Decoding the Role of Interleukin-30 in the Crosstalk between Cancer and Myeloid Cells

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. IL-30 as a Self-Standing Cytokine or Cytokine-Like Subunit Paired with Soluble Receptor-Like Proteins to Form Different Heterodimeric Complexes
2.1. IL-27 Involvement in Cancer-Myeloid Cell Crosstalk
2.2. IL-30/CLF
2.3. IL-30/IL-12p40
2.4. EBI3, IL-35, and IL-39 Involvement in Cancer-Myeloid Cell Crosstalk
2.5. IL-30 Immunobiology in Man and Mouse
3. IL-30 promotes the Expression of Myeloid Cell Growth and Chemotactic Factors in Cancer Cells
4. Tumor- and Myeloid Cell-Derived IL-30 Contributes to the Maintenance of the Cancer Stem Cell Niche
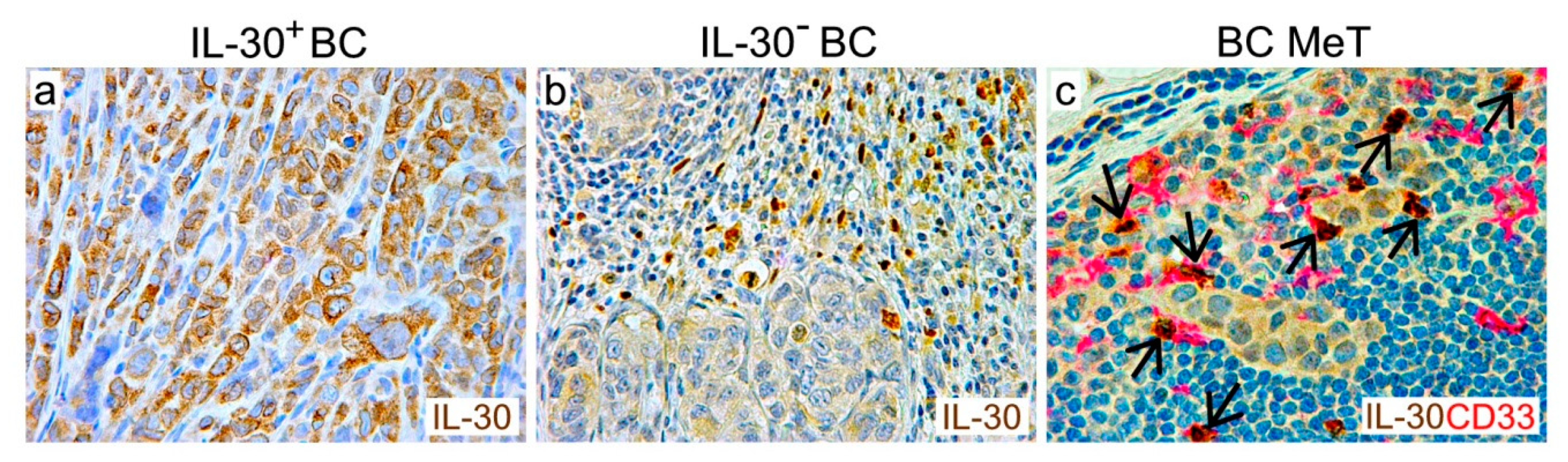
- IL-30 boosts PC–SLC expression of the KIT-ligand, TLR3, Myeloid differentiation primary response 88 (Myd88), Foxp3, and CD274/PD-L1 [82], which counteract the host’s anti-tumor immune response, and expression of chemokine C-C motif ligands 4 (CCL4), CSF2, CSF3, C-X-C motif ligand 1 (CXCL1) and 2 (CXCL2), prostaglandin-endoperoxide synthase 2 (PTGS2), also known as cyclooxygenase-2 (COX2), which along with IL-1β, IL-6, and TNFα [81], promote myeloid cell accumulation and suppressive functions [87]. In fact, the fast-growing tumors that develop from IL-30-overexpressing PC–SLCs, show a prominent myeloid cell infiltrate that mainly consists of F4/80+ macrophages, CD11b+ myeloid cells, and Ly-6G+ granulocytes, which along with tumor progression is hampered by IL-30 knockdown [81];
- IL-30 promotes ‘epithelial–immune cell-like transition’ [93] of PC–SLCs, mainly via STAT1/STAT3 pathways, through induction/up-regulation of expression of the chemokine receptors C-C chemokine receptor type 1 (CCR1), C-X-C motif chemokine receptor 1, 4, and 5 (CXCR1, CXCR4, and CXCR5). Specifically, IL-30 promotes PC–SLC spread to the bone marrow, by boosting local production of CXCL13, and cancer cell expression of CXCR5, which is suppressed by IL-30-silencing [81]. IL-30′s involvement in bone marrow colonization of PC–SLCs suggest that targeting of this cytokine can impact on bone metastasis, which is a crucial step in cancer progression and a leading cause of worsening outcome.
