Satellite Cells in Skeletal Muscle of the Hibernating Dormouse, a Natural Model of Quiescence and Re-Activation: Focus on the Cell Nucleus





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
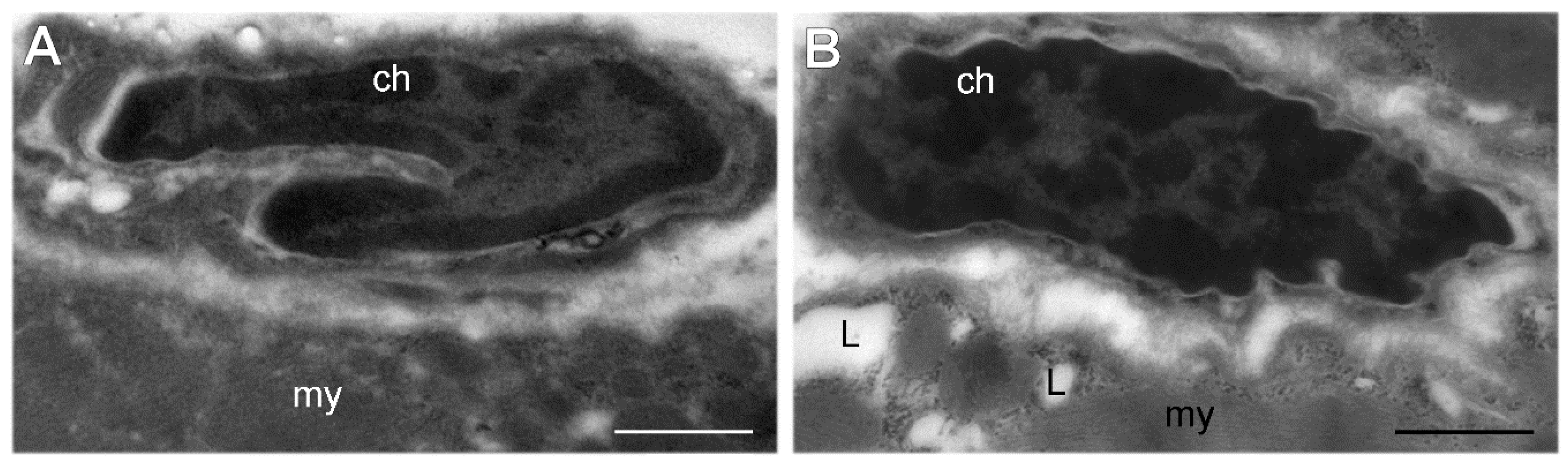
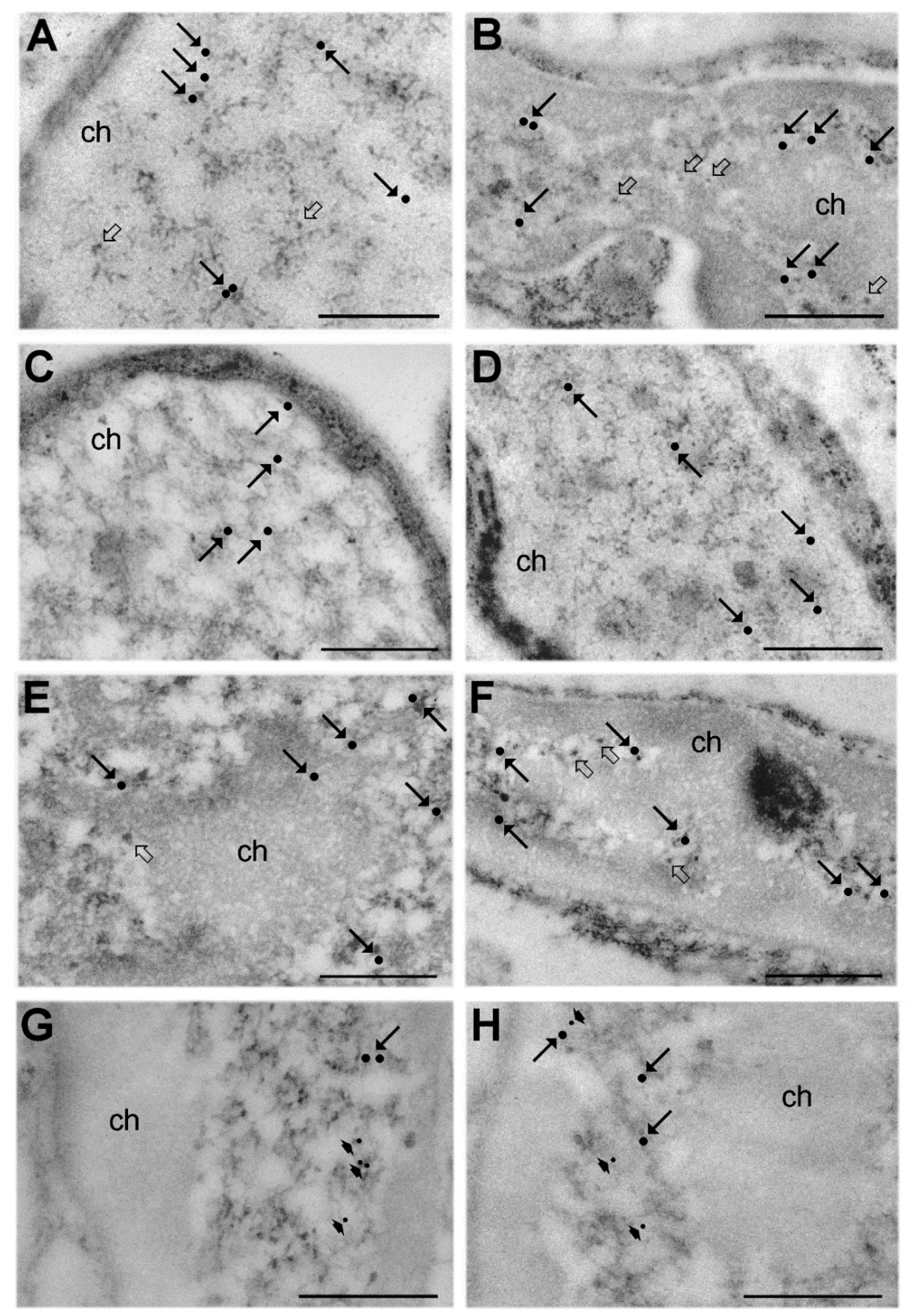
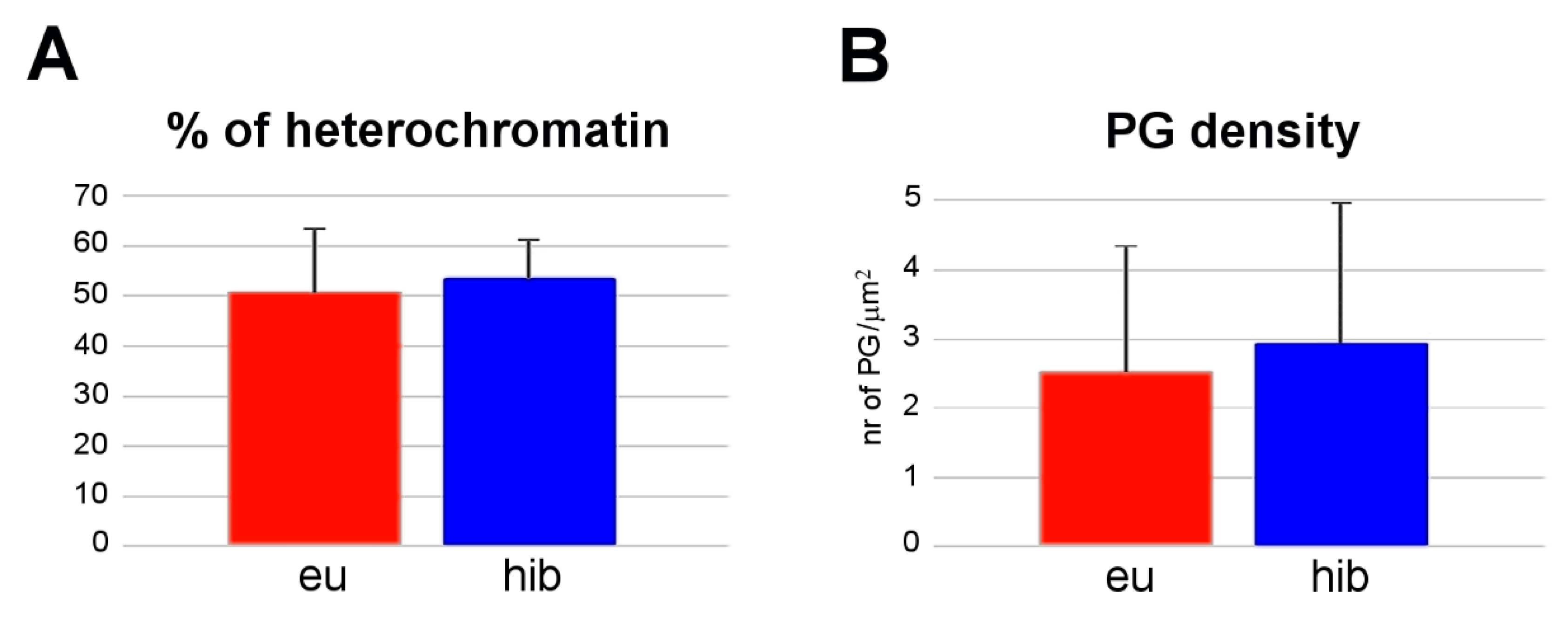
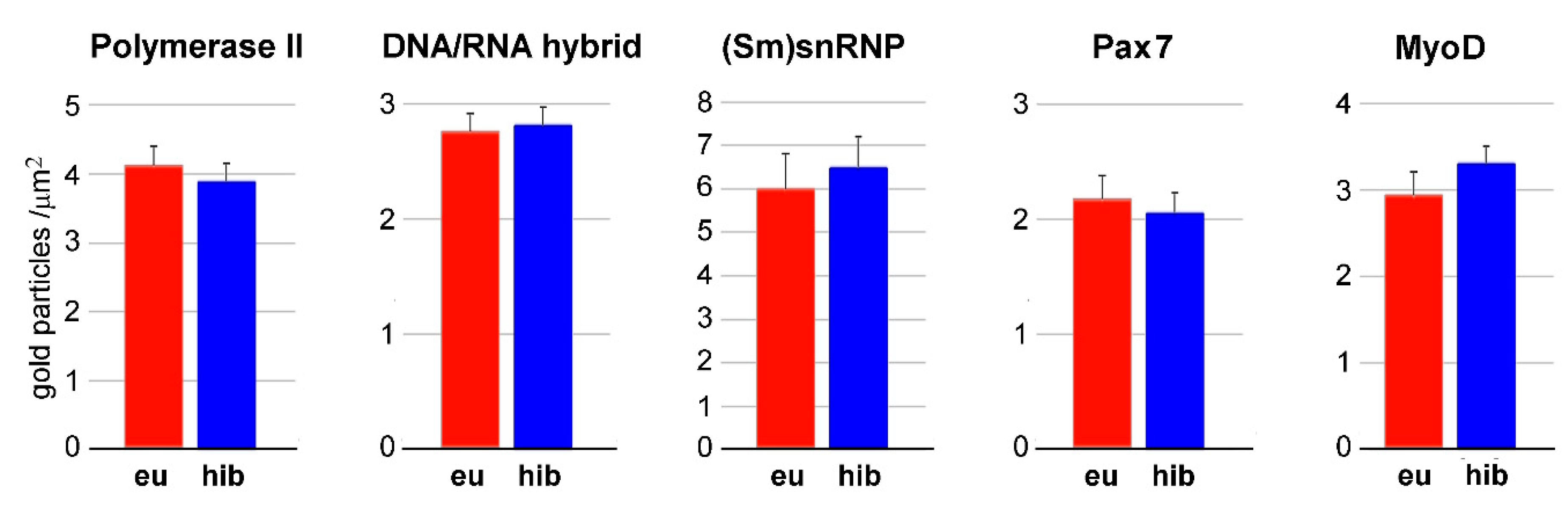
3. Results
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Dumont, N.A.; Bentzinger, C.F.; Sincennes, M.C.; Rudnicki, M.A. Satellite cells and skeletal muscle regeneration. Compr. Physiol. 2015, 5, 1027–1059. [Google Scholar] [PubMed]
- Mauro, A. Satellite cell of skeletal muscle fibers. J. Biophys. Biochem. Cytol. 1961, 9, 493–495. [Google Scholar] [CrossRef] [PubMed]
- Snijders, T.; Nederveen, J.P.; McKay, B.R.; Joanisse, S.; Verdijk, L.B.; van Loon, L.J.; Parise, G. Satellite cells in human skeletal muscle plasticity. Front. Physiol. 2015, 6, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forcina, L.; Miano, C.; Pelosi, L.; Musarò, A. An Overview about the biology of skeletal muscle satellite cells. Cur. Genomics. 2019, 20, 24–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashinchian, O.; Pisconti, A.; Le Moal, E.; Bentzinger, C.F. The muscle stem cell niche in health and disease. Curr. Top. Dev. Biol. 2018, 126, 23–65. [Google Scholar] [PubMed]
- Yin, H.; Price, F.; Rudnicki, M.A. Satellite cells and the muscle stem cell niche. Physiol. Rev. 2013, 93, 23–67. [Google Scholar] [CrossRef] [Green Version]
- Muir, A.R.; Kanji, A.H.; Allbrook, D. The structure of the satellite cells in skeletal muscle. J. Anat. 1965, 99, 435–444. [Google Scholar]
