α2-Adrenergic Receptor in Liver Fibrosis: Implications for the Adrenoblocker Mesedin


 ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures
2.2. Animal Models
2.3. RNA Isolation and Quantification
2.4. Western Blot Analysis
2.5. Immunofluorescence Analyses
2.6. Statistical Analyses
3. Results
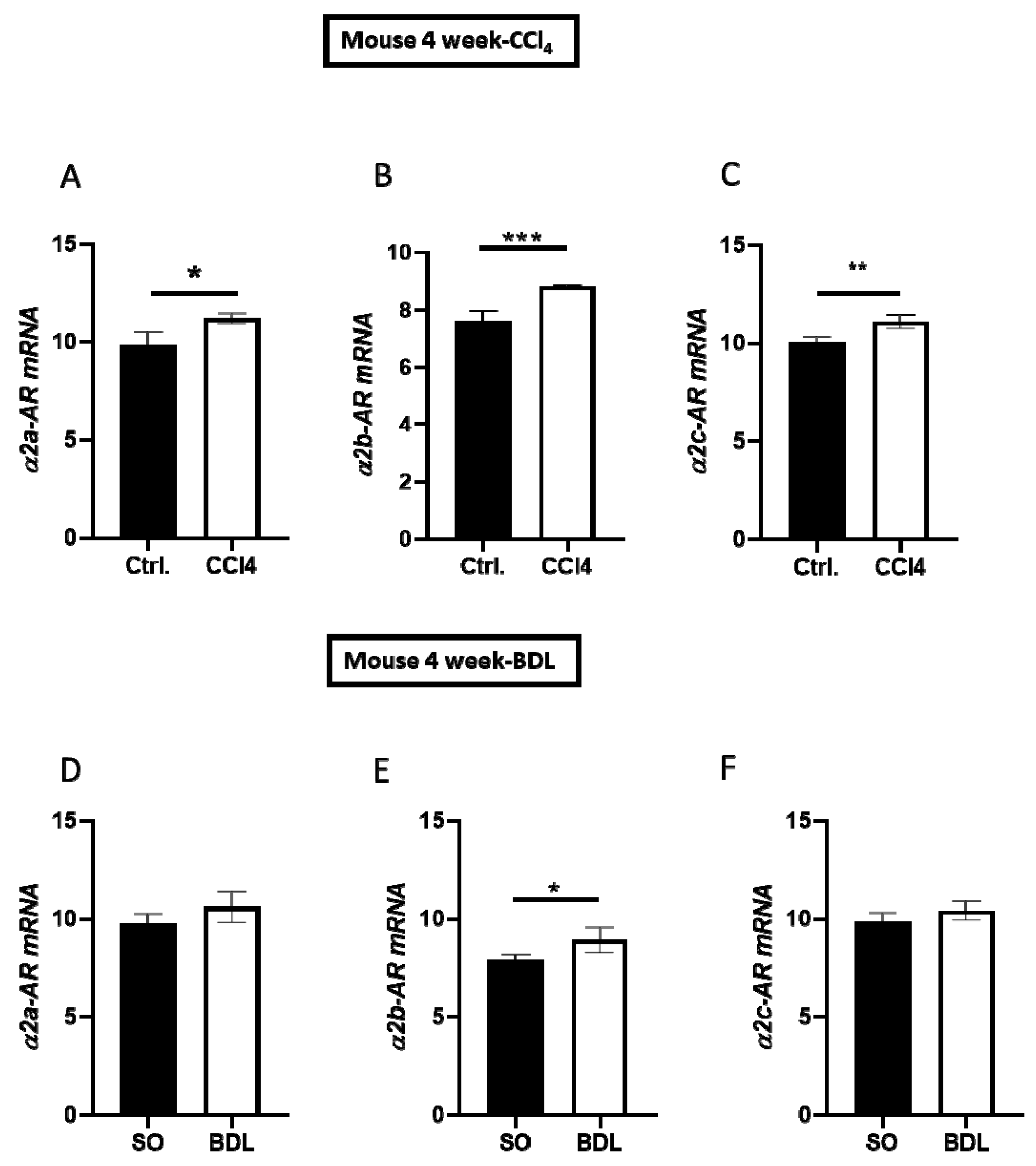
3.1. α2 Receptors are Upregulated in Fibrotic/Cirrhotic Livers
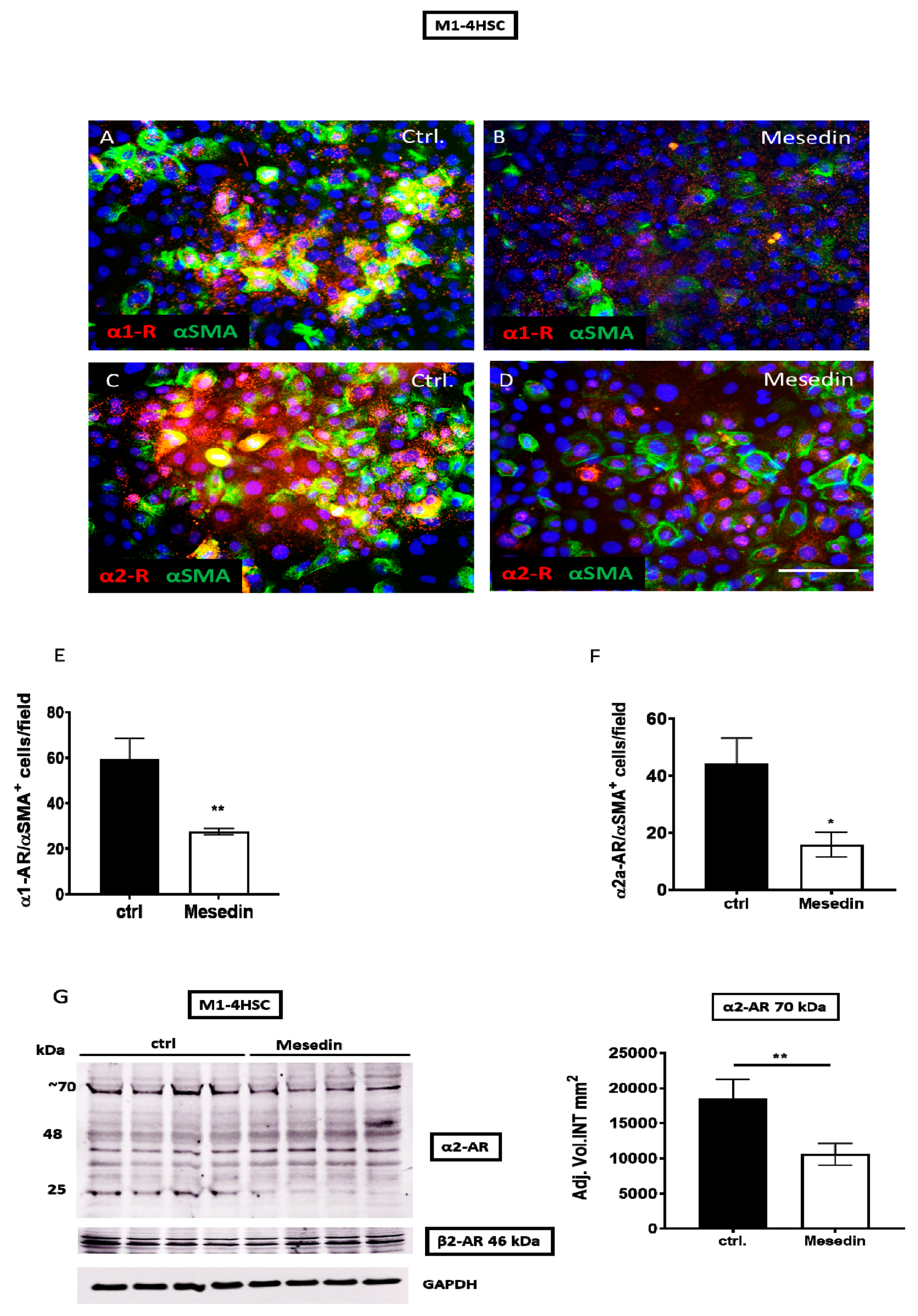
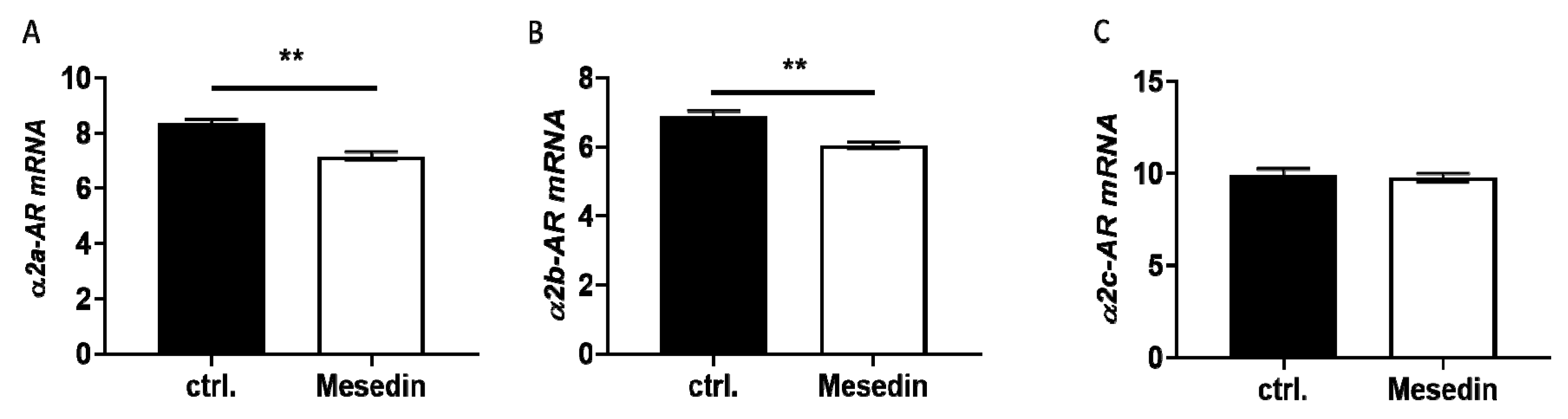
3.2. Mesedin Decreased the Expression of α1, α2a, and α2b Receptors in HSCs
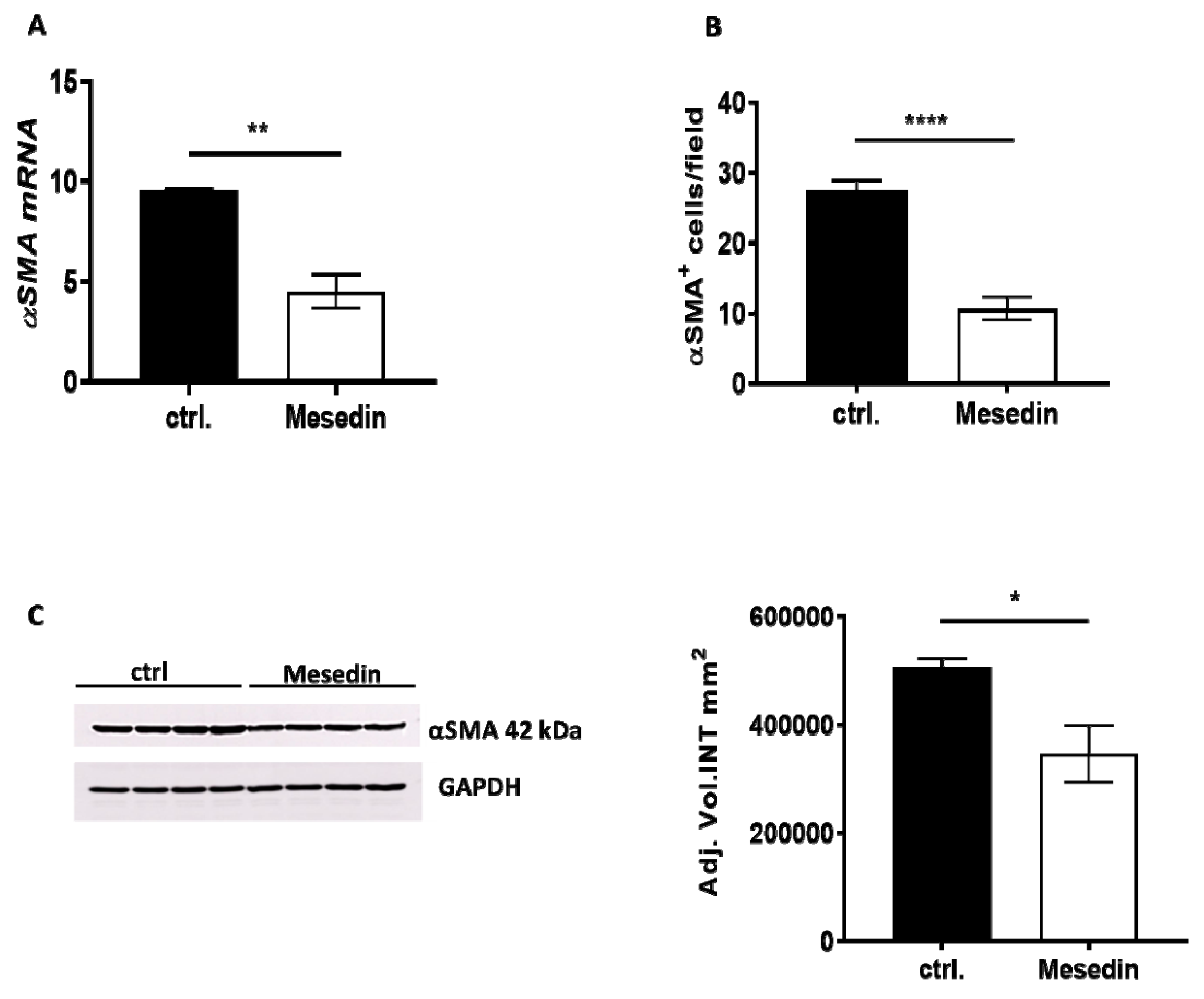
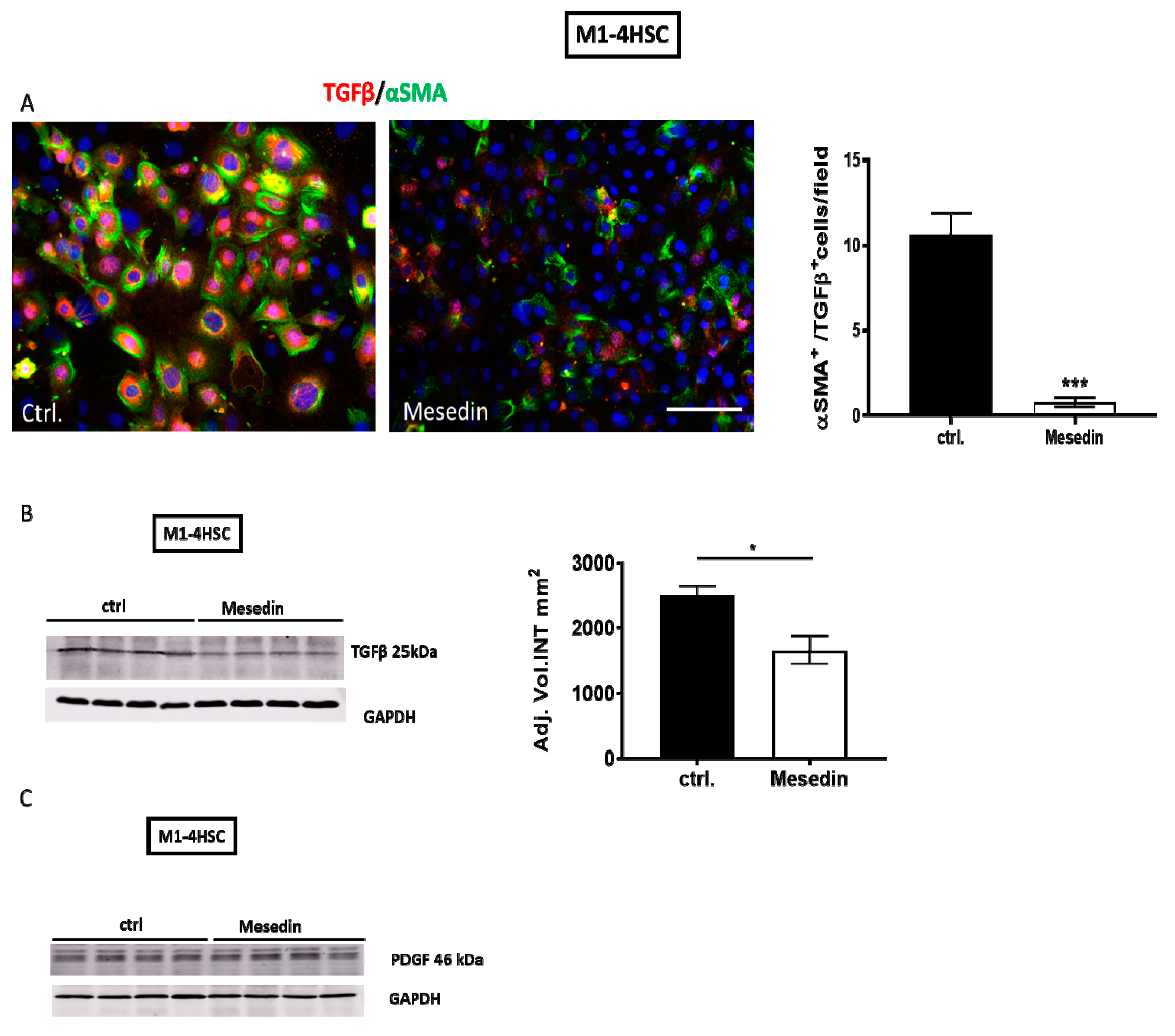
3.3. Antifibrotic Effects of Mesedin
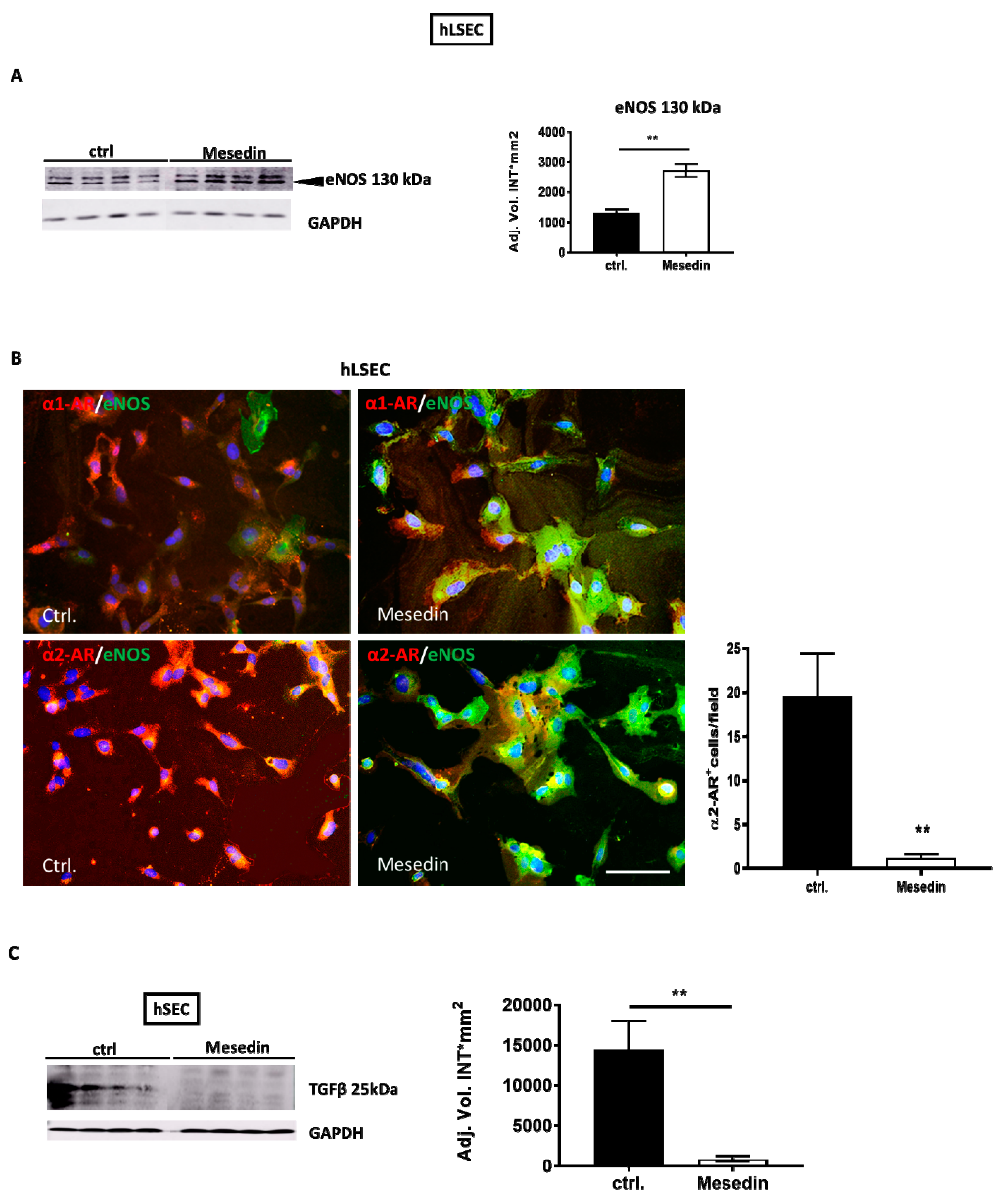
3.4. Influence of the α2 Blockade on Permeability Marker Expression in hLSECs
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Henriksen, J.H.; Møller, S.; Ring-Larsen, H.; Christensen, N.J. The sympathetic nervous system in liver disease. J. Hepatol. 1998, 29, 328–341. [Google Scholar] [CrossRef]
- Oben, J.A.; Roskams, T.; Yang, S.; Lin, H.; Sinelli, N.; Torbenson, M.; Smedh, U.; Moran, T.H.; Li, Z.; Huang, J.; et al. Hepatic fibrogenesis requires sympathetic neurotransmitters. Gut 2004, 53, 438–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oben, J.A.; Roskams, T.; Yang, S.; Lin, H.; Sinelli, N.; Li, Z.; Torbenson, M.; Thomas, S.A.; Diehl, A.M. Norepinephrine induces hepatic fibrogenesis in leptin deficient ob/ob mice. Biochem. Biophys. Res. Commun. 2003, 308, 284–292. [Google Scholar] [CrossRef]
