Monocytes in atherosclerosis: Subsets and functions. Nat. Rev. Cardiol. 2010, 7, 77–86. [Google Scholar] [CrossRef]
- Lampiasi, N.; Russo, R.; Zito, F. The Alternative Faces of Macrophage Generate Osteoclasts. BioMed Res. Int. 2016, 2016, 9. [Google Scholar] [CrossRef] [Green Version]
- Zizzo, G.; Cohen, P.L. IL-17 stimulates differentiation of human anti-inflammatory macrophages and phagocytosis of apoptotic neutrophils in response to IL-10 and glucocorticoids. J. Immunol. 2013, 190, 5237–5246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, P.; Clements, V.K.; Ostrand-Rosenberg, S. Interleukin-13-regulated M2 macrophages in combination with myeloid suppressor cells block immune surveillance against metastasis. Cancer Res. 2005, 65, 11743–11751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weis, N.; Weigert, A.; von Knethen, A.; Brune, B. Heme oxygenase-1 contributes to an alternative macrophage activation profile induced by apoptotic cell supernatants. Mol. Biol. Cell. 2009, 20, 1280–1288. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Roos, A.; Schlagwein, N.; Woltman, A.M.; Daha, M.R.; van Kooten, C. IL-10-producing macrophages preferentially clear early apoptotic cells. Blood 2006, 107, 4930–4937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Libby, P.; Ridker, P.M.; Hansson, G.K. Inflammation in atherosclerosis: From pathophysiology to practice. J. Am. Coll. Cardiol. 2009, 54, 2129–2138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galkina, E.; Ley, K. Immune and inflammatory mechanisms of atherosclerosis (*). Annu. Rev. Immunol. 2009, 27, 165–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, K.J.; Tabas, I. Macrophages in the pathogenesis of atherosclerosis. Cell 2011, 145, 341–355. [Google Scholar] [CrossRef] [Green Version]
- Seimon, T.; Tabas, I. Mechanisms and consequences of macrophage apoptosis in atherosclerosis. J. Lipid. Res. 2009, 50, 382–387. [Google Scholar] [CrossRef] [Green Version]
- Tardif, G.; Pelletier, J.P.; Boileau, C.; Martel-Pelletier, J. The BMP antagonists follistatin and gremlin in normal and early osteoarthritic cartilage: An immunohistochemical study. Osteoarthr. Cartil. 2009, 17, 263–270. [Google Scholar] [CrossRef] [Green Version]
- Araki, K.; Ellebedy, A.H.; Ahmed, R. TOR in the immune system. Curr. Opin. Cell. Biol. 2011, 23, 707–715. [Google Scholar] [CrossRef]
- Martin, M.; Schifferle, R.E.; Cuesta, N.; Vogel, S.N.; Katz, J.; Michalek, S.M. Role of the phosphatidylinositol 3 kinase-Akt pathway in the regulation of IL-10 and IL-12 by Porphyromonas gingivalis lipopolysaccharide. J. Immunol. 2003, 171, 717–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Rajasingh, J.; Bord, E.; Luedemann, C.; Asai, J.; Hamada, H.; Thorne, T.; Qin, G.; Goukassian, D.; Zhu, Y.; Losordo, D.W.; et al. IL-10-induced TNF-alpha mRNA destabilization is mediated via IL-10 suppression of p38 MAP kinase activation and inhibition of HuR expression. FASEB J. 2006, 20, 2112–2114. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Types | Alternate Names | Tissues that Express | Functions | Receptors |
|---|---|---|---|---|
| BMP-1 | BMP-1 is a metalloproteinase | major end organs (heart, lung, liver, pancreas, kidney, and brain), lymphoid organs (bone marrow, thymus, spleen and lymph nodes), exocrine glands (prostate and mammary gland) organ protectors (muscle and bone) | Metalloprotease that cleaves COOH–propeptides of procollagens I, II, and III/induces cartilage formation/cleaves BMP antagonist chordin | _____ |
| BMP-2 | BMP-2A, XBMP2, xBMP-2, MGC114605 | major end organs (lung, pancreas, and kidney), lymphoid organ (spleen) | Induces bone and cartilage formation. Plays a role in skeletal repair and regeneration/heart formation | ALK-2, 3, 6 BMPR-II; ActR-IIA, ActR-IIB |
| BMP-3a & 3b | Osteogenin, BMP-3A | major end organs (brain, heart, pancreas), exocrine gland (prostate), organ protector (skeletal muscle), lymphoid organs (bone marrow, spleen and thymus), BMP-3b also expresses in spinal cord | Negative regulator of bone morphogenesis Cell differentiation regulation; skeletal morphogenesis; Regulates cell growth and differentiation in both embryonic and adult tissues | ALK-4 ActR-IIA, ActR-IIB |