- IL-30 overproduction by PC–SLCs also promotes lung metastasis, involving the CXCR4/CXCL12 axis, since it induces CXCR4 expression on PC–SLCs and promotes their migration towards CXCL12+ pleural covering and bronchiolar walls. More than 80% of mice bearing IL-30 overexpressing tumors developed lung metastasis, versus 46–52% of mice bearing control tumors. Of note, once again, myeloid cells are a critical component of IL-30 conditioned microenvironment, since lung metastasis developed from IL-30 overexpressing tumors reveal a content of F4/80+, CD11b+, and Ly-6G+ cells that are significantly higher than the controls, in association with a prominent metastatic cell proliferation and vascularization [81]. Figure 3 summarizes the effects of IL-30 expression by cancer and myeloid cells on the TME, antitumor immune response, and tumor behavior.
5. Concluding Remarks and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Greten, F.R.; Grivennikov, S.I. Inflammation and Cancer: Triggers, Mechanisms, and Consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, N.A.; Sanchez-Salas, R.; Peske, J.D.; Vano, Y.; Becht, E.; Petitprez, F.; Validire, P.; Ingels, A.; Cathelineau, X.; Fridman, W.H.; et al. The clinical role of the TME in solid cancer. Br. J. Cancer 2019, 120, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Mittal, D.; Gubin, M.M.; Schreiber, R.D.; Smyth, M.J. New insights into cancer immunoediting and its three component phases—Elimination, equilibrium and escape. Curr. Opin. Immunol. 2014, 27, 16–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, X.; Tang, Y.; Hua, S. Immunological Approaches towards Cancer and Inflammation: A Cross Talk. Front. Immunol. 2018, 9, 563. [Google Scholar] [CrossRef] [Green Version]
- Ritter, B.; Greten, F.R. Modulating inflammation for cancer therapy. J. Exp. Med. 2019, 216, 1234–1243. [Google Scholar] [CrossRef]
- Becht, E.; Giraldo, N.A.; Dieu-Nosjean, M.-C.; Sautès-Fridman, C.; Fridman, W.H. Cancer immune contexture and immunotherapy. Curr. Opin. Immunol. 2016, 39, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Solinas, G.; Germano, G.; Mantovani, A.; Allavena, P. Tumor-associated macrophages (TAM) as major players of the cancer-related inflammation. J. Leukoc. Biol. 2009, 86, 1065–1073. [Google Scholar] [CrossRef] [Green Version]
- Tcyganov, E.; Mastio, J.; Chen, E.; Gabrilovich, D.I. Plasticity of myeloid-derived suppressor cells in cancer. Curr. Opin. Immunol. 2018, 51, 76–82. [Google Scholar] [CrossRef]
- Jahchan, N.S.; Mujal, A.M.; Pollack, J.L.; Binnewies, M.; Sriram, V.; Reyno, L.; Krummel, M.F. Tuning the Tumor Myeloid Microenvironment to Fight Cancer. Front. Immunol. 2019, 10, 1611. [Google Scholar] [CrossRef] [Green Version]
- Prima, V.; Kaliberova, L.N.; Kaliberov, S.; Curiel, D.T.; Kusmartsev, S. COX2/mPGES1/PGE2 pathway regulates PD-L1 expression in tumor-associated macrophages and myeloid-derived suppressor cells. Proc. Natl. Acad. Sci. USA 2017, 114, 1117–1122. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Smyth, M.J. Myeloid immunosuppression and immune checkpoints in the tumor microenvironment. Cell. Mol. Immunol. 2020, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ye, Y.; Liu, P.; Yu, W.; Wei, F.; Li, H.; Yu, J. Suppression of T cells by myeloid-derived suppressor cells in cancer. Hum. Immunol. 2017, 78, 113–119. [Google Scholar] [CrossRef]
- Groth, C.; Hu, X.; Weber, R.; Fleming, V.; Altevogt, P.; Utikal, J.; Umansky, V. Immunosuppression mediated by myeloid-derived suppressor cells (MDSCs) during tumour progression. Br. J. Cancer 2019, 120, 16–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pflanz, S.; Timans, J.C.; Cheung, J.; Rosales, R.; Kanzler, H.; Gilbert, J.; Hibbert, L.; Churakova, T.; Travis, M.; Vaisberg, E.; et al. IL-27, a heterodimeric cytokine composed of EBI3 and p28 protein, induces proliferation of naive CD4+ T cells. Immunity 2002, 16, 779–790. [Google Scholar] [CrossRef] [Green Version]