- Miksa, M.; Das, P.; Zhou, M.; Wu, R.; Dong, W.; Ji, Y.; Goyert, S.M.; Ravikumar, T.S.; Wang, P. Pivotal role of the α2A-adrenoceptor in producing inflammation and organ injury in a rat model of sepsis. PLoS ONE 2009, 4, e5504. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, B.B.; Dukes, D.F.; Lefkowitz, R.J. Alpha-adrenergic receptors in liver membranes: Delineation with subtype selective radioligands. Life Sci. 1981, 28, 265–272. [Google Scholar] [CrossRef]
- Bylund, D.B. Subtypes of alpha-1 and alpha-2 adrenergic receptors. Eur. Neuropsychopharmacol. 1992, 2, 229–230. [Google Scholar] [CrossRef]
- Zhou, M.; Yang, S.; Koo, D.J.; Ornan, D.A.; Chaudry, I.H.; Wang, P. The role of Kupffer cell α2-adrenoceptors in norepinephrine-induced TNF-α production. Biochim. Biophys. Acta-Mol. Basis Dis. 2001, 1537, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Xuanfei, L.; Hao, C.; Zhujun, Y.; Yanming, L.; Jianping, G. Imidazoline I2 receptor inhibitor idazoxan regulates the progression of hepatic fibrosis via Akt-Nrf2-Smad2/3 signaling pathway. Oncotarget 2017, 8, 21015–21030. [Google Scholar] [CrossRef] [Green Version]
- Tananyan, A.; Balasanyan, M.P. 4.c.008 Prevention of focal ischemia induced memory deficit and anxiety by mesedin. Eur. Neuropsychopharmacol. 2014, 24, S610. [Google Scholar] [CrossRef]
- Tananyan, A.G.; Balasanyan, M.G.; Baykov, A.V.; Hovsepyan, L.M.; Ghazaryan, G.S. The effect of mesedin on the content of oxidative stress biomarkers in the brain tissue in ischemia. Neurochem. J. 2019, 13, 68–72. [Google Scholar] [CrossRef]
- Melkonyan, M.M.; Hunanyan, L.; Lourhmati, A.; Layer, N.; Beer-Hammer, S.; Yenkoyan, K.; Schwab, M.; Danielyan, L. Neuroprotective, neurogenic, and amyloid beta reducing effect of a novel alpha 2-Adrenoblocker, mesedin, on astroglia and neuronal progenitors upon hypoxia and glutamate exposure. Int. J. Mol. Sci. 2018, 19, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.C.; Peng, Y.J.; Wang, S.Y.; Lai, M.J.; Young, T.H.; Salter, D.M.; Lee, H.S. Sympathetic nervous system control of carbon tetrachloride-induced oxidative stress in liver through α-adrenergic signaling. Oxid. Med. Cell. Longev. 