SIV Infection and the HIV Proteins Tat and Nef Induce Senescence in Adipose Tissue and Human Adipose Stem Cells, Resulting in Adipocyte Dysfunction

 , , and
, , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Macaque Infection and Sample Collection
2.2. Isolation of ASCs, Cell Culture, and Cell Treatments
2.3. Evaluation of Cell Proliferation
2.4. Adipocyte Differentiation
2.5. Cellular Senescence Assay
2.6. Protein Secretion Assay
2.7. Assays of Oxidative Stress and Mitochondrial Dysfunction
2.8. RNA Isolation and Quantitative RT-PCR
2.9. Protein Extraction and Western Blotting
2.10. Statistical Analysis
3. Results
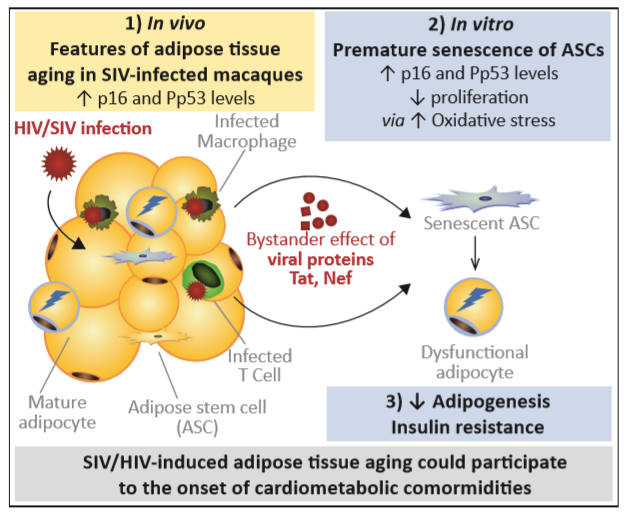
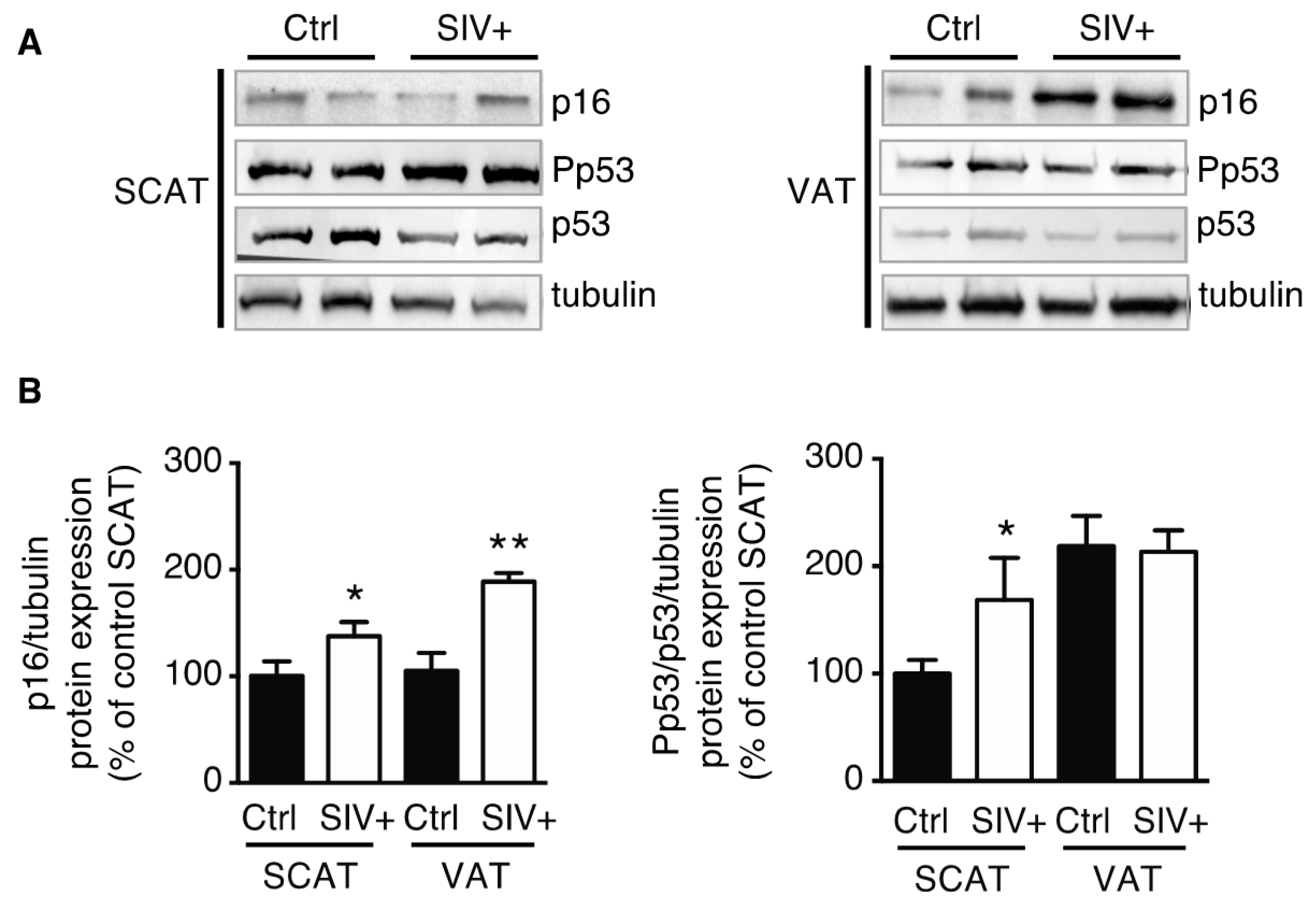
3.1. Adipose Tissue in SIV-Infected Macaques Displays Features of Senescence
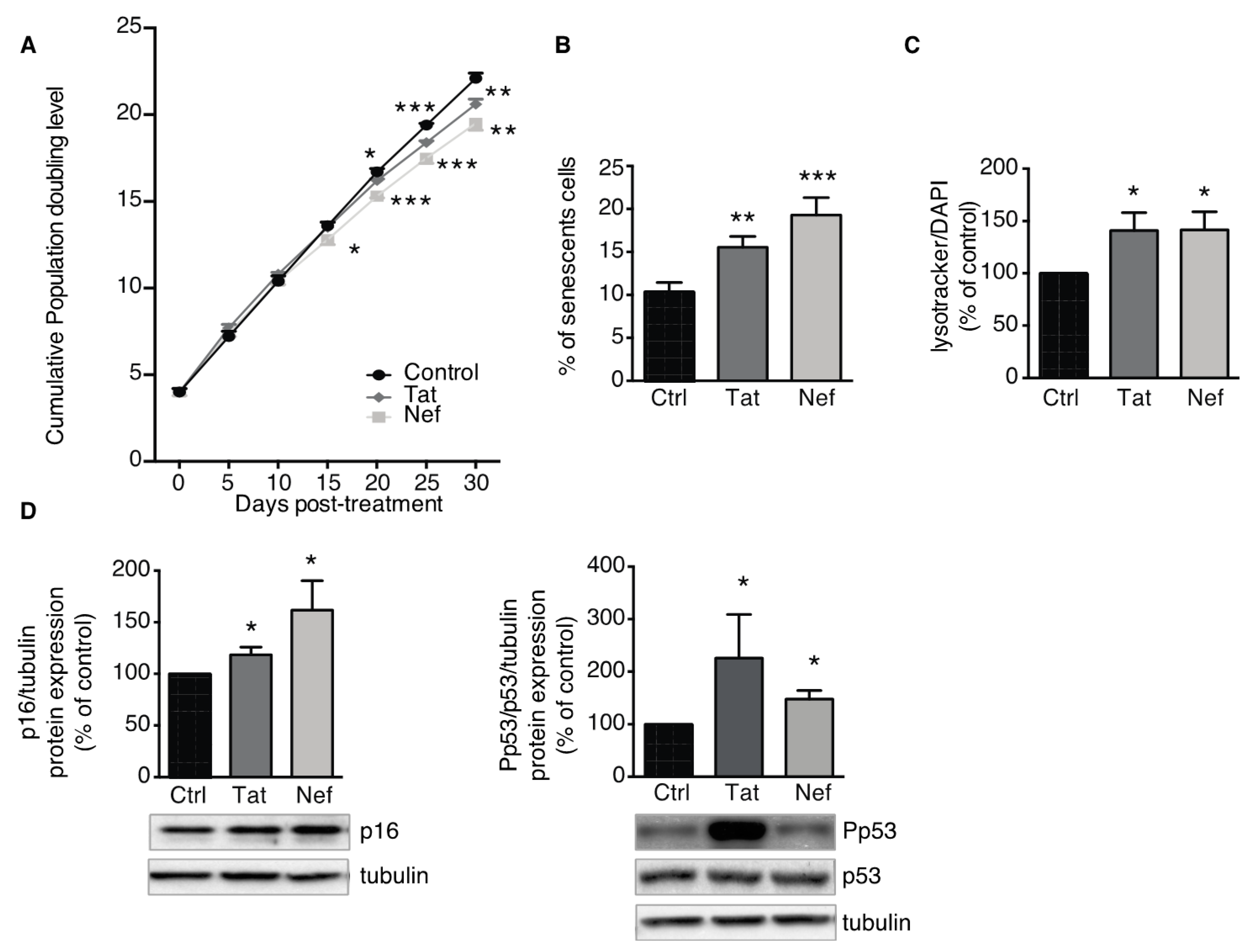
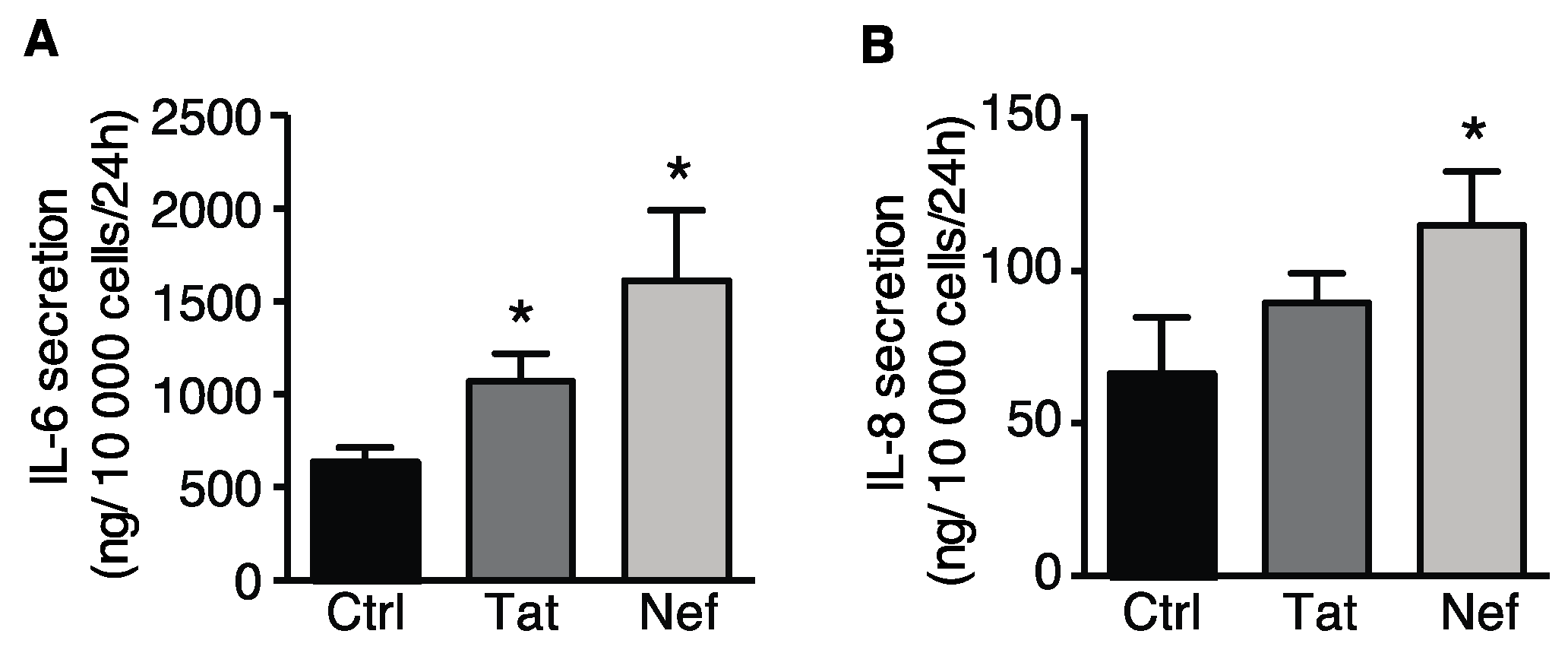
3.2. Tat- and Nef-Induced Cell Senescence in ASCs
3.2.1. Treatment of ASCs with Tat and Nef Resulted in a Lower Proliferative Capacity and Higher Levels of Senescence Markers
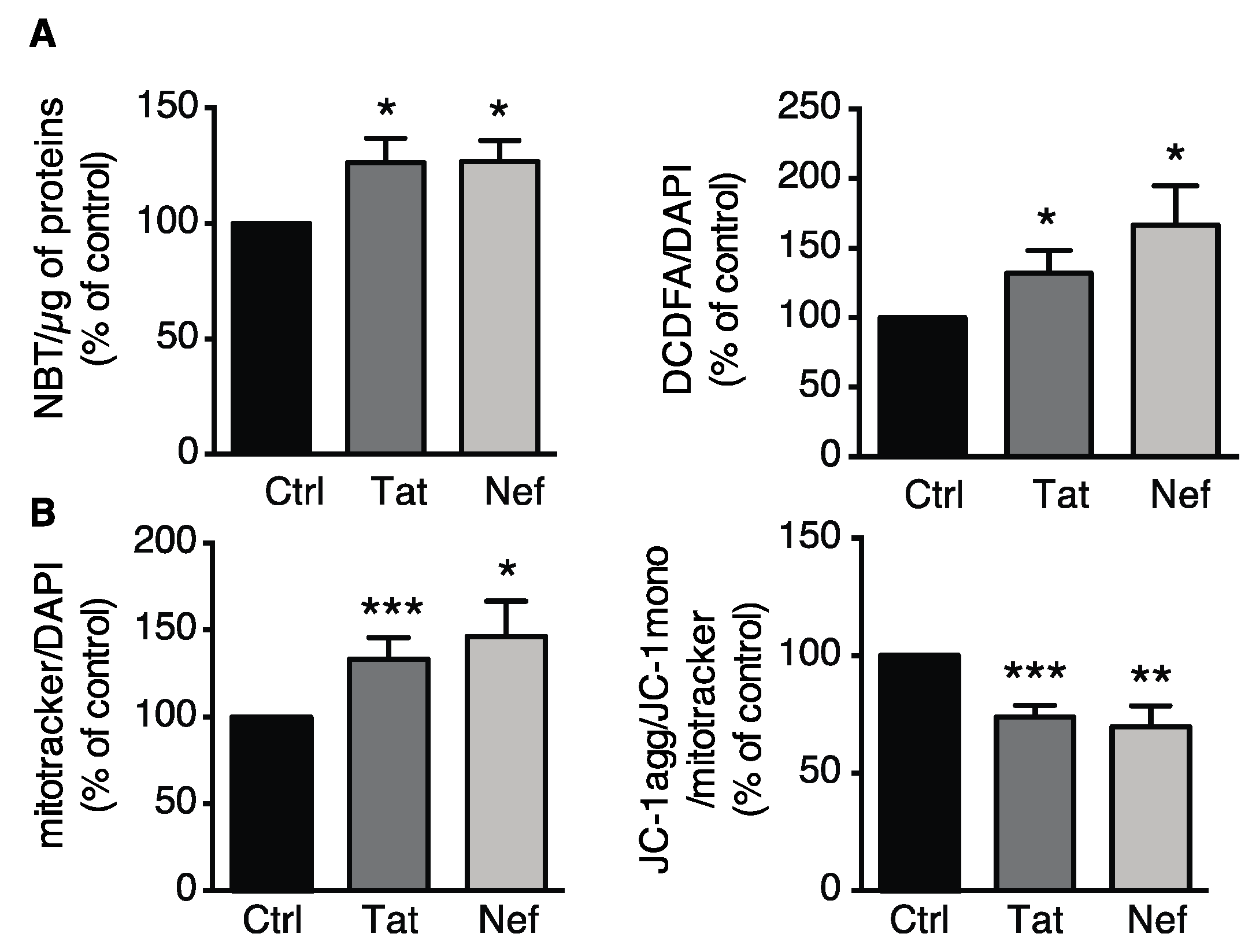
3.2.2. Tat- or Nef-Induced Oxidative Stress and Mitochondrial Dysfunction in ASCs
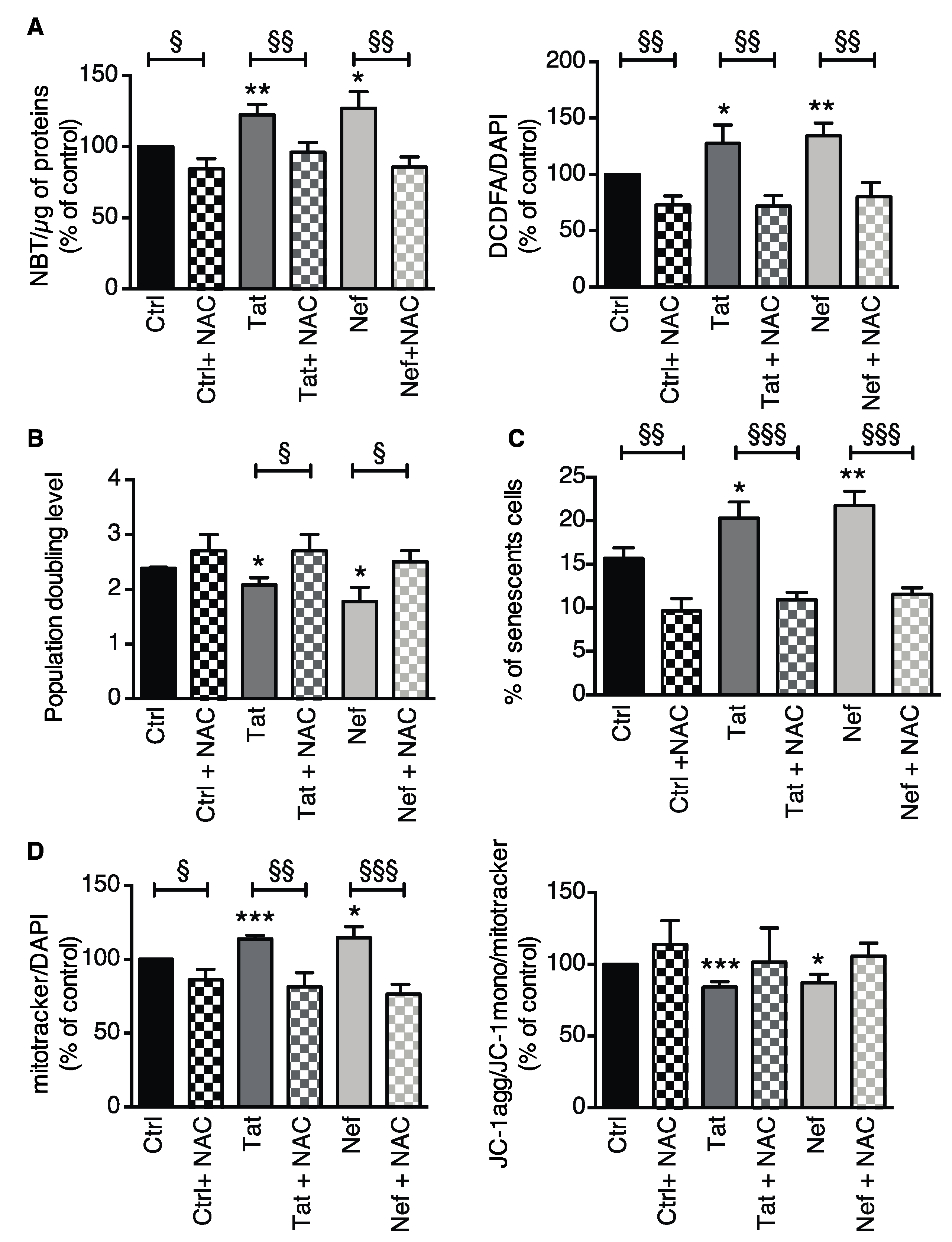
3.2.3. Inhibition of Oxidative Stress Prevented Tat- and Nef-Induced Senescence in ASCs
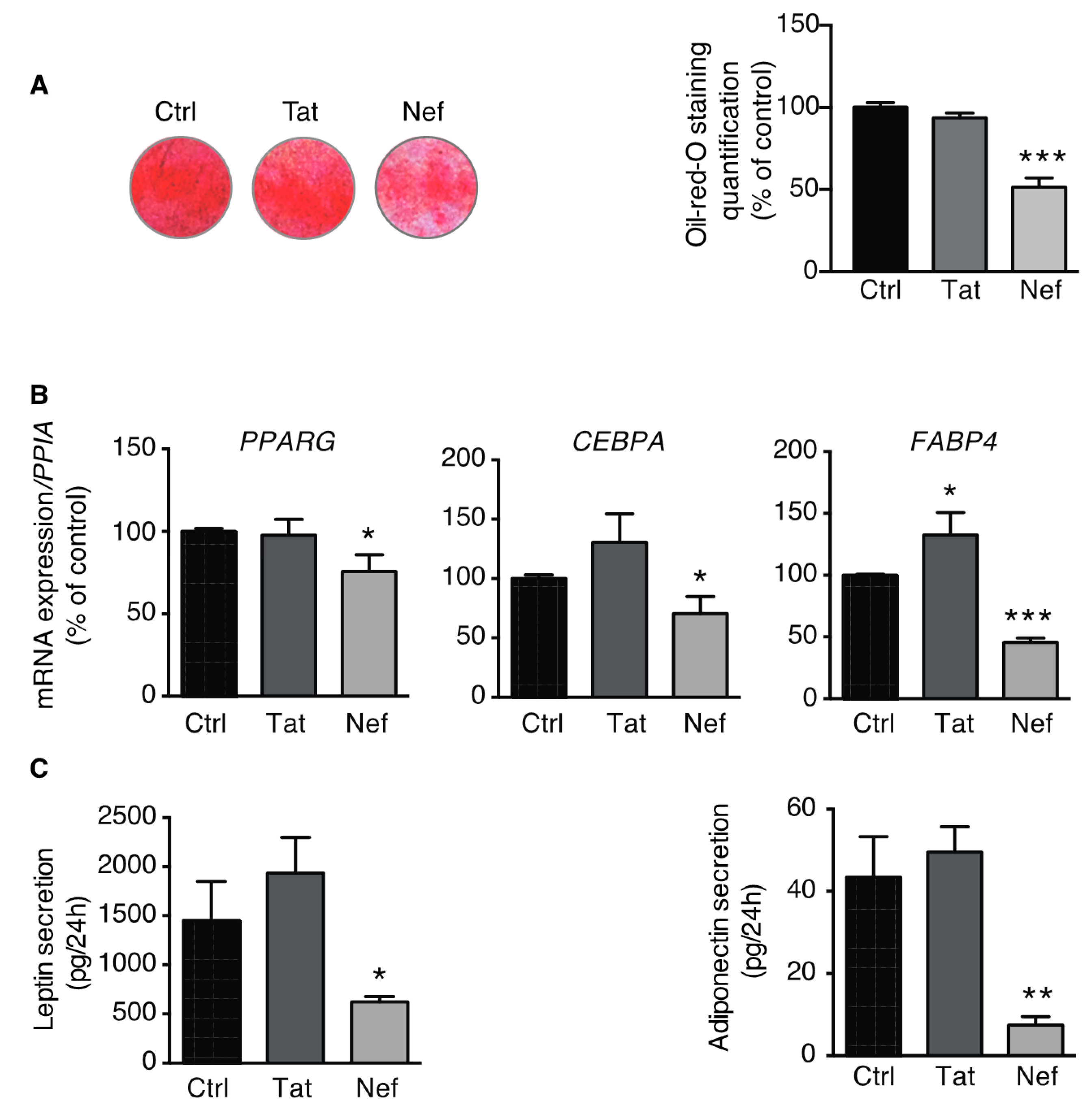
3.3. Adipogenesis is Impaired in ASCs Displaying Nef-Induced Senescence
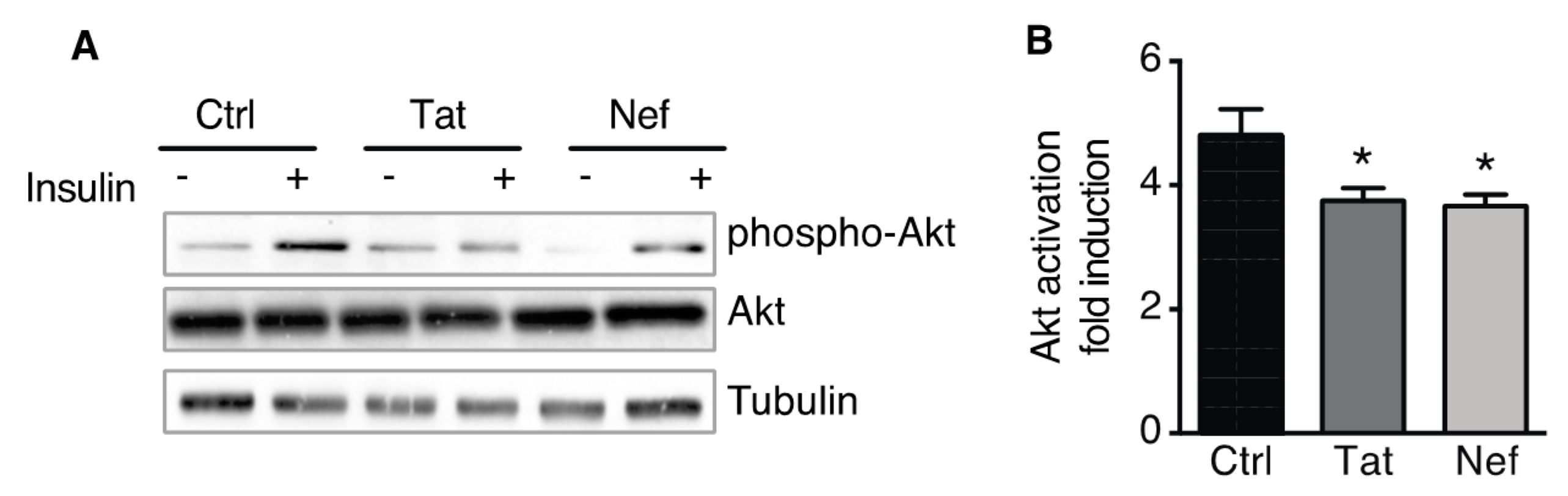
3.4. Treatment of ASCs with Nef or Tat is Associated with Insulin Resistance in ASC-Derived Adipocytes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stout, M.B.; Justice, J.N.; Nicklas, B.J.; Kirkland, J.L. Physiological Aging: Links Among Adipose Tissue Dysfunction, Diabetes, and Frailty. Physiology (Bethesda) 2017, 32, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Lagathu, C.; Cossarizza, A.; Bereziat, V.; Nasi, M.; Capeau, J.; Pinti, M. Basic science and pathogenesis of ageing with HIV: Potential mechanisms and biomarkers. AIDS (London, UK) 2017, 31 (Suppl. 2), S105–S119. [Google Scholar] [CrossRef] [PubMed]
- Caron-Debarle, M.; Lagathu, C.; Boccara, F.; Vigouroux, C.; Capeau, J. HIV-associated lipodystrophy: From fat injury to premature aging. Trends Mol. Med. 2010, 16, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Lagathu, C.; Bereziat, V.; Gorwood, J.; Fellahi, S.; Bastard, J.P.; Vigouroux, C.; Boccara, F.; Capeau, J. Metabolic complications affecting adipose tissue, lipid and glucose metabolism associated with HIV antiretroviral treatment. Expert Opin. Drug Saf. 2019, 18, 829–840. [Google Scholar] [CrossRef]