- Di Meo, S.; Airoldi, I.; Sorrentino, C.; Zorzoli, A.; Esposito, S.; Di Carlo, E. Interleukin-30 expression in prostate cancer and its draining lymph nodes correlates with advanced grade and stage. Clin. Cancer Res. 2014, 20, 585–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Airoldi, I.; Cocco, C.; Sorrentino, C.; Angelucci, D.; Di Meo, S.; Manzoli, L.; Esposito, S.; Ribatti, D.; Bertolotto, M.; Iezzi, L.; et al. Interleukin-30 Promotes Breast Cancer Growth and Progression. Cancer Res. 2016, 76, 6218–6229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosmann, M.; Ward, P.A. Modulation of inflammation by interleukin-27. J. Leukoc. Biol. 2013, 94, 1159–1165. [Google Scholar] [CrossRef] [Green Version]
- Curran, M.A.; Geiger, T.L.; Montalvo, W.; Kim, M.; Reiner, S.L.; Al-Shamkhani, A.; Sun, J.C.; Allison, J.P. Systemic 4-1BB activation induces a novel T cell phenotype driven by high expression of Eomesodermin. J. Exp. Med. 2013, 210, 743–755. [Google Scholar] [CrossRef] [Green Version]
- Dibra, D.; Cutrera, J.J.; Li, S. Coordination between TLR9 signaling in macrophages and CD3 signaling in T cells induces robust expression of IL-30. J. Immunol. 2012, 188, 3709–3715. [Google Scholar] [CrossRef]
- Van Seventer, J.M.; Nagai, T.; van Seventer, G.A. Interferon-beta differentially regulates expression of the IL-12 family members p35, p40, p19 and EBI3 in activated human dendritic cells. J. Neuroimmunol. 2002, 133, 60–71. [Google Scholar] [CrossRef]
- Remoli, M.E.; Gafa, V.; Giacomini, E.; Severa, M.; Lande, R.; Coccia, E.M. IFN-beta modulates the response to TLR stimulation in human DC: Involvement of IFN regulatory factor-1 (IRF-1) in IL-27 gene expression. Eur. J. Immunol. 2007, 37, 3499–3508. [Google Scholar] [CrossRef] [PubMed]
- Molle, C.; Nguyen, M.; Flamand, V.; Renneson, J.; Trottein, F.; De Wit, D.; Willems, F.; Goldman, M.; Goriely, S. IL-27 synthesis induced by TLR ligation critically depends on IFN regulatory factor 3. J. Immunol. 2007, 178, 7607–7615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Guan, X.; Ma, X. Regulation of IL-27 p28 gene expression in macrophages through MyD88- and interferon-gamma-mediated pathways. J. Exp. Med. 2007, 204, 141–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirhonen, J.; Sirén, J.; Julkunen, I.; Matikainen, S. IFN-alpha regulates Toll-like receptor-mediated IL-27 gene expression in human macrophages. J. Leukoc. Biol. 2007, 82, 1185–1192. [Google Scholar] [CrossRef] [PubMed]
- D’Almeida, S.M.; Kauffenstein, G.; Roy, C.; Basset, L.; Papargyris, L.; Henrion, D.; Catros, V.; Ifrah, N.; Descamps, P.; Croue, A.; et al. The ecto-ATPDase CD39 is involved in the acquisition of the immunoregulatory phenotype by M-CSF-macrophages and ovarian cancer tumor-associated macrophages: Regulatory role of IL-27. Oncoimmunology 2016, 5, e1178025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, A.O.; Silver, J.S.; Hunter, C.A. The immunobiology of IL-27. Adv. Immunol. 2012, 115, 1–44. [Google Scholar]
- Villarino, A.V.; Larkin, J.; Saris, C.J.M.; Caton, A.J.; Lucas, S.; Wong, T.; de Sauvage, F.J.; Hunter, C.A. Positive and negative regulation of the IL-27 receptor during lymphoid cell activation. J. Immunol. 2005, 174, 7684–7691. [Google Scholar] [CrossRef]
- Pflanz, S.; Hibbert, L.; Mattson, J.; Rosales, R.; Vaisberg, E.; Bazan, J.F.; Phillips, J.H.; McClanahan, T.K.; de Waal Malefyt, R.; Kastelein, R.A. WSX-1 and glycoprotein 130 constitute a signal-transducing receptor for IL-27. J. Immunol. 2004, 172, 2225–2231. [Google Scholar] [CrossRef]
- Zorzoli, A.; Di Carlo, E.; Cocco, C.; Ognio, E.; Ribatti, D.; Ferretti, E.; Dufour, C.; Locatelli, F.; Montagna, D.; Airoldi, I. Interleukin-27 inhibits the growth of pediatric acute myeloid leukemia in NOD/SCID/Il2rg−/− mice. Clin. Cancer Res. 2012, 18, 1630–1640. [Google Scholar] [CrossRef] [Green Version]
