Local Mast Cell Activation Promotes Neovascularization

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Collection from Patients with End-Stage Peripheral Artery Disease
2.2. Hind Limb Ischemia Model
2.3. Local Mast Cell Activation with DPN treatment
2.4. Laser Doppler Perfusion Measurements
2.5. Immunohistochemistry
2.6. FACS Analysis
2.7. Statistical Analysis
3. Results
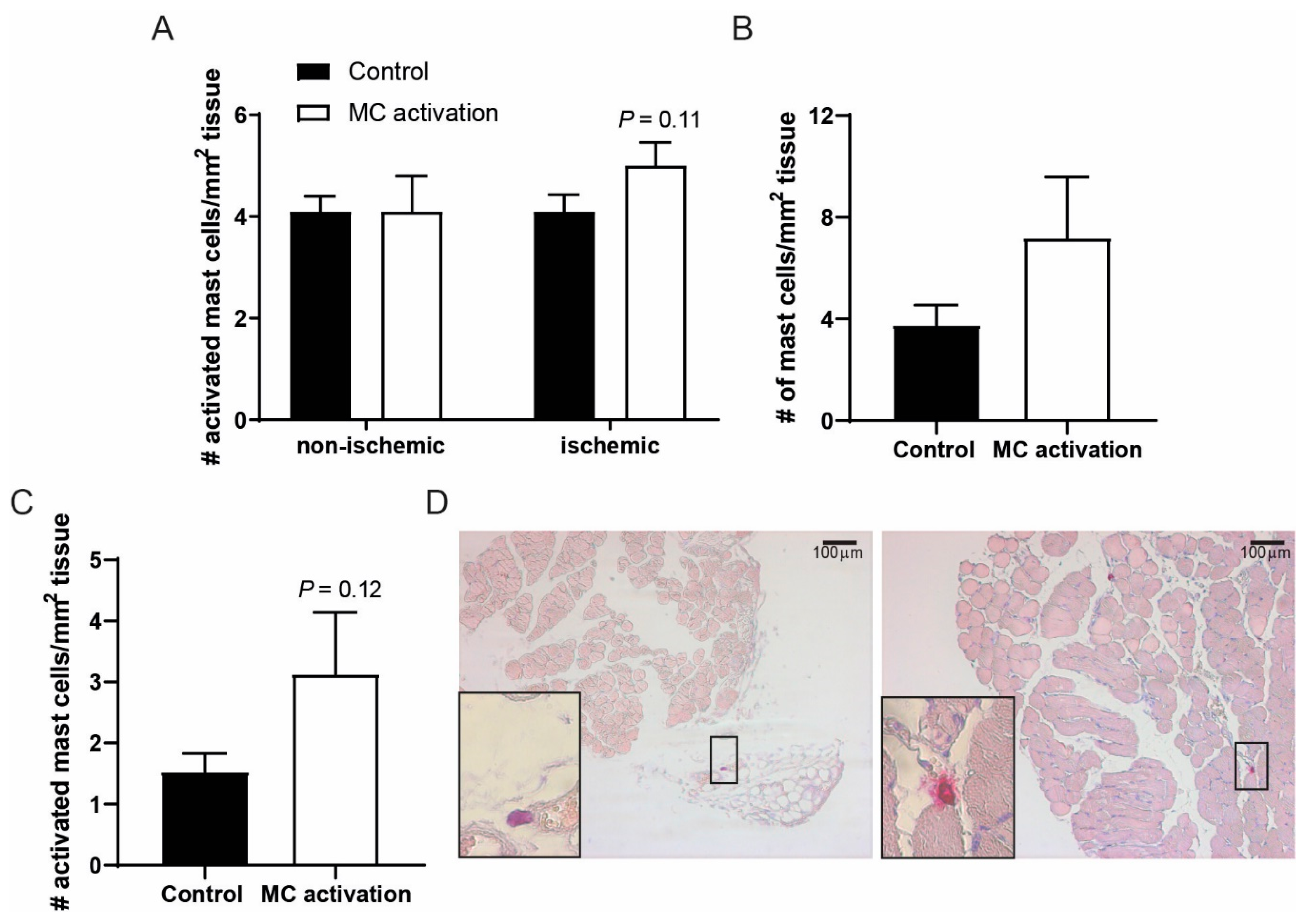
3.1. Mast Cells Localization in Calf Muscles of Peripheral Artery Disease Patients
3.2. Activation of Mast Cells upon Ischemia in a Murine Hindlimb Ischemia Model
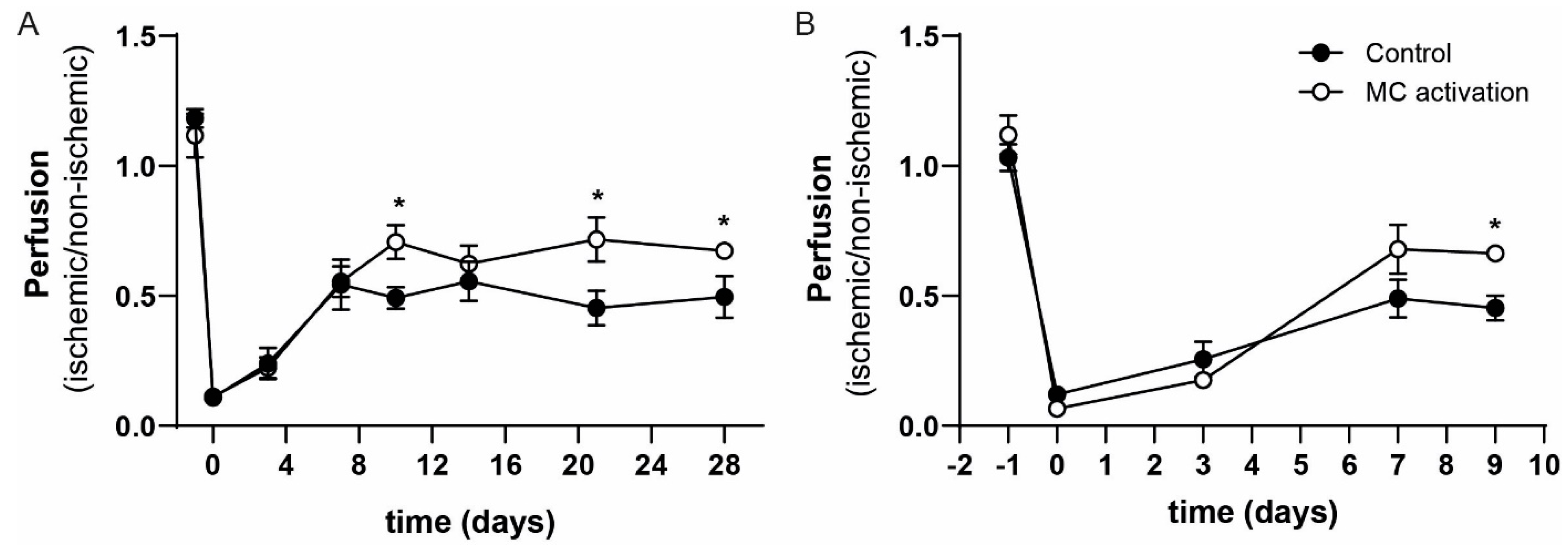
3.3. Effect of Local Mast Cell Activation on Post Ischemic Blood Flow Recovery.
3.4. Local Mast Cell Activation Induced by DNP
3.5. Contribution of Arteriogenesis to the Increase in Blood Flow Recovery by Activated Mast Cells
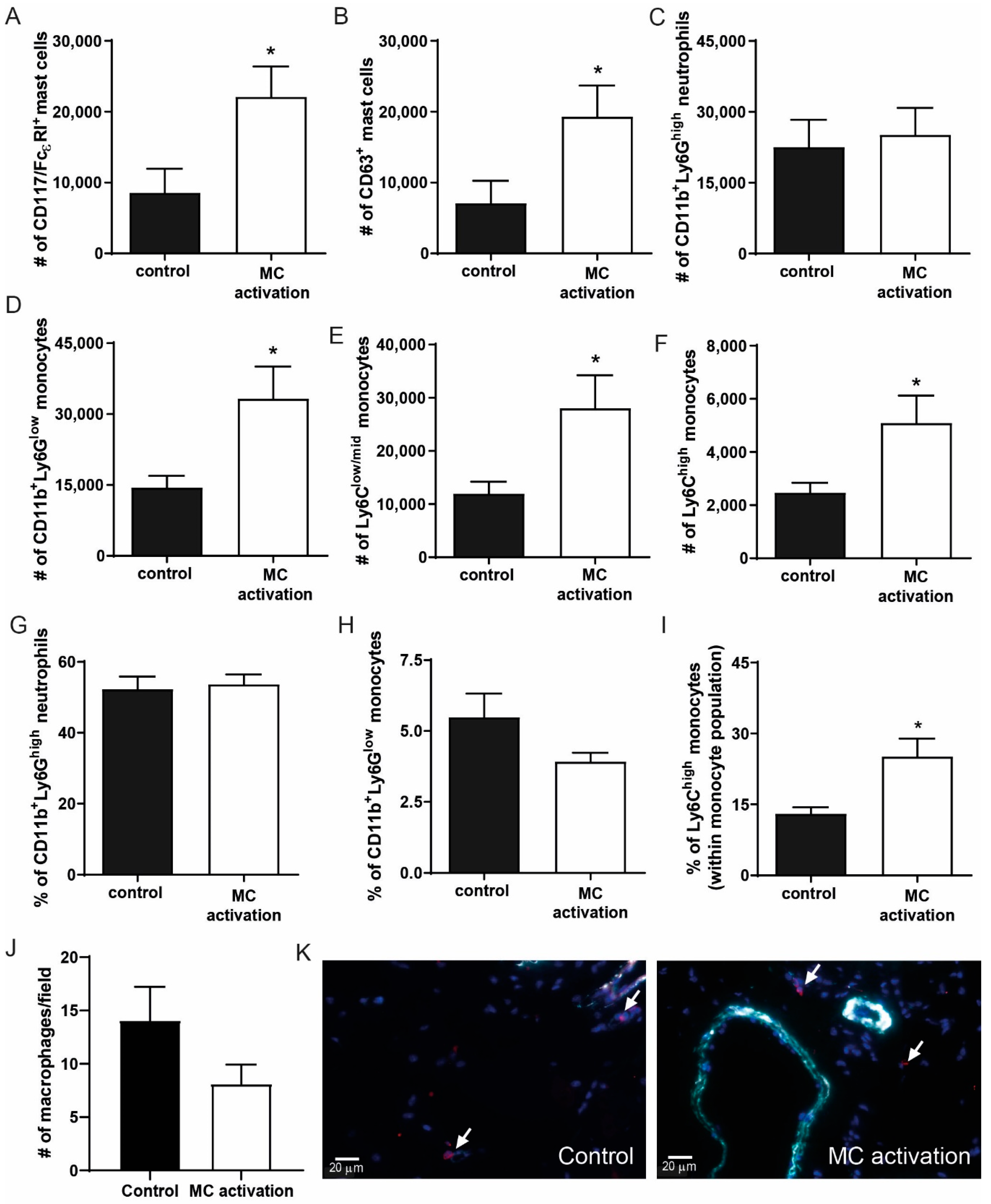
3.6. Effects of Mast Cell Activation on Angiogenesis
3.7. Local Mast Cell Activation Induces a Pro-Inflammatory Monocyte Response
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Kaartinen, M.; Penttila, A.; Kovanen, P.T. Accumulation of activated mast cells in the shoulder region of human coronary atheroma, the predilection site of atheromatous rupture. Circulation 1994, 90, 1669–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovanen, P.T.; Kaartinen, M.; Paavonen, T. Infiltrates of activated mast cells at the site of coronary atheromatous erosion or rupture in myocardial infarction. Circulation 1995, 92, 1084–1088. [Google Scholar] [CrossRef] [PubMed]
- Bot, I.; Shi, G.P.; Kovanen, P.T. Mast cells as effectors in atherosclerosis. Arter. Thromb. Vasc. Biol. 2015, 35, 265–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, G.P.; Bot, I.; Kovanen, P.T. Mast cells in human and experimental cardiometabolic diseases. Nat. Rev. Cardiol. 2015, 12, 643–658. [Google Scholar] [CrossRef]