- Gelpi, M.; Afzal, S.; Lundgren, J.; Ronit, A.; Roen, A.; Mocroft, A.; Gerstoft, J.; Lebech, A.M.; Lindegaard, B.; Kofoed, K.F.; et al. Higher Risk of Abdominal Obesity, Elevated Low-Density Lipoprotein Cholesterol, and Hypertriglyceridemia, but not of Hypertension, in People Living with Human Immunodeficiency Virus (HIV): Results from the Copenhagen Comorbidity in HIV Infection Study. Clin. Infect. Dis. 2018, 67, 579–586. [Google Scholar] [CrossRef]
- Utay, N.S.; Kitch, D.W.; Yeh, E.; Fichtenbaum, C.J.; Lederman, M.M.; Estes, J.D.; Deleage, C.; Magyar, C.; Nelson, S.D.; Klingman, K.L.; et al. Telmisartan Therapy Does Not Improve Lymph Node or Adipose Tissue Fibrosis More Than Continued Antiretroviral Therapy Alone. J. Infect. Dis. 2018, 217, 1770–1781. [Google Scholar] [CrossRef] [Green Version]
- Bastard, J.P.; Caron, M.; Vidal, H.; Jan, V.; Auclair, M.; Vigouroux, C.; Luboinski, J.; Laville, M.; Maachi, M.; Girard, P.M.; et al. Association between altered expression of adipogenic factor SREBP1 in lipoatrophic adipose tissue from HIV-1-infected patients and abnormal adipocyte differentiation and insulin resistance. Lancet 2002, 359, 1026–1031. [Google Scholar] [CrossRef]
- Bereziat, V.; Cervera, P.; Le Dour, C.; Verpont, M.C.; Dumont, S.; Vantyghem, M.C.; Capeau, J.; Vigouroux, C.; Lipodystrophy Study, G. LMNA mutations induce a non-inflammatory fibrosis and a brown fat-like dystrophy of enlarged cervical adipose tissue. Am. J. Pathol. 2011, 179, 2443–2453. [Google Scholar] [CrossRef]
- Gorwood, J.; Bourgeois, C.; Mantecon, M.; Atlan, M.; Pourcher, V.; Pourcher, G.; Le Grand, R.; Desjardins, D.; Feve, B.; Lambotte, O.; et al. Impact of HIV/simian immunodeficiency virus infection and viral proteins on adipose tissue fibrosis and adipogenesis. AIDS (London, UK) 2019, 33, 953–964. [Google Scholar] [CrossRef]
- Hayflick, L.; Moorhead, P.S. The serial cultivation of human diploid cell strains. Exp. Cell Res. 1961, 25, 585–621. [Google Scholar] [CrossRef]
- Tchkonia, T.; Morbeck, D.E.; Von Zglinicki, T.; Van Deursen, J.; Lustgarten, J.; Scrable, H.; Khosla, S.; Jensen, M.D.; Kirkland, J.L. Fat tissue, aging, and cellular senescence. Aging Cell 2010, 9, 667–684. [Google Scholar] [CrossRef] [PubMed]
- Rodier, F.; Campisi, J. Four faces of cellular senescence. J. Cell Biol. 2011, 192, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Coppe, J.P.; Patil, C.K.; Rodier, F.; Sun, Y.; Munoz, D.P.; Goldstein, J.; Nelson, P.S.; Desprez, P.Y.; Campisi, J. Senescence-associated secretory phenotypes reveal cell-nonautonomous functions of oncogenic RAS and the p53 tumor suppressor. PLoS Biol. 2008, 6, 2853–2868. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.C.; Yin, P.H.; Chi, C.W.; Wei, Y.H. Increase in mitochondrial mass in human fibroblasts under oxidative stress and during replicative cell senescence. J. Biomed. Sci. 2002, 9, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Afonso, P.; Auclair, M.; Boccara, F.; Vantyghem, M.C.; Katlama, C.; Capeau, J.; Vigouroux, C.; Caron-Debarle, M. LMNA mutations resulting in lipodystrophy and HIV protease inhibitors trigger vascular smooth muscle cell senescence and calcification: Role of ZMPSTE24 downregulation. Atherosclerosis 2016, 245, 200–211. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Vallejo, S.J.; Beaupere, C.; Larghero, J.; Capeau, J.; Lagathu, C. HIV protease inhibitors induce senescence and alter osteoblastic potential of human bone marrow mesenchymal stem cells: Beneficial effect of pravastatin. Aging Cell 2013, 12, 955–965. [Google Scholar] [CrossRef]
- Caron, M.; Auclair, M.; Donadille, B.; Bereziat, V.; Guerci, B.; Laville, M.; Narbonne, H.; Bodemer, C.; Lascols, O.; Capeau, J.; et al. Human lipodystrophies linked to mutations in A-type lamins and to HIV protease inhibitor therapy are both associated with prelamin A accumulation, oxidative stress and premature cellular senescence. Cell Death Differ. 2007, 14, 1759–1767. [Google Scholar] [CrossRef]
- Fiume, G.; Vecchio, E.; De Laurentiis, A.; Trimboli, F.; Palmieri, C.; Pisano, A.; Falcone, C.; Pontoriero, M.; Rossi, A.; Scialdone, A.; et al. Human immunodeficiency virus-1 That activates NF-kappaB via physical interaction with IkappaB-alpha and p65. Nucleic Acids Res. 2012, 40, 3548–3562. [Google Scholar] [CrossRef] [Green Version]
- Beaupere, C.; Garcia, M.; Larghero, J.; Feve, B.; Capeau, J.; Lagathu, C. The HIV proteins Tat and Nef promote human bone marrow mesenchymal stem cell senescence and alter osteoblastic differentiation. Aging Cell 2015, 14, 534–546. [Google Scholar] [CrossRef]
- Damouche, A.; Lazure, T.; Avettand-Fenoel, V.; Huot, N.; Dejucq-Rainsford, N.; Satie, A.P.; Melard, A.; David, L.; Gommet, C.; Ghosn, J.; et al. Adipose Tissue Is a Neglected Viral Reservoir and an Inflammatory Site during Chronic HIV and SIV Infection. PLoS Pathog. 2015, 11, e1005153. [Google Scholar] [CrossRef] [Green Version]