- Canale, S.; Cocco, C.; Frasson, C.; Seganfreddo, E.; Di Carlo, E.; Ognio, E.; Sorrentino, C.; Ribatti, D.; Zorzoli, A.; Basso, G.; et al. Interleukin-27 inhibits pediatric B-acute lymphoblastic leukemia cell spreading in a preclinical model. Leukemia 2011, 25, 1815–1824. [Google Scholar] [CrossRef]
- Cocco, C.; Di Carlo, E.; Zupo, S.; Canale, S.; Zorzoli, A.; Ribatti, D.; Morandi, F.; Ognio, E.; Airoldi, I. Complementary IL-23 and IL-27 anti-tumor activities cause strong inhibition of human follicular and diffuse large B-cell lymphoma growth in vivo. Leukemia 2012, 26, 1365–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Carlo, E.; Sorrentino, C.; Zorzoli, A.; Di Meo, S.; Tupone, M.G.; Ognio, E.; Mincione, G.; Airoldi, I. The antitumor potential of Interleukin-27 in prostate cancer. Oncotarget 2014, 5, 10332–10341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dibra, D.; Cutrera, J.J.; Xia, X.; Birkenbach, M.P.; Li, S. Expression of WSX1 in tumors sensitizes IL-27 signaling-independent natural killer cell surveillance. Cancer Res. 2009, 69, 5505–5513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tait Wojno, E.D.; Hunter, C.A.; Stumhofer, J.S. The Immunobiology of the Interleukin-12 Family: Room for Discovery. Immunity 2019, 50, 851–870. [Google Scholar] [CrossRef] [PubMed]
- Fabbi, M.; Carbotti, G.; Ferrini, S. Dual Roles of IL-27 in Cancer Biology and Immunotherapy. Mediat. Inflamm. 2017, 2017, 3958069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Airoldi, I.; Tupone, M.G.; Esposito, S.; Russo, M.V.; Barbarito, G.; Cipollone, G.; Di Carlo, E. Interleukin-27 re-educates intratumoral myeloid cells and down-regulates stemness genes in non-small cell lung cancer. Oncotarget 2015, 6, 3694–3708. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, H.; Miyazaki, Y. Regulation of immune responses by interleukin-27. Immunol. Rev. 2008, 226, 234–247. [Google Scholar] [CrossRef]
- Kalliolias, G.D.; Ivashkiv, L.B. IL-27 activates human monocytes via STAT1 and suppresses IL-10 production but the inflammatory functions of IL-27 are abrogated by TLRs and p38. J. Immunol. 2008, 180, 6325–6333. [Google Scholar] [CrossRef] [Green Version]
- Guzzo, C.; Ayer, A.; Basta, S.; Banfield, B.W.; Gee, K. IL-27 enhances LPS-induced proinflammatory cytokine production via upregulation of TLR4 expression and signaling in human monocytes. J. Immunol. 2012, 188, 864–873. [Google Scholar] [CrossRef] [Green Version]
- Morandi, F.; Airoldi, I.; Pistoia, V. IL-27 driven upregulation of surface HLA-E expression on monocytes inhibits IFN-gamma release by autologous NK cells. J. Immunol. Res. 2014, 2014, 938561. [Google Scholar] [CrossRef] [Green Version]
- Alspach, E.; Lussier, D.M.; Schreiber, R.D. Interferon γ and Its Important Roles in Promoting and Inhibiting Spontaneous and Therapeutic Cancer Immunity. Cold Spring Harb. Perspect. Biol. 2019, 11, a028480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziblat, A.; Domaica, C.I.; Spallanzani, R.G.; Iraolagoitia, X.L.; Rossi, L.E.; Avila, D.E.; Torres, N.I.; Fuertes, M.B.; Zwirner, N.W. IL-27 stimulates human NK-cell effector functions and primes NK cells for IL-18 responsiveness. Eur. J. Immunol. 2015, 45, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Laroni, A.; Gandhi, R.; Beynon, V.; Weiner, H.L. IL-27 imparts immunoregulatory function to human NK cell subsets. PLoS ONE 2011, 6, e26173. [Google Scholar] [CrossRef] [Green Version]
- Horlad, H.; Ma, C.; Yano, H.; Pan, C.; Ohnishi, K.; Fujiwara, Y.; Endo, S.; Kikukawa, Y.; Okuno, Y.; Matsuoka, M.; et al. An IL-27/Stat3 axis induces expression of programmed cell death 1 ligands (PD-L1/2) on infiltrating macrophages in lymphoma. Cancer Sci. 2016, 107, 1696–1704. [Google Scholar] [CrossRef] [PubMed]