2016, 2016, 3190617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigala, B.; McKee, C.; Soeda, J.; Pazienza, V.; Morgan, M.; Lin, C.I.; Selden, C.; Vander Borght, S.; Mazzoccoli, G.; Roskams, T.; et al. Sympathetic nervous system catecholamines and neuropeptide Y neurotransmitters are upregulated in human NAFLD and modulate the fibrogenic function of hepatic stellate cells. PLoS ONE 2013, 8, e72928. [Google Scholar] [CrossRef] [PubMed]
- Sha, J.; Zhang, H.; Zhao, Y.; Feng, X.; Hu, X.; Wang, C.; Song, M.; Fan, H. Dexmedetomidine attenuates lipopolysaccharide-induced liver oxidative stress and cell apoptosis in rats by increasing GSK-3β/MKP-1/Nrf2 pathway activity via the α2 adrenergic receptor. Toxicol. Appl. Pharmacol. 2019, 364, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Vartanyan, S.O.; Avakyan, A.S.; Sargsyan, A.B.; Arutyunyan, S.A.; Noravyan, O.S.; Tsatinyan, A.S. Synthesis and biologic properties of new thiazolylbenzodioxane derivatives. Russ. J. Org. Chem. 2016, 52, 244–248. [Google Scholar] [CrossRef]
- Garcia-Tsao, G. Beta blockers in cirrhosis: The window re-opens. J. Hepatol. 2016, 64, 532–534. [Google Scholar] [CrossRef] [Green Version]
- Borkham-Kamphorst, E.; van de Leur, E.; Zimmermann, H.W.; Karlmark, K.R.; Tihaa, L.; Haas, U.; Tacke, F.; Berger, T.; Mak, T.W.; Weiskirchen, R. Protective effects of lipocalin-2 (LCN2) in acute liver injury suggest a novel function in liver homeostasis. Biochim. Biophys. Acta-Mol. Basis Dis. 2013, 1832, 660–673. [Google Scholar] [CrossRef] [Green Version]