- Schultz, E.; Gibson, M.C.; Champion, T. Satellite cells are mitotically quiescent in mature mouse muscle: An EM and radioautographic study. J. Exp. Zool. 1978, 206, 451–456. [Google Scholar] [CrossRef]
- Shi, X.; Garry, D.J. Muscle stem cells in development, regeneration, and disease. Genes Dev. 2006, 20, 1692–1708. [Google Scholar] [CrossRef]
- Geiser, F. Hibernation. Curr. Biol. 2013, 23, R188–R193. [Google Scholar] [CrossRef] [Green Version]
- Cotton, C.J.; Harlow, H.J. Avoidance of skeletal muscle atrophy in spontaneous and facultative hibernators. Physiol. Biochem. Zool. 2010, 83, 551–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andres-Mateos, E.; Brinkmeier, H.; Burks, T.N.; Mejias, R.; Files, D.C.; Steinberger, M.; Soleimani, A.; Marx, R.; Simmers, J.L.; Lin, B.; et al. Activation of serum/glucocorticoid-induced kinase 1 (SGK1) is important to maintain skeletal muscle homeostasis and prevent atrophy. EMBO Mol. Med. 2013, 5, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Ivakine, E.A.; Cohn, R.D. Maintaining skeletal muscle mass: Lessons learned from hibernation. Exp. Physiol. 2014, 99, 632–637. [Google Scholar] [CrossRef] [PubMed]
- Tessier, S.N.; Storey, K.B. Lessons from mammalian hibernators: Molecular insights into striated muscle plasticity and remodeling. Biomol. Concepts. 2016, 7, 69–92. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.; Peng, X.; Yan, X.; Zhang, J.; Xu, S.; Wang, H.; Wang, Z.; Ma, X.; Gao, Y. Autophagy and Akt-mTOR signaling display periodic oscillations during torpor-arousal cycles in oxidative skeletal muscle of Daurian ground squirrels (Spermophilus dauricus). J. Comp. Physiol. B 2020, 190, 113–123. [Google Scholar] [CrossRef]