- Carbotti, G.; Barisione, G.; Airoldi, I.; Mezzanzanica, D.; Bagnoli, M.; Ferrero, S.; Petretto, A.; Fabbi, M.; Ferrini, S. IL-27 induces the expression of IDO and PD-L1 in human cancer cells. Oncotarget 2015, 6, 43267–43280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.-Y.; Gleave Parson, M.; Kraft, J.D.; Lyda, L.; Kobe, B.; Davis, C.; Robinson, J.; Peña, M.M.O.; Robinson, C.M. Elevated interleukin-27 levels in human neonatal macrophages regulate indoleamine dioxygenase in a STAT-1 and STAT-3-dependent manner. Immunology 2016, 149, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Villarino, A.V.; Huang, E.; Hunter, C.A. Understanding the pro- and anti-inflammatory properties of IL-27. J. Immunol. 2004, 173, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Kalliolias, G.D.; Gordon, R.A.; Ivashkiv, L.B. Suppression of TNF-α and IL-1 signaling identifies a mechanism of homeostatic regulation of macrophages by IL-27. J. Immunol. 2010, 185, 7047–7056. [Google Scholar] [CrossRef]
- Karakhanova, S.; Bedke, T.; Enk, A.H.; Mahnke, K. IL-27 renders DC immunosuppressive by induction of B7-H1. J. Leukoc. Biol. 2011, 89, 837–845. [Google Scholar] [CrossRef]
- Morandi, F.; Di Carlo, E.; Ferrone, S.; Petretto, A.; Pistoia, V.; Airoldi, I. IL-27 in human secondary lymphoid organs attracts myeloid dendritic cells and impairs HLA class I-restricted antigen presentation. J. Immunol. 2014, 192, 2634–2642. [Google Scholar] [CrossRef] [Green Version]
- Birkholz, J.; Doganci, A.; Darstein, C.; Gehring, S.; Zepp, F.; Meyer, C.U. IL-27 improves migrational and antiviral potential of CB dendritic cells. Hum. Immunol. 2014, 75, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Crabé, S.; Guay-Giroux, A.; Tormo, A.J.; Duluc, D.; Lissilaa, R.; Guilhot, F.; Mavoungou-Bigouagou, U.; Lefouili, F.; Cognet, I.; Ferlin, W.; et al. The IL-27 p28 subunit binds cytokine-like factor 1 to form a cytokine regulating NK and T cell activities requiring IL-6R for signaling. J. Immunol. 2009, 183, 7692–7702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tormo, A.J.; Meliani, Y.; Beaupré, L.A.; Sharma, M.; Fritz, J.H.; Elson, G.; Crabé, S.; Gauchat, J.-F. The composite cytokine p28/cytokine-like factor 1 sustains B cell proliferation and promotes plasma cell differentiation. J. Immunol. 2013, 191, 1657–1665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.-X.; Yu, C.-R.; Mahdi, R.M.; Egwuagu, C.E. Novel IL27p28/IL12p40 cytokine suppressed experimental autoimmune uveitis by inhibiting autoreactive Th1/Th17 cells and promoting expansion of regulatory T cells. J. Biol. Chem. 2012, 287, 36012–36021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gearing, D.P.; Cosman, D. Homology of the p40 subunit of natural killer cell stimulatory factor (NKSF) with the extracellular domain of the interleukin-6 receptor. Cell 1991, 66, 9–10. [Google Scholar] [CrossRef]
- Yoshida, H.; Hunter, C.A. The immunobiology of interleukin-27. Annu. Rev. Immunol. 2015, 33, 417–443. [Google Scholar] [CrossRef]
- Devergne, O.; Hummel, M.; Koeppen, H.; Le Beau, M.M.; Nathanson, E.C.; Kieff, E.; Birkenbach, M. A novel interleukin-12 p40-related protein induced by latent Epstein-Barr virus infection in B lymphocytes. J. Virol. 1996, 70, 1143–1153. [Google Scholar] [CrossRef] [Green Version]
- Niedobitek, G.; Päzolt, D.; Teichmann, M.; Devergne, O. Frequent expression of the Epstein-Barr virus (EBV)-induced gene, EBI3, an IL-12 p40-related cytokine, in Hodgkin and Reed-Sternberg cells. J. Pathol. 2002, 198, 310–316. [Google Scholar] [CrossRef]
- Larousserie, F.; Bardel, E.; Pflanz, S.; Arnulf, B.; Lome-Maldonado, C.; Hermine, O.; Brégeaud, L.; Perennec, M.; Brousse, N.; Kastelein, R.; et al. Analysis of interleukin-27 (EBI3/p28) expression in Epstein-Barr virus- and human T-cell leukemia virus type 1-associated lymphomas: Heterogeneous expression of EBI3 subunit by tumoral cells. Am. J. Pathol. 2005, 166, 1217–1228. [Google Scholar] [CrossRef]