- Tag, C.; Weiskirchen, S.; Hittatiya, K.; Tacke, F.; Tolba, R.; Weiskirchen, R. Induction of experimental obstructive cholestasis in mice. Lab. Anim. 2015, 49, 70–80. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Proell, V.; Mikula, M.; Fuchs, E.; Mikulits, W. The plasticity of p19ARF null hepatic stellate cells and the dynamics of activation. Biochim. Biophys. Acta-Mol. Cell Res. 2005, 1744, 76–87. [Google Scholar] [CrossRef] [Green Version]
- De Salamanca, A.E.; Siemasko, K.F.; Diebold, Y.; Calonge, M.; Gao, J.; Juárez-Campo, M.; Stern, M.E. Expression of muscarinic and adrenergic receptors in normal human conjunctival epithelium. Investig. Ophthalmol. Vis. Sci. 2005, 46, 504–513. [Google Scholar] [CrossRef] [PubMed]
- Deleve, L.D. Liver sinusoidal endothelial cells in hepatic fibrosis. Hepatology 2015, 61, 1740–1746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, M.; Nishikawa, Y.; Omori, Y.; Yoshioka, T.; Tokairin, T.; McCourt, P.; Enomoto, K. Involvement of signaling of VEGF and TGF-β in differentiation of sinusoidal endothelial cells during culture of fetal rat liver cells. Cell Tissue Res. 2007, 329, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Chen, L.; Leng, X.; Du, R. Expression of alpha1 adrenoceptor subtypes mRNA in hepatic tissues of cirrhotic patients with portal hypertension. Zhonghua Wai Ke Za Zhi 2000, 38, 325–328. [Google Scholar]
- Trebicka, J.; Hennenberg, M.; Pröbsting, A.S.; Laleman, W.; Klein, S.; Granzow, M.; Nevens, F.; Zaagsma, J.; Heller, J.; Sauerbruch, T. Role of β3-adrenoceptors for intrahepatic resistance and portal hypertension in liver cirrhosis. Hepatology 2009, 50, 1924–1935. [Google Scholar] [CrossRef]