| BMP-4 | BMP-2B, BMP2B1, ZYME, OFC11, MCOPS6 | major end organs (brain, heart, pancreas, liver, lung, kidney), exocrine gland (prostate), organ protector (skeletal muscle), lymphoid organs (bone marrow, spleen and thymus), spinal cord | Skeletal repair and regeneration; kidney formation; Induces cartilage and bone formation; limb formation; tooth development. | ALK-2,3,5,6 BMPR-II, ActR-IIA |
| BMP-5 | MGC34244 | major end organs (brain, heart, pancreas, liver, lung, kidney), exocrine gland (prostate), organ protector (skeletal muscle), lymphoid organs (bone marrow, spleen and thymus), spinal cord | Limb development; induces bone and cartilage morphogenesis; connecting soft tissues | ALK-3 BMPR-II; ActR-IIA, ActR-IIB |
| BMP-6 | Vgr1, DVR-6 | major end organs (brain, heart, pancreas, liver, lung, kidney); exocrine gland (prostate); organ protector (muscle and bone), lymphoid organs (bone marrow, spleen and thymus); spinal cord | Cartilage hypertrophy; bone morphogenesis; nervous system development; Plays a role in early development | ALK-2, 3, 6 BMPR-II; ActR-IIA, ActR-IIB |
| BMP-7 | OP-1 | major end organs (brain, heart, pancreas, liver, lung, kidney), exocrine gland (prostate) organ protector (skeletal muscle), lymphoid organs (bone marrow, spleen and thymus), spinal cord. | Skeletal repair and regeneration; kidney and eye formation; nervous system development plays a major role in calcium regulation and bone homeostasis | ALK 2, 3, 6 BMPR-II; |
| BMP-8a & 8b | OP-2, FLJ14351, FLJ45264 OP-3, PC-8, MGC131757 | major end organs (brain, heart, kidney, lung, liver, pancreas), exocrine gland (prostate), organ protector (skeletal muscle), lymphoid organs (spleen, thymus bone marrow) spinal cord | Induces cartilage formation; Bone morphogenesis and spermatogenesis; calcium regulation and bone homeostasis. | ALK 2; 3; 4; 6; 7 BMPR-II; ALK3,6 BMPR-II; ActR-IIA, ActR-IIB |
| BMP-9 | GDF-2 | major end organ (liver) | Bone morphogenesis; cholinergic neurons development; in glucose metabolism; potent inhibitor of angiogenesis | ALK-1,2 BMPR-II; ActR-IIA, ActR-IIB |
| BMP-10 | MGC126783 | major end organs (brain, heart, kidney, lung, liver, pancreas), exocrine gland (prostate), organ protector (skeletal muscle), lymphoid organs (spleen, thymus, bone marrow) spinal cord. | Heart morphogenesis maintains the proliferative activity of embryonic cardiomyocytes by preventing premature activation of the negative cell cycle regulator; inhibits endothelial cell migration and growth | ALK-1, 3, 6 ActR-IIA, ActR-IIB |
| BMP-11 | GDF-11 | major end organs (brain, pancreas), exocrine gland (prostate), lymphoid organs (spleen, thymus bone marrow) spinal cord. | Pattering mesodermal and neural tissues, dentin formation | ALK-3, 4, 5, 7 BMPR-II; ActR-IIA, ActR-IIB |
| BMP-12 | GDF-7, CDMP-3 | _____ | Ligament and tendon development/sensory neuron development | ALK-3, 6 BMPR-II; ActR-IIA |
| BMP-13 | GDF-6, CDMP-2, KFS, KFSL, SGM1, MGC158100, MGC158101 | _____ | Normal formation of bones and joins; skeletal morphogenesis and chondrogenesis Plays a key role in establishing boundaries between skeletal elements during development | ALK-3, 6 BMPR-II; ActR-IIA, ActR-IIB |
| BMP-14 | GDF-5, CDMP-1, OS5, LAP4, SYNS2, MP52 | sensory organs (eye, skin), major end organs (brain, heart; kidney, liver, lung), embryonic tissue, mixed connective tissue, pituitary gland, salivary gland; exocrine gland (prostate), reproductive system related (uterus), lymphoid organ (bone marrow) | Bone and cartilage formation; Skeletal repair and regeneration | ALK-3, 6 BMPR-II; ActR-IIA |
| BMP-15 | GDF-9B, ODG2, POF4 | _______ | Oocyte and follicular development | ALK-6 |
| BMP-16 | _____ | embryonic tissue; reproductive system (testis) | Skeletal repair and regeneration Essential for mesoderm formation and axial patterning during embryonic development | _____ |
| BMP-17 | _____ | major end organ (brain, lung, liver, pancreas, spleen) lymphoid organ (lymph node); exocrine gland (mammary gland); sensory organ (skin); reproductive organ (testis); bladder; embryonic tissue; intestine; joints; | Required for left-right axis determination as a regulator of LEFTY2 and NODAL | _____ |
| BMP-18 | _____ | major end organ (brain), embryonic tissue, reproductive system (testis) | Required for left-right (L-R) asymmetry determination of organ systems in mammals. May play a role in endometrial bleeding | _____ |
| Inhibitors | Name | Role |
|---|---|---|