- Nishino, R.; Takano, A.; Oshita, H.; Ishikawa, N.; Akiyama, H.; Ito, H.; Nakayama, H.; Miyagi, Y.; Tsuchiya, E.; Kohno, N.; et al. Identification of Epstein-Barr virus-induced gene 3 as a novel serum and tissue biomarker and a therapeutic target for lung cancer. Clin. Cancer Res. 2011, 17, 6272–6286. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Chen, Q.; Du, W.; Chen, C.; Li, F.; Yang, J.; Peng, J.; Kang, D.; Lin, B.; Chai, X.; et al. Epstein-Barr Virus-Induced Gene 3 (EBI3) Blocking Leads to Induce Antitumor Cytotoxic T Lymphocyte Response and Suppress Tumor Growth in Colorectal Cancer by Bidirectional Reciprocal-Regulation STAT3 Signaling Pathway. Mediat. Inflamm. 2016, 2016, 3214105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egwuagu, C.E.; Yu, C.-R.; Sun, L.; Wang, R. Interleukin 35: Critical regulator of immunity and lymphocyte-mediated diseases. Cytokine Growth Factor Rev. 2015, 26, 587–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pylayeva-Gupta, Y. Molecular Pathways: Interleukin-35 in Autoimmunity and Cancer. Clin. Cancer Res. 2016, 22, 4973–4978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.-C.; Lin, J.-C.; Hwang, W.-L.; Kuo, Y.-J.; Chen, H.-K.; Tai, S.-K.; Lin, C.-C.; Yang, M.-H. Macrophage-secreted interleukin-35 regulates cancer cell plasticity to facilitate metastatic colonization. Nat. Commun. 2018, 9, 3763. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Liu, J.-Q.; Liu, Z.; Shen, R.; Zhang, G.; Xu, J.; Basu, S.; Feng, Y.; Bai, X.-F. Tumor-derived IL-35 promotes tumor growth by enhancing myeloid cell accumulation and angiogenesis. J. Immunol. 2013, 190, 2415–2423. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wei, Y.; Xiao, H.; Liu, X.; Zhang, Y.; Han, G.; Chen, G.; Hou, C.; Ma, N.; Shen, B.; et al. A novel IL-23p19/Ebi3 (IL-39) cytokine mediates inflammation in Lupus-like mice. Eur. J. Immunol. 2016, 46, 1343–1350. [Google Scholar] [CrossRef] [Green Version]
- Bridgewood, C.; Alase, A.; Watad, A.; Wittmann, M.; Cuthbert, R.; McGonagle, D. The IL-23p19/EBI3 heterodimeric cytokine termed IL-39 remains a theoretical cytokine in man. Inflamm. Res. 2019, 68, 423–426. [Google Scholar] [CrossRef] [Green Version]
- Manning, A.A.; Zhao, L.; Zhu, Z.; Xiao, H.; Redington, C.G.; Ding, V.A.; Stewart-Hester, T.; Bai, Q.; Dunlap, J.; Wakefield, M.R.; et al. IL-39 acts as a friend to pancreatic cancer. Med. Oncol. 2018, 36, 12. [Google Scholar] [CrossRef]
- Garbers, C.; Spudy, B.; Aparicio-Siegmund, S.; Waetzig, G.H.; Sommer, J.; Hölscher, C.; Rose-John, S.; Grötzinger, J.; Lorenzen, I.; Scheller, J. An interleukin-6 receptor-dependent molecular switch mediates signal transduction of the IL-27 cytokine subunit p28 (IL-30) via a gp130 protein receptor homodimer. J. Biol. Chem. 2013, 288, 4346–4354. [Google Scholar] [CrossRef] [Green Version]
- Shimozato, O.; Sato, A.; Kawamura, K.; Chiyo, M.; Ma, G.; Li, Q.; Tagawa, M. The secreted form of p28 subunit of interleukin (IL)-27 inhibits biological functions of IL-27 and suppresses anti-allogeneic immune responses. Immunology 2009, 128, e816–e825. [Google Scholar] [CrossRef]
- Stumhofer, J.S.; Tait, E.D.; Quinn, W.J.; Hosken, N.; Spudy, B.; Goenka, R.; Fielding, C.A.; O’Hara, A.C.; Chen, Y.; Jones, M.L.; et al. A role for IL-27p28 as an antagonist of gp130-mediated signaling. Nat. Immunol. 2010, 11, 1119–1126. [Google Scholar] [CrossRef] [PubMed]
- Müller, S.I.; Friedl, A.; Aschenbrenner, I.; Esser-von Bieren, J.; Zacharias, M.; Devergne, O.; Feige, M.J. A folding switch regulates interleukin 27 biogenesis and secretion of its α-subunit as a cytokine. Proc. Natl. Acad. Sci. USA 2019, 116, 1585–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.-F.; Zhao, A.-D. Common Polymorphisms in IL-27 Genes May Contribute to Risk of Various Human Diseases in Asian Populations: A Meta-Analysis. Med. Sci. Monit. 2016, 22, 766–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, T.; Zhao, H.; Wang, K.; Sun, W.; Li, L.; Wang, W. Association of IL-27 polymorphisms and cancer risk in Chinese population. J. Recept. Signal Transduct. Res. 2015, 35, 180–183. [Google Scholar] [CrossRef] [PubMed]