- Couturier, J.; Suliburk, J.W.; Brown, J.M.; Luke, D.J.; Agarwal, N.; Yu, X.; Nguyen, C.; Iyer, D.; Kozinetz, C.A.; Overbeek, P.A.; et al. Human adipose tissue as a reservoir for memory CD4+ T cells and HIV. AIDS (London, UK) 2015, 29, 667–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boya, P.; Pauleau, A.L.; Poncet, D.; Gonzalez-Polo, R.A.; Zamzami, N.; Kroemer, G. Viral proteins targeting mitochondria: Controlling cell death. Biochim. Biophys. Acta 2004, 1659, 178–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raymond, A.D.; Campbell-Sims, T.C.; Khan, M.; Lang, M.; Huang, M.B.; Bond, V.C.; Powell, M.D. HIV Type 1 Nef is released from infected cells in CD45(+) microvesicles and is present in the plasma of HIV-infected individuals. AIDS Res. Hum. Retrovir. 2011, 27, 167–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gougeon, M.L. Apoptosis as an HIV strategy to escape immune attack. Nat. Rev. Immunol. 2003, 3, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Lagathu, C.; Eustace, B.; Prot, M.; Frantz, D.; Gu, Y.; Bastard, J.P.; Maachi, M.; Azoulay, S.; Briggs, M.; Caron, M.; et al. Some HIV antiretrovirals increase oxidative stress and alter chemokine, cytokine or adiponectin production in human adipocytes and macrophages. Antivir. Ther. 2007, 12, 489–500. [Google Scholar]
- Couturier, J.; Agarwal, N.; Nehete, P.N.; Baze, W.B.; Barry, M.A.; Jagannadha Sastry, K.; Balasubramanyam, A.; Lewis, D.E. Infectious SIV resides in adipose tissue and induces metabolic defects in chronically infected rhesus macaques. Retrovirology 2016, 13, 30. [Google Scholar] [CrossRef] [Green Version]
- Campisi, J. Parsing p53 Transactivation. Dev. Cell 2011, 20, 573–574. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, A.K.; O’Brien, M.; Mau, T.; Qi, N.; Yung, R. Adipose Tissue Senescence and Inflammation in Aging is Reversed by the Young Milieu. J. Gerontol. Ser. A 2019, 74, 1709–1715. [Google Scholar] [CrossRef]
- Schutz, E.; Gogiraju, R.; Pavlaki, M.; Drosos, I.; Georgiadis, G.S.; Argyriou, C.; Rim Ben Hallou, A.; Konstantinou, F.; Mikroulis, D.; Schuler, R.; et al. Age-Dependent and -Independent Effects of Perivascular Adipose Tissue and Its Paracrine Activities during Neointima Formation. Int. J. Mol. Sci. 2019, 21, 282. [Google Scholar] [CrossRef] [Green Version]
- Tugizov, S.M.; Herrera, R.; Chin-Hong, P.; Veluppillai, P.; Greenspan, D.; Michael Berry, J.; Pilcher, C.D.; Shiboski, C.H.; Jay, N.; Rubin, M.; et al. HIV-associated disruption of mucosal epithelium facilitates paracellular penetration by human papillomavirus. Virology 2013, 446, 378–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, M.K.; Kaminski, R.; Mullen, B.; Gordon, J.; Burdo, T.H.; Cheung, J.Y.; Feldman, A.M.; Madesh, M.; Khalili, K. HIV-1 Nef-induced cardiotoxicity through dysregulation of autophagy. Sci. Rep. 2017, 7, 8572. [Google Scholar] [CrossRef] [PubMed]