- Muñoz-Ortega, M.H.; Llamas-Ramírez, R.W.; Romero-Delgadillo, N.I.; Elías-Flores, T.G.; De Jesus Tavares-Rodríguez, E.; Del Rosario Campos-Esparza, M.; Cervantes-García, D.; Muñoz-Fernández, L.; Gerardo-Rodríguez, M.; Ventura-Juárez, J. Doxazosin treatment attenuates carbon tetrachloride-induced liver fibrosis in hamsters through a decrease in transforming growth factor β secretion. Gut Liver 2016, 10, 101–108. [Google Scholar] [CrossRef]
- Fabregat, I.; Moreno-Càceres, J.; Sánchez, A.; Dooley, S.; Dewidar, B.; Giannelli, G.; ten Dijke, P. TGF-β signalling and liver disease. FEBS J. 2016, 2219–2232. [Google Scholar] [CrossRef] [Green Version]
- Briest, W.; Homagk, L.; Raßler, B.; Ziegelhoffer-Mihalovicova, B.; Meier, H.; Tannapfel, A.; Leiblein, S.; Saalbach, A.; Deten, A.; Zimmer, H.G. Norepinephrine-induced changes in cardiac transforming growth factor-β isoform expression pattern of female and male rats. Hypertension 2004, 44, 410–418. [Google Scholar] [CrossRef] [Green Version]
- Medina, L.D.C.; Vazquez-Prado, J.; Garcia-SHIRSCH-ERNSTainz, J.A. Cross-talk between receptors with intrinsic tyrosine kinase activity and alpha1b-adrenoceptors [In Process Citation] 154 154. Biochem. J. 2000, 350 Pt 2, 413–419. [Google Scholar] [CrossRef]
- Nozik-Grayck, E.; Whalen, E.J.; Stamler, J.S.; McMahon, T.J.; Chitano, P.; Piantadosi, C.A. S-nitrosoglutathione inhibits α1-adrenergic receptor-mediated vasoconstriction and ligand binding in pulmonary artery. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2006, 290. [Google Scholar] [CrossRef]