- Skiniotis, G.; Lupardus, P.J.; Martick, M.; Walz, T.; Garcia, K.C. Structural organization of a full-length gp130/LIF-R cytokine receptor transmembrane complex. Mol. Cell 2008, 31, 737–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bravo, J.; Heath, J.K. Receptor recognition by gp130 cytokines. EMBO J. 2000, 19, 2399–2411. [Google Scholar] [CrossRef] [Green Version]
- Petes, C.; Mariani, M.K.; Yang, Y.; Grandvaux, N.; Gee, K. Interleukin (IL)-6 Inhibits IL-27- and IL-30-Mediated Inflammatory Responses in Human Monocytes. Front. Immunol. 2018, 9, 256. [Google Scholar] [CrossRef] [Green Version]
- Grivennikov, S.; Karin, M. Autocrine IL-6 signaling: A key event in tumorigenesis? Cancer Cell 2008, 13, 7–9. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T.; Kishimoto, T. The biology and medical implications of interleukin-6. Cancer Immunol. Res. 2014, 2, 288–294. [Google Scholar] [CrossRef] [Green Version]
- Di Carlo, E. Interleukin-30: A novel microenvironmental hallmark of prostate cancer progression. Oncoimmunology 2014, 3, e27618. [Google Scholar] [CrossRef] [Green Version]
- Sorrentino, C.; Ciummo, S.L.; Cipollone, G.; Caputo, S.; Bellone, M.; Di Carlo, E. Interleukin-30/IL27p28 Shapes Prostate Cancer Stem-like Cell Behavior and Is Critical for Tumor Onset and Metastasization. Cancer Res. 2018, 78, 2654–2668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priceman, S.J.; Sung, J.L.; Shaposhnik, Z.; Burton, J.B.; Torres-Collado, A.X.; Moughon, D.L.; Johnson, M.; Lusis, A.J.; Cohen, D.A.; Iruela-Arispe, M.L.; et al. Targeting distinct tumor-infiltrating myeloid cells by inhibiting CSF-1 receptor: Combating tumor evasion of antiangiogenic therapy. Blood 2010, 115, 1461–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Han, X.; Sun, Y.; Shang, C.; Wei, M.; Ba, X.; Zeng, X. Chemokine (C-X-C motif) ligand 1 and CXCL2 produced by tumor promote the generation of monocytic myeloid-derived suppressor cells. Cancer Sci. 2018, 109, 3826–3839. [Google Scholar] [CrossRef] [PubMed]
- Najjar, Y.G.; Rayman, P.; Jia, X.; Pavicic, P.G.; Rini, B.I.; Tannenbaum, C.; Ko, J.; Haywood, S.; Cohen, P.; Hamilton, T.; et al. Myeloid-Derived Suppressor Cell Subset Accumulation in Renal Cell Carcinoma Parenchyma Is Associated with Intratumoral Expression of IL1β, IL8, CXCL5, and Mip-1α. Clin. Cancer Res. 2017, 23, 2346–2355. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.; Chen, J.; Zhang, W.; Zhang, R.; Ye, Y.; Liu, P.; Yu, W.; Wei, F.; Ren, X.; Yu, J. Interleukin-6 Trans-Signaling Pathway Promotes Immunosuppressive Myeloid-Derived Suppressor Cells via Suppression of Suppressor of Cytokine Signaling 3 in Breast Cancer. Front. Immunol. 2017, 8, 1840. [Google Scholar] [CrossRef] [Green Version]
- Pal, S.K.; Kortylewski, M. Breaking bad habits: Targeting MDSCs to alleviate immunosuppression in prostate cancer. Oncoimmunology 2016, 5, e1078060. [Google Scholar] [CrossRef] [Green Version]
- Veglia, F.; Perego, M.; Gabrilovich, D. Myeloid-derived suppressor cells coming of age. Nat. Immunol. 2018, 19, 108–119. [Google Scholar] [CrossRef]
- Greenberg, N.M.; DeMayo, F.; Finegold, M.J.; Medina, D.; Tilley, W.D.; Aspinall, J.O.; Cunha, G.R.; Donjacour, A.A.; Matusik, R.J.; Rosen, J.M. Prostate cancer in a transgenic mouse. Proc. Natl. Acad. Sci. USA 1995, 92, 3439–3443. [Google Scholar] [CrossRef] [Green Version]
- Kaplan-Lefko, P.J.; Chen, T.-M.; Ittmann, M.M.; Barrios, R.J.; Ayala, G.E.; Huss, W.J.; Maddison, L.A.; Foster, B.A.; Greenberg, N.M. Pathobiology of autochthonous prostate cancer in a pre-clinical transgenic mouse model. Prostate 2003, 55, 219–237. [Google Scholar] [CrossRef]