- Brooks, N.E.; Myburgh, K.H.; Storey, K.B. Muscle satellite cells increase during hibernation in ground squirrels. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2015, 189, 55–61. [Google Scholar] [CrossRef]
- Andres-Mateos, E.; Mejias, R.; Soleimani, A.; Lin, B.M.; Burks, T.N.; Marx, R.; Lin, B.; Zellars, R.C.; Zhang, Y.; Huso, D.L.; et al. Impaired skeletal muscle regeneration in the absence of fibrosis during hibernation in 13-lined ground squirrels. PLoS ONE 2012, 7, e48884. [Google Scholar] [CrossRef] [Green Version]
- Malatesta, M.; Zancanaro, C.; Biggiogera, M. Immunoelectron microscopic characterization of nucleolus-associated domains during hibernation. Microsc. Res. Tech. 2011, 74, 47–53. [Google Scholar] [CrossRef]
- Suozzi, A.; Malatesta, M.; Zancanaro, C. Subcellular distribution of key enzymes of lipid metabolism during the euthermia-hibernation-arousal cycle. J. Anat. 2009, 214, 956–962. [Google Scholar] [CrossRef]
- Malatesta, M.; Biggiogera, M.; Baldelli, B.; Barabino, S.M.; Martin, T.E.; Zancanaro, C. Hibernation as a far-reaching program for the modulation of RNA transcription. Microsc. Res. Tech. 2008, 71, 564–572. [Google Scholar] [CrossRef]
- Malatesta, M.; Luchetti, F.; Marcheggiani, F.; Fakan, S.; Gazzanelli, G. Disassembly of nuclear bodies during arousal from hibernation: An in vitro study. Chromosoma 2001, 110, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Malatesta, M.; Gazzanelli, G.; Battistelli, S.; Martin, T.E.; Amalric, F.; Fakan, S. Nucleoli undergo structural and molecular modifications during hibernation. Chromosoma 2000, 109, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Malatesta, M.; Cardinali, A.; Battistelli, S.; Zancanaro, C.; Martin, T.E.; Fakan, S.; Gazzanelli, G. Nuclear bodies are usual constituents in tissues of hibernating dormice. Anat. Rec. 1999, 254, 389–395. [Google Scholar] [CrossRef]
- Tamburini, M.; Malatesta, M.; Zancanaro, C.; Martin, T.E.; Fu, X.D.; Vogel, P.; Fakan, S. Dense granular bodies: A novel nucleoplasmic structure in hibernating dormice. Histochem. Cell. Biol. 1996, 106, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Malatesta, M.; Zancanaro, C.; Tamburini, M.; Martin, T.E.; Fu, X.D.; Vogel, P.; Fakan, S. Novel nuclear ribonucleoprotein structural components in the dormouse adrenal cortex during hibernation. Chromosoma 1995, 104, 121–128. [Google Scholar] [CrossRef]
- Malatesta, M.; Zancanaro, C.; Martin, T.E.; Chan, E.K.; Amalric, F.; Lührmann, R.; Vogel, P.; Fakan, S. Cytochemical and immunocytochemical characterization of nuclear bodies during hibernation. Eur. J. Cell. Biol. 1994, 65, 82–93. [Google Scholar]
- Malatesta, M.; Zancanaro, C.; Martin, T.E.; Chan, E.K.; Amalric, F.; Lührmann, R.; Vogel, P.; Fakan, S. Is the coiled body involved in nucleolar functions? Exp. Cell. Res. 1994, 211, 415–419. [Google Scholar] [CrossRef]
- Malatesta, M.; Perdoni, F.; Battistelli, S.; Muller, S.; Zancanaro, C. The cell nuclei of skeletal muscle cells are transcriptionally active in hibernating edible dormice. BMC Cell Biol. 2009, 10, 19. [Google Scholar] [CrossRef]
