Spatiotemporal Mislocalization of Nuclear Membrane-Associated Proteins in γ-Irradiation-Induced Senescent Cells




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Cell Culture
2.2. Cell Irradiation
2.3. Antibodies and Immunofluorescence
2.4. Confocal Fluorescence Microscopy of Spatially Fixed Cells
2.5. Senescence-Associated β-Galactosidase Assay
2.6. SDS-PAGE and Western Blot
3. Results