- Wang, P.; Tait, S.M.; Chaudry, I.H. Sustained elevation of norepinephrine depresses hepatocellular function. Biochim. Biophys. Acta-Mol. Basis Dis. 2000, 1535, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Ueta, C.B.; Fernandes, G.W.; Capelo, L.P.; Fonseca, T.L.; Maculan, F.D.A.; Gouveia, C.H.A.; Brum, P.C.; Christoffolete, M.A.; Aoki, M.S.; Lancellotti, C.L.; et al. β1 Adrenergic receptor is key to cold-and diet-induced thermogenesis in mice. J. Endocrinol. 2012, 214, 359–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vairappan, B. Endothelial dysfunction in cirrhosis: Role of inflammation and oxidative stress. World J. Hepatol. 2015, 7, 443–459. [Google Scholar] [CrossRef] [PubMed]
- Bosch, J.; Abraldes, J.G.; Fernández, M.; García-Pagán, J.C. Hepatic endothelial dysfunction and abnormal angiogenesis: New targets in the treatment of portal hypertension. J. Hepatol. 2010, 53, 558–567. [Google Scholar] [CrossRef] [Green Version]
- Duong, H.T.T.; Dong, Z.; Su, L.; Boyer, C.; George, J.; Davis, T.P.; Wang, J. The use of nanoparticles to deliver nitric oxide to hepatic stellate cells for treating liver fibrosis and portal hypertension. Small 2015, 11, 2291–2304. [Google Scholar] [CrossRef]
- Angus, J.A.; Cocks, T.M.; Satoh, K. The α adrenoceptors on endothelial cells. Fed. Proc. 1986, 45, 2355–2359. [Google Scholar]
- Chen, Y.; Luan, L.; Wang, C.; Song, M.; Zhao, Y.; Yao, Y.; Yang, H.; Ma, B.; Fan, H. Dexmedetomidine protects against lipopolysaccharide-induced early acute kidney injury by inhibiting the iNOS/NO signaling pathway in rats. Nitric Oxide-Biol. Chem. 2019, 85, 1–9. [Google Scholar] [CrossRef]
- Vanhoutte, P.M. Endothelial adrenoceptors. J. Cardiovasc. Pharmacol. 2001, 38, 796–808. [Google Scholar] [CrossRef]
- Rao, M.Y.; Raghu, J.; Deshmukh, S.; Amaravathi, K.S.; Sudhir, U. Arterial hypoxemia in patients with cirrhosis of liver. J. Assoc. Physicians India 2008, 56, 681–684. [Google Scholar]