- Hijmans, J.G.; Stockleman, K.; Reiakvam, W.; Levy, M.V.; Brewster, L.M.; Bammert, T.D.; Greiner, J.J.; Connick, E.; DeSouza, C.A. Effects of HIV-1 gp120 and tat on endothelial cell sensescence and senescence-associated microRNAs. Physiol. Rep. 2018, 6, e13647. [Google Scholar] [CrossRef] [PubMed]
- Maredziak, M.; Marycz, K.; Tomaszewski, K.A.; Kornicka, K.; Henry, B.M. The Influence of Aging on the Regenerative Potential of Human Adipose Derived Mesenchymal Stem Cells. Stem Cells Int. 2016, 2016, 2152435. [Google Scholar] [CrossRef]
- Garrabou, G.; Lopez, S.; Moren, C.; Martinez, E.; Fontdevila, J.; Cardellach, F.; Gatell, J.M.; Miro, O. Mitochondrial damage in adipose tissue of untreated HIV-infected patients. AIDS (London, UK) 2011, 25, 165–170. [Google Scholar] [CrossRef]
- Giralt, M.; Domingo, P.; Guallar, J.P.; Rodriguez de la Concepcion, M.L.; Alegre, M.; Domingo, J.C.; Villarroya, F. HIV-1 infection alters gene expression in adipose tissue, which contributes to HIV- 1/HAART-associated lipodystrophy. Antivir. Ther. 2006, 11, 729–740. [Google Scholar]
- Westendorp, M.O.; Shatrov, V.A.; Schulze-Osthoff, K.; Frank, R.; Kraft, M.; Los, M.; Krammer, P.H.; Droge, W.; Lehmann, V. HIV-1 Tat potentiates TNF-induced NF-kappa B activation and cytotoxicity by altering the cellular redox state. EMBO J. 1995, 14, 546–554. [Google Scholar] [CrossRef]
- Rozzi, S.J.; Avdoshina, V.; Fields, J.A.; Trejo, M.; Ton, H.T.; Ahern, G.P.; Mocchetti, I. Human Immunodeficiency Virus Promotes Mitochondrial Toxicity. Neurotox Res. 2017, 32, 723–733. [Google Scholar] [CrossRef]
- Lacasa, D.; Taleb, S.; Keophiphath, M.; Miranville, A.; Clement, K. Macrophage-secreted factors impair human adipogenesis: Involvement of proinflammatory state in preadipocytes. Endocrinology 2007, 148, 868–877. [Google Scholar] [CrossRef] [Green Version]
- Vila, I.K.; Badin, P.M.; Marques, M.A.; Monbrun, L.; Lefort, C.; Mir, L.; Louche, K.; Bourlier, V.; Roussel, B.; Gui, P.; et al. Immune cell Toll-like receptor 4 mediates the development of obesity- and endotoxemia-associated adipose tissue fibrosis. Cell Rep. 2014, 7, 1116–1129. [Google Scholar] [CrossRef]
- Okuno, Y.; Fukuhara, A.; Hashimoto, E.; Kobayashi, H.; Kobayashi, S.; Otsuki, M.; Shimomura, I. Oxidative Stress Inhibits Healthy Adipose Expansion Through Suppression of SREBF1-Mediated Lipogenic Pathway. Diabetes 2018, 67, 1113–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansilla, E.; Diaz Aquino, V.; Zambon, D.; Marin, G.H.; Martire, K.; Roque, G.; Ichim, T.; Riordan, N.H.; Patel, A.; Sturla, F.; et al. Could metabolic syndrome, lipodystrophy, and aging be mesenchymal stem cell exhaustion syndromes? Stem Cells Int. 2011, 2011, 