- Willems, S.; Vink, A.; Bot, I.; Quax, P.H.; de Borst, G.J.; de Vries, J.P.; van de Weg, S.M.; Moll, F.L.; Kuiper, J.; Kovanen, P.T.; et al. Mast cells in human carotid atherosclerotic plaques are associated with intraplaque microvessel density and the occurrence of future cardiovascular events. Eur. Heart J. 2013, 34, 3699–3706. [Google Scholar] [CrossRef] [Green Version]
- Chillo, O.; Kleinert, E.C.; Lautz, T.; Lasch, M.; Pagel, J.I.; Heun, Y.; Troidl, K.; Fischer, S.; Caballero-Martinez, A.; Mauer, A.; et al. Perivascular Mast Cells Govern Shear Stress-Induced Arteriogenesis by Orchestrating Leukocyte Function. Cell Rep. 2016, 16, 2197–2207. [Google Scholar] [CrossRef] [Green Version]
- Bot, I.; de Jager, S.C.; Zernecke, A.; Lindstedt, K.A.; van Berkel, T.J.; Weber, C.; Biessen, E.A. Perivascular mast cells promote atherogenesis and induce plaque destabilization in apolipoprotein E-deficient mice. Circulation 2007, 115, 2516–2525. [Google Scholar] [CrossRef]
- Sun, J.; Sukhova, G.K.; Wolters, P.J.; Yang, M.; Kitamoto, S.; Libby, P.; MacFarlane, L.A.; Mallen-St, C.J.; Shi, G.P. Mast cells promote atherosclerosis by releasing proinflammatory cytokines. Nat. Med. 2007, 13, 719–724. [Google Scholar] [CrossRef]
- Indhirajanti, S.; van Daele, P.L.A.; Bos, S.; Mulder, M.T.; Bot, I.; Roeters van Lennep, J.E. Systemic mastocytosis associates with cardiovascular events despite lower plasma lipid levels. Atherosclerosis 2018, 268, 152–156. [Google Scholar] [CrossRef] [Green Version]