- Mesarwi, O.A.; Shin, M.K.; Bevans-Fonti, S.; Schlesinger, C.; Shaw, J.; Polotsky, V.Y. Hepatocyte hypoxia inducible factor-1 mediates the development of liver fibrosis in a mouse model of nonalcoholic fatty liver disease. PLoS ONE 2016, 11, e0168572. [Google Scholar] [CrossRef] [Green Version]
- Abd-Elgawad, H.; Abu-Elsaad, N.; El-Karef, A.; Ibrahim, T. Piceatannol increases the expression of hepatocyte growth factor and IL-10 thereby protecting hepatocytes in thioacetamide-induced liver fibrosis. Can. J. Physiol. Pharmacol. 2016, 94, 779–787. [Google Scholar] [CrossRef]
- Huang, Y.-H.; Chen, M.-H.; Guo, Q.-L.; Chen, Z.-X.; Chen, Q.-D.; Wang, X.-Z. Interleukin-10 induces senescence of activated hepatic stellate cells via STAT3-p53 pathway to attenuate liver fibrosis. Cell. Signal. 2020, 66, 109445. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Alias | Gene Symbol | RefSeq | Assay ID |
|---|---|---|---|
| α-SMA | Acta2 | NM_007392.3 | Mm01204962_gH |
| α2a-AR | Adra2a | NM_007417.4 | Mm00845383_s1 |
| α2b-AR | Adra2b | NM_009633.3 | Mm00477390_s1 |
| α2c-AR | Adra2c | NM_007418.3 | Mm00431686_s1 |
| GAPDH | Gapdh | NM_008084.3; NM_001289726.1 | Mm99999915_g1 |
| Antibody | Catalog Number | Supplier |
|---|---|---|
| α-SMA (Western) (rabbit monoclonal) | ab124964 | abcam, Cambridge, UK |
| α-SMA (IHC) (mouse monoclonal) | 61001 | Progen, Heidelberg, Germany |
| TGF-β (rabbit polyclonal) | ab92486 | abcam |
| α2a-AR (rabbit polyclonal) | SAB4500548 | Sigma-Aldrich, Taufkirchen, Germany |
| α1-AR (rabbit polyclonal) | ab3462 | abcam |
| eNOS (rabbit monoclonal) | #32027 | Cell Signaling Technology, Frankfurt am Main, Germany |
| PDGF-AA (rabbit polyclonal) | ab135881 | abcam |