943216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caso, G.; McNurlan, M.A.; Mileva, I.; Zemlyak, A.; Mynarcik, D.C.; Gelato, M.C. Peripheral fat loss and decline in adipogenesis in older humans. Metab. Clin. Exp. 2013, 62, 337–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karagiannides, I.; Tchkonia, T.; Dobson, D.E.; Steppan, C.M.; Cummins, P.; Chan, G.; Salvatori, K.; Hadzopoulou-Cladaras, M.; Kirkland, J.L. Altered expression of C/EBP family members results in decreased adipogenesis with aging. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 280, R1772–R1780. [Google Scholar] [CrossRef]
- Prost, S.; Le Dantec, M.; Auge, S.; Le Grand, R.; Derdouch, S.; Auregan, G.; Deglon, N.; Relouzat, F.; Aubertin, A.M.; Maillere, B.; et al. Human and simian immunodeficiency viruses deregulate early hematopoiesis through a Nef/PPARgamma/STAT5 signaling pathway in macaques. J. Clin. Investig. 2008, 118, 1765–1775. [Google Scholar] [CrossRef]
- Otake, K.; Omoto, S.; Yamamoto, T.; Okuyama, H.; Okada, H.; Okada, N.; Kawai, M.; Saksena, N.K.; Fujii, Y.R. HIV-1 Nef protein in the nucleus influences adipogenesis as well as viral transcription through the peroxisome proliferator-activated receptors. AIDS (London, UK) 2004, 18, 189–198. [Google Scholar] [CrossRef]
- Cotter, E.J.; Chew, N.; Powderly, W.G.; Doran, P.P. HIV type 1 alters mesenchymal stem cell differentiation potential and cell phenotype ex vivo. AIDS Res. Hum. Retrovir. 2011, 27, 187–199. [Google Scholar] [CrossRef]
- Diaz-Delfin, J.; Domingo, P.; Wabitsch, M.; Giralt, M.; Villarroya, F. HIV-1 Tat protein impairs adipogenesis and induces the expression and secretion of proinflammatory cytokines in human SGBS adipocytes. Antivir. Ther. 2012, 17, 529–540. [Google Scholar] [CrossRef] [Green Version]
- Hulgan, T.; Ramsey, B.S.; Koethe, J.R.; Samuels, D.C.; Gerschenson, M.; Libutti, D.E.; Sax, P.E.; Daar, E.S.; McComsey, G.A.; Brown, T.T. Relationships Between Adipose Mitochondrial Function, Serum Adiponectin, and Insulin Resistance in Persons with HIV After 96 Weeks of Antiretroviral Therapy. J. Acquir. Immune Defic. Syndr. 2019, 80, 358–366. [Google Scholar] [CrossRef]
- Kuller, L.H.; Tracy, R.; Belloso, W.; De Wit, S.; Drummond, F.; Lane, H.C.; Ledergerber, B.; Lundgren, J.; Neuhaus, J.; Nixon, D.; et al. Inflammatory and coagulation biomarkers and mortality in patients with HIV infection. PLoS Med. 2008, 5, e203. [Google Scholar] [CrossRef]
- Hulgan, T. Factors Associated with Insulin Resistance in Adults with HIV Receiving Contemporary Antiretroviral Therapy: A Brief Update. Curr. HIV/AIDS Rep. 2018, 15, 223–232. [Google Scholar] [CrossRef]