- Testillano, P.S.; Gorab, E.; Risueno, M.C. A new approach to map transcription sites at the ultrastructural level. J. Histochem. Cytochem. 1994, 42, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Malatesta, M.; Perdoni, F.; Muller, S.; Zancanaro, C.; Pellicciari, C. Nuclei of aged myofibres undergo structural and functional changes suggesting impairment in RNA processing. Eur. J. Histochem. 2009, 53, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Lührmann, R.; Kastner, B.; Bach, M. Structure of spliceosomal snRNPs and their role in pre-mRNA splicing. Biochim. Biophys. Acta 1990, 1087, 265–292. [Google Scholar] [CrossRef]
- Legerlotz, K.; Smith, H.K. Role of MyoD in denervated, disused, and exercised muscle. Muscle Nerve 2008, 38, 1087–1100. [Google Scholar] [CrossRef] [PubMed]
- Seale, P.; Sabourin, L.A.; Girgis-Gabardo, A.; Mansouri, A.; Gruss, P.; Rudnicki, M.A. Pax7 is required for the specification of myogenic satellite cells. Cell 2000, 102, 777–786. [Google Scholar] [CrossRef] [Green Version]
- Fakan, S. Ultrastructural cytochemical analyses of nuclear functional architecture. Eur. J. Histochem. 2004, 48, 5–14. [Google Scholar] [PubMed]
- Biggiogera, M.; Cisterna, B.; Spedito, A.; Vecchio, L.; Malatesta, M. Perichromatin fibrils as early markers of transcriptional alterations. Differentiation 2008, 76, 57–65. [Google Scholar] [CrossRef]
- Cisterna, B.; Giagnacovo, M.; Costanzo, M.; Fattoretti, P.; Zancanaro, C.; Pellicciari, C.; Malatesta, M. Adapted physical exercise enhances activation and differentiation potential of satellite cells in the skeletal muscle of old mice. J. Anat. 2016, 228, 771–783. [Google Scholar] [CrossRef]
- Puvion-Dutilleul, F.; Puvion, E. Relationship between chromatin and perichromatin granules in cadmium-treated isolated hepatocytes. J. Ultrastruct. Res. 1981, 74, 341–350. [Google Scholar] [CrossRef]