- Jachetti, E.; Mazzoleni, S.; Grioni, M.; Ricupito, A.; Brambillasca, C.; Generoso, L.; Calcinotto, A.; Freschi, M.; Mondino, A.; Galli, R.; et al. Prostate cancer stem cells are targets of both innate and adaptive immunity and elicit tumor-specific immune responses. Oncoimmunology 2013, 2, e24520. [Google Scholar] [CrossRef] [Green Version]
- Sorrentino, C.; Yin, Z.; Ciummo, S.; Lanuti, P.; Lu, L.-F.; Marchisio, M.; Bellone, M.; Di Carlo, E. Targeting Interleukin(IL)-30/IL-27p28 signaling in cancer stem-like cells and host environment synergistically inhibits prostate cancer growth and improves survival. J. Immunother. Cancer 2019, 7, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzoleni, S.; Jachetti, E.; Morosini, S.; Grioni, M.; Piras, I.S.; Pala, M.; Bulfone, A.; Freschi, M.; Bellone, M.; Galli, R. Gene signatures distinguish stage-specific prostate cancer stem cells isolated from transgenic adenocarcinoma of the mouse prostate lesions and predict the malignancy of human tumors. Stem Cells Transl. Med. 2013, 2, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Wang, X.; Choi, S.Y.C.; Ci, X.; Dong, X.; Wang, Y. Immune phenotypes of prostate cancer cells: Evidence of epithelial immune cell-like transition? Asian J. Urol. 2016, 3, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Liang, R.; Luo, W.; Liu, C.; Wu, X.; Gao, Y.; Hao, J.; Cao, G.; Chen, X.; Wei, J.; et al. High susceptibility to liver injury in IL-27 p28 conditional knockout mice involves intrinsic interferon-γ dysregulation of CD4+ T cells. Hepatology 2013, 57, 1620–1631. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wang, X.; Wang, L.; Ma, X.; Gong, Z.; Zhang, S.; Li, Y. Targeting the IDO1 pathway in cancer: From bench to bedside. J. Hematol. Oncol. 2018, 11, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smyth, M.J.; Ngiow, S.F.; Ribas, A.; Teng, M.W.L. Combination cancer immunotherapies tailored to the tumour microenvironment. Nat. Rev. Clin. Oncol. 2016, 13, 143–158. [Google Scholar] [CrossRef]
- Wei, S.C.; Duffy, C.R.; Allison, J.P. Fundamental Mechanisms of Immune Checkpoint Blockade Therapy. Cancer Discov. 2018, 8, 1069–1086. [Google Scholar] [CrossRef] [Green Version]
- Marra, A.; Viale, G.; Curigliano, G. Recent advances in triple negative breast cancer: The immunotherapy era. BMC Med. 2019, 17, 90. [Google Scholar] [CrossRef] [Green Version]
- Sunshine, J.; Taube, J.M. PD-1/PD-L1 inhibitors. Curr. Opin. Pharmacol. 2015, 23, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Dallos, M.C.; Drake, C.G. Blocking PD-1/PD-L1 in Genitourinary Malignancies: To Immunity and Beyond. Cancer J. 2018, 24, 20–30. [Google Scholar] [CrossRef]
- Amoozgar, Z.; Goldberg, M.S. Targeting myeloid cells using nanoparticles to improve cancer immunotherapy. Adv. Drug Deliv. Rev. 2015, 91, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Kantoff, P.W.; Wooster, R.; Farokhzad, O.C. Cancer nanomedicine: Progress, challenges and opportunities. Nat. Rev. Cancer 2017, 17, 20–37. [Google Scholar] [CrossRef] [PubMed]
- Ediriwickrema, A.; Saltzman, W.M. Nanotherapy for Cancer: Targeting and Multifunctionality in the Future of Cancer Therapies. ACS Biomater. Sci. Eng. 2015, 1, 64–78. [Google Scholar] [CrossRef] [PubMed]



© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Carlo, E. Decoding the Role of Interleukin-30 in the Crosstalk between Cancer and Myeloid Cells. Cells 2020, 9, 615. https://doi.org/10.3390/cells9030615
Di Carlo E. Decoding the Role of Interleukin-30 in the Crosstalk between Cancer and Myeloid Cells. Cells. 2020; 9(3):615. https://doi.org/10.3390/cells9030615
Chicago/Turabian StyleDi Carlo, Emma. 2020. "Decoding the Role of Interleukin-30 in the Crosstalk between Cancer and Myeloid Cells" Cells 9, no. 3: 615. https://doi.org/10.3390/cells9030615
APA StyleDi Carlo, E. (2020). Decoding the Role of Interleukin-30 in the Crosstalk between Cancer and Myeloid Cells. Cells, 9(3), 615. https://doi.org/10.3390/cells9030615