| Neuroblastoma Dan family | DAN | plays a role in tumor suppression; cell proliferation |
| PRDC/GRem2 (protein related to DAN and Cerberus) | PRDC is a secreted, cysteine knot-containing BMP antagonist; play a role in regulation of BMP signaling in ovary, brain, and other adult tissues | |
| Gremlin | higher level expression in Basal cell carcinoma stromal cells; promotes proliferation and tumor growth; induces cell cycle progression via p21; interacts directly with target endothelial cells; acts as a proangiogenic factor to regulate angiogenesis; blocks osteoblast differentiation and function by blocking BMP signaling | |
| Cerberus/Cer1 | anterior neural induction and somite formation during embryogenesis; regulate Nodal signaling during gastrulation as well as the formation and patterning of the primitive streak Blocks Nodal, BMP, and Wnt signaling | |
| Coco/Dand5 | antagonizes NODAL and BMP4 signaling during development, organogenesis, tissue growth and differentiation; Blocks BMP/TGF-β and Wnt signaling | |
| Caronte | antagonizing symmetrically expressed BMP signals | |
| USAG-1 | BMP and Wnt antagonist during the development of kidney, tooth, and mammary tissues | |
| Sclerostin/ SOST | endogenous antagonist of the Wnt/β-catenin pathway in the regulation of bone mass; acts as negative regulator of bone formation | |
| Dante/Dte | plays potential role during early stages of mouse embryonic development; inhibit BMP signaling | |
| Chordin family | Chordin, | functions as BMP antagonist that blocks BMP activity by binding to the BMPs and inhibiting their interaction with their receptors |
| Ventroptin/ Chordin-like-1/ Neuralin 1 | regulates retinal angiogenesis via modulation of BMP4 actions in endothelial cells. | |
| Chordin-like-2, | prevents the binding of BMPs to type 1 and type 2 receptors as well as BMP-induced cellular responses; reduces the rate of matrix deposition by mesenchymal cells, acts as a negative regulator of cartilage formation. | |
| Kielin | enhances BMP signaling in a paracrine way; inhibits both the activin-A and TGFB1-mediated signaling pathways | |
| Nell | promotes the osteogenic differentiation of adipose-derived stromal/stem cells and inhibits adipogenic differentiation. Binding of NELL1 to Integrin beta 1 was shown to be critical for its role in promoting osteogenic differentiation and adhesion to the extracellular matrix. | |
| Crossveinless2 | bone morphogenetic protein-binding endothelial cell precursor-derived regulator (BMPER). Secreted CV-2 interacts with BMP and inhibits its function | |
| Brorin | Brorin binds and antagonizes BMPs, interacting via the von Willebrand factor C domain. It promotes neurogenesis in mouse neural precursors | |
| Noggin | promotes skin tumorigenesis; reduces tumor size and decreases bone loss compared to untreated control animals; suppresses BMP4 induction of vascular endothelial growth factor receptor 2 in embryonic blood vessels; inhibits BMP interaction with their receptors | |
| Follistatin | acts as a modulator of gonadal tumor progression and the activin stimulated wasting syndrome; inhibits BMP interaction with their receptors | |
| Twisted gastrulation | regulates the extracellular availability of a mesoderm inducer, BMP 4 As agonist-enhances cleavage of BMP/chordin complex by BMP1/tolloid (releasing free BMP) Required to specify the dorsal-most structures in embryo. | |
| Follistatin-related gene (FLRG) | acts as activin antagonist and inhibits tumor cell growth |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aluganti Narasimhulu, C.; Singla, D.K. The Role of Bone Morphogenetic Protein 7 (BMP-7) in Inflammation in Heart Diseases. Cells 2020, 9, 280. https://doi.org/10.3390/cells9020280
Aluganti Narasimhulu C, Singla DK. The Role of Bone Morphogenetic Protein 7 (BMP-7) in Inflammation in Heart Diseases. Cells. 2020; 9(2):280. https://doi.org/10.3390/cells9020280
Chicago/Turabian StyleAluganti Narasimhulu, Chandrakala, and Dinender K Singla. 2020. "The Role of Bone Morphogenetic Protein 7 (BMP-7) in Inflammation in Heart Diseases" Cells 9, no. 2: 280. https://doi.org/10.3390/cells9020280
APA StyleAluganti Narasimhulu, C., & Singla, D. K. (2020). The Role of Bone Morphogenetic Protein 7 (BMP-7) in Inflammation in Heart Diseases. Cells, 9(2), 280. https://doi.org/10.3390/cells9020280