- Malatesta, M.; Biggiogera, M.; Cisterna, B.; Balietti, M.; Bertoni-Freddari, C.; Fattoretti, P. Perichromatin fibrils accumulation in hepatocyte nuclei reveals alterations of pre-mRNA processing during aging. Dna Cell Biol. 2010, 29, 49–57. [Google Scholar] [CrossRef]
- Zancanaro, C.; Malatesta, M.; Vogel, P.; Osculati, F.; Fakan, S. Ultrastructural and morphomentrical analyses of the brown adipocyte nucleus in a hibernating dormouse. Biol. Cell 1993, 79, 55–61. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malatesta, M.; Costanzo, M.; Cisterna, B.; Zancanaro, C. Satellite Cells in Skeletal Muscle of the Hibernating Dormouse, a Natural Model of Quiescence and Re-Activation: Focus on the Cell Nucleus. Cells 2020, 9, 1050. https://doi.org/10.3390/cells9041050
Malatesta M, Costanzo M, Cisterna B, Zancanaro C. Satellite Cells in Skeletal Muscle of the Hibernating Dormouse, a Natural Model of Quiescence and Re-Activation: Focus on the Cell Nucleus. Cells. 2020; 9(4):1050. https://doi.org/10.3390/cells9041050
Chicago/Turabian StyleMalatesta, Manuela, Manuela Costanzo, Barbara Cisterna, and Carlo Zancanaro. 2020. "Satellite Cells in Skeletal Muscle of the Hibernating Dormouse, a Natural Model of Quiescence and Re-Activation: Focus on the Cell Nucleus" Cells 9, no. 4: 1050. https://doi.org/10.3390/cells9041050
APA StyleMalatesta, M., Costanzo, M., Cisterna, B., & Zancanaro, C. (2020). Satellite Cells in Skeletal Muscle of the Hibernating Dormouse, a Natural Model of Quiescence and Re-Activation: Focus on the Cell Nucleus. Cells, 9(4), 1050. https://doi.org/10.3390/cells9041050