- Albini, A.; Bruno, A.; Noonan, D.M.; Mortara, L. Contribution to Tumor Angiogenesis From Innate Immune Cells Within the Tumor Microenvironment: Implications for Immunotherapy. Front. Immunol. 2018, 9, 527. [Google Scholar] [CrossRef]
- van Weel, V.; Seghers, L.; de Vries, M.R.; Kuiper, E.J.; Schlingemann, R.O.; Bajema, I.M.; Lindeman, J.H.; Delis-van Diemen, P.M.; van Hinsbergh, V.W.; van Bockel, J.H.; et al. Expression of vascular endothelial growth factor, stromal cell-derived factor-1, and CXCR4 in human limb muscle with acute and chronic ischemia. Arter. Thromb. Vasc. Biol. 2007, 27, 1426–1432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campia, U.; Gerhard-Herman, M.; Piazza, G.; Goldhaber, S.Z. Peripheral Artery Disease: Past, Present, and Future. Am. J. Med. 2019, 132, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Epstein, S.E.; Stabile, E.; Kinnaird, T.; Lee, C.W.; Clavijo, L.; Burnett, M.S. Janus phenomenon: The interrelated tradeoffs inherent in therapies designed to enhance collateral formation and those designed to inhibit atherogenesis. Circulation 2004, 109, 2826–2831. [Google Scholar] [CrossRef] [PubMed]
- de Vries, M.R.; Wezel, A.; Schepers, A.; van Santbrink, P.J.; Woodruff, T.M.; Niessen, H.W.; Hamming, J.F.; Kuiper, J.; Bot, I.; Quax, P.H. Complement factor C5a as mast cell activator mediates vascular remodelling in vein graft disease. Cardiovasc. Res. 2013, 97, 311–320. [Google Scholar] [CrossRef] [Green Version]
- de Vries, M.R.; Seghers, L.; van, B.J.; Peters, H.A.; de Jong, R.C.; Hamming, J.F.; Toes, R.E.; van Hinsbergh, V.W.; Quax, P.H. C57BL/6 NK cell gene complex is crucially involved in vascular remodeling. J. Mol. Cell Cardiol. 2013, 64, 51–58. [Google Scholar] [CrossRef]