- Herrin, M.; Tate, J.P.; Akgun, K.M.; Butt, A.A.; Crothers, K.; Freiberg, M.S.; Gibert, C.L.; Leaf, D.A.; Rimland, D.; Rodriguez-Barradas, M.C.; et al. Weight Gain and Incident Diabetes Among HIV-Infected Veterans Initiating Antiretroviral Therapy Compared with Uninfected Individuals. J. Acquir. Immune Defic. Syndr. 2016, 73, 228–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanna, D.B.; Post, W.S.; Deal, J.A.; Hodis, H.N.; Jacobson, L.P.; Mack, W.J.; Anastos, K.; Gange, S.J.; Landay, A.L.; Lazar, J.M.; et al. HIV Infection Is Associated with Progression of Subclinical Carotid Atherosclerosis. Clin. Infect. Dis. 2015, 61, 640–650. [Google Scholar] [CrossRef] [Green Version]
- Hsue, P.Y.; Hunt, P.W.; Schnell, A.; Kalapus, S.C.; Hoh, R.; Ganz, P.; Martin, J.N.; Deeks, S.G. Role of viral replication, antiretroviral therapy, and immunodeficiency in HIV-associated atherosclerosis. AIDS (London, UK) 2009, 23, 1059–1067. [Google Scholar] [CrossRef] [PubMed]
- Caron, M.; Auclairt, M.; Vissian, A.; Vigouroux, C.; Capeau, J. Contribution of mitochondrial dysfunction and oxidative stress to cellular premature senescence induced by antiretroviral thymidine analogues. Antivir. Ther. 2008, 13, 27–38. [Google Scholar] [PubMed]
- Lefevre, C.; Auclair, M.; Boccara, F.; Bastard, J.P.; Capeau, J.; Vigouroux, C.; Caron-Debarle, M. Premature senescence of vascular cells is induced by HIV protease inhibitors: Implication of prelamin A and reversion by statin. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Auclair, M.; Afonso, P.; Capel, E.; Caron-Debarle, M.; Capeau, J. Impact of darunavir, atazanavir and lopinavir boosted with ritonavir on cultured human endothelial cells: Beneficial effect of pravastatin. Antivir. Ther. 2014, 19, 773–782. [Google Scholar] [CrossRef] [Green Version]
- Afonso, P.; Auclair, M.; Caron-Debarle, M.; Capeau, J. Impact of CCR5, integrase and protease inhibitors on human endothelial cell function, stress, inflammation and senescence. Antivir. Ther. 2017, 10.3851/IMP3160. [Google Scholar] [CrossRef] [Green Version]
- Auclair, M.; Guenantin, A.C.; Fellahi, S.; Garcia, M.; Capeau, J. HIV antiretroviral drugs, dolutegravir, maraviroc and ritonavir-boosted atazanavir use different pathways to affect inflammation, senescence and insulin sensitivity in human coronary endothelial cells. PLoS ONE 2020, 15, e0226924. [Google Scholar] [CrossRef] [PubMed]