| Goat antimouse IgG Fluorescein isothiocyanate (FITC)-conjugated (polyclonal) | 115-095-003 | Jackson ImmunoResearch, Ely, UK |
| Goat antirabbit IgG Cy3-conjugated (polyclonal) | 111-165-144 | Jackson ImmunoResearch |
| Goat antirabbit Cy3-conjugated (monoclonal) | 28901106V | GE Healthcare UK Ltd., Amersham, Little Chalfont, UK |
| Goat antimouse Cy5-conjugated (monoclonal) | PA45009V | GE Healthcare UK Ltd. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schwinghammer, U.A.; Melkonyan, M.M.; Hunanyan, L.; Tremmel, R.; Weiskirchen, R.; Borkham-Kamphorst, E.; Schaeffeler, E.; Seferyan, T.; Mikulits, W.; Yenkoyan, K.; et al. α2-Adrenergic Receptor in Liver Fibrosis: Implications for the Adrenoblocker Mesedin. Cells 2020, 9, 456. https://doi.org/10.3390/cells9020456
Schwinghammer UA, Melkonyan MM, Hunanyan L, Tremmel R, Weiskirchen R, Borkham-Kamphorst E, Schaeffeler E, Seferyan T, Mikulits W, Yenkoyan K, et al. α2-Adrenergic Receptor in Liver Fibrosis: Implications for the Adrenoblocker Mesedin. Cells. 2020; 9(2):456. https://doi.org/10.3390/cells9020456
Chicago/Turabian StyleSchwinghammer, Ute A., Magda M. Melkonyan, Lilit Hunanyan, Roman Tremmel, Ralf Weiskirchen, Erawan Borkham-Kamphorst, Elke Schaeffeler, Torgom Seferyan, Wolfgang Mikulits, Konstantin Yenkoyan, and et al. 2020. "α2-Adrenergic Receptor in Liver Fibrosis: Implications for the Adrenoblocker Mesedin" Cells 9, no. 2: 456. https://doi.org/10.3390/cells9020456
APA StyleSchwinghammer, U. A., Melkonyan, M. M., Hunanyan, L., Tremmel, R., Weiskirchen, R., Borkham-Kamphorst, E., Schaeffeler, E., Seferyan, T., Mikulits, W., Yenkoyan, K., Schwab, M., & Danielyan, L. (2020). α2-Adrenergic Receptor in Liver Fibrosis: Implications for the Adrenoblocker Mesedin. Cells, 9(2), 456. https://doi.org/10.3390/cells9020456