- Simons, K.H.; Aref, Z.; Peters, H.A.B.; Welten, S.P.; Nossent, A.Y.; Jukema, J.W.; Hamming, J.F.; Arens, R.; de Vries, M.R.; Quax, P.H.A. The role of CD27-CD70-mediated T cell co-stimulation in vasculogenesis, arteriogenesis and angiogenesis. Int. J. Cardiol. 2018, 260, 184–190. [Google Scholar] [CrossRef] [Green Version]
- Kraneveld, A.D.; Buckley, T.L.; van Heuven-Nolsen, D.; van, S.Y.; Koster, A.S.; Nijkamp, F.P. Delayed-type hypersensitivity-induced increase in vascular permeability in the mouse small intestine: Inhibition by depletion of sensory neuropeptides and NK1 receptor blockade. Br. J. Pharm. 1995, 114, 1483–1489. [Google Scholar] [CrossRef] [Green Version]
- den Dekker, W.K.; Tempel, D.; Bot, I.; Biessen, E.A.; Joosten, L.A.; Netea, M.G.; van der Meer, J.W.; Cheng, C.; Duckers, H.J. Mast cells induce vascular smooth muscle cell apoptosis via a toll-like receptor 4 activation pathway. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1960–1969. [Google Scholar] [CrossRef] [Green Version]
- Aref, Z.; de Vries, M.R.; Quax, P.H.A. Variations in Surgical Procedures for Inducing Hind Limb Ischemia in Mice and the Impact of These Variations on Neovascularization Assessment. Int J. Mol. Sci 2019, 20, 3704. [Google Scholar] [CrossRef] [Green Version]
- Kritikou, E.; van der Heijden, T.; Swart, M.; van Duijn, J.; Slutter, B.; Wezel, A.; Smeets, H.J.; Maffia, P.; Kuiper, J.; Bot, I. Hypercholesterolemia Induces a Mast Cell-CD4(+) T Cell Interaction in Atherosclerosis. J. Immunol. 2019, 202, 1531–1539. [Google Scholar] [CrossRef]
- Wezel, A.; Lagraauw, H.M.; van der Velden, D.; de Jager, S.C.; Quax, P.H.; Kuiper, J.; Bot, I. Mast cells mediate neutrophil recruitment during atherosclerotic plaque progression. Atherosclerosis 2015, 241, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Nowak-Sliwinska, P.; Alitalo, K.; Allen, E.; Anisimov, A.; Aplin, A.C.; Auerbach, R.; Augustin, H.G.; Bates, D.O.; van