- Gorwood, J.; Bourgeois, C.; Pourcher, V.; Pourcher, G.; Charlotte, F.; Mantecon, M.; Rose, C.; Morichon, R.; Atlan, M.; Le Grand, R.; et al. The integrase inhibitors dolutegravir and raltegravir exert pro-adipogenic and profibrotic effects and induce insulin resistance in human/simian adipose tissue and human adipocytes. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Venter, W.D.F.; Moorhouse, M.; Sokhela, S.; Fairlie, L.; Mashabane, N.; Masenya, M.; Serenata, C.; Akpomiemie, G.; Qavi, A.; Chandiwana, N.; et al. Dolutegravir plus Two Different Prodrugs of Tenofovir to Treat HIV. N. Engl. J. Med. 2019, 381, 803–815. [Google Scholar] [CrossRef] [PubMed]
- Debroy, P.; Sim, M.; Erlandson, K.M.; Falutz, J.; Prado, C.M.; Brown, T.T.; Guaraldi, G.; Lake, J.E. Progressive increases in fat mass occur in adults living with HIV on antiretroviral therapy, but patterns differ by sex and anatomic depot. J. Antimicrob. Chemother. 2019, 74, 1028–1034. [Google Scholar] [CrossRef] [PubMed]
- Norwood, J.; Turner, M.; Bofill, C.; Rebeiro, P.; Shepherd, B.; Bebawy, S.; Hulgan, T.; Raffanti, S.; Haas, D.W.; Sterling, T.R.; et al. Brief Report: Weight Gain in Persons with HIV Switched from Efavirenz-Based to Integrase Strand Transfer Inhibitor-Based Regimens. J. Acquir. Immune Defic. Syndr. 2017, 76, 527–531. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorwood, J.; Ejlalmanesh, T.; Bourgeois, C.; Mantecon, M.; Rose, C.; Atlan, M.; Desjardins, D.; Le Grand, R.; Fève, B.; Lambotte, O.; et al. SIV Infection and the HIV Proteins Tat and Nef Induce Senescence in Adipose Tissue and Human Adipose Stem Cells, Resulting in Adipocyte Dysfunction. Cells 2020, 9, 854. https://doi.org/10.3390/cells9040854
Gorwood J, Ejlalmanesh T, Bourgeois C, Mantecon M, Rose C, Atlan M, Desjardins D, Le Grand R, Fève B, Lambotte O, et al. SIV Infection and the HIV Proteins Tat and Nef Induce Senescence in Adipose Tissue and Human Adipose Stem Cells, Resulting in Adipocyte Dysfunction. Cells. 2020; 9(4):854. https://doi.org/10.3390/cells9040854
Chicago/Turabian StyleGorwood, Jennifer, Tina Ejlalmanesh, Christine Bourgeois, Matthieu Mantecon, Cindy Rose, Michael Atlan, Delphine Desjardins, Roger Le Grand, Bruno Fève, Olivier Lambotte, and et al. 2020. "SIV Infection and the HIV Proteins Tat and Nef Induce Senescence in Adipose Tissue and Human Adipose Stem Cells, Resulting in Adipocyte Dysfunction" Cells 9, no. 4: 854. https://doi.org/10.3390/cells9040854
APA StyleGorwood, J., Ejlalmanesh, T., Bourgeois, C., Mantecon, M., Rose, C., Atlan, M., Desjardins, D., Le Grand, R., Fève, B., Lambotte, O., Capeau, J., Béréziat, V., & Lagathu, C. (2020). SIV Infection and the HIV Proteins Tat and Nef Induce Senescence in Adipose Tissue and Human Adipose Stem Cells, Resulting in Adipocyte Dysfunction. Cells, 9(4), 854. https://doi.org/10.3390/cells9040854