Beijnum, J.R.; Bender, R.H.F.; et al. Consensus guidelines for the use and interpretation of angiogenesis assays. Angiogenesis 2018, 21, 425–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ollikainen, E.; Tulamo, R.; Frosen, J.; Lehti, S.; Honkanen, P.; Hernesniemi, J.; Niemela, M.; Kovanen, P.T. Mast cells, neovascularization, and microhemorrhages are associated with saccular intracranial artery aneurysm wall remodeling. J. Neuropathol Exp. Neurol. 2014, 73, 855–864. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Ma, C.; Yu, T.; Song, J.; Leng, J.; Gu, X.; Li, J. Activation mechanisms and multifaceted effects of mast cells in ischemia reperfusion injury. Exp. Cell Res. 2019, 376, 227–235. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bot, I.; van der Velden, D.; Bouwman, M.; Kröner, M.J.; Kuiper, J.; Quax, P.H.A.; de Vries, M.R. Local Mast Cell Activation Promotes Neovascularization. Cells 2020, 9, 701. https://doi.org/10.3390/cells9030701
Bot I, van der Velden D, Bouwman M, Kröner MJ, Kuiper J, Quax PHA, de Vries MR. Local Mast Cell Activation Promotes Neovascularization. Cells. 2020; 9(3):701. https://doi.org/10.3390/cells9030701
Chicago/Turabian StyleBot, Ilze, Daniël van der Velden, Merel Bouwman, Mara J. Kröner, Johan Kuiper, Paul H. A. Quax, and Margreet R. de Vries. 2020. "Local Mast Cell Activation Promotes Neovascularization" Cells 9, no. 3: 701. https://doi.org/10.3390/cells9030701
APA StyleBot, I., van der Velden, D., Bouwman, M., Kröner, M. J., Kuiper, J., Quax, P. H. A., & de Vries, M. R. (2020). Local Mast Cell Activation Promotes Neovascularization. Cells, 9(3), 701. https://doi.org/10.3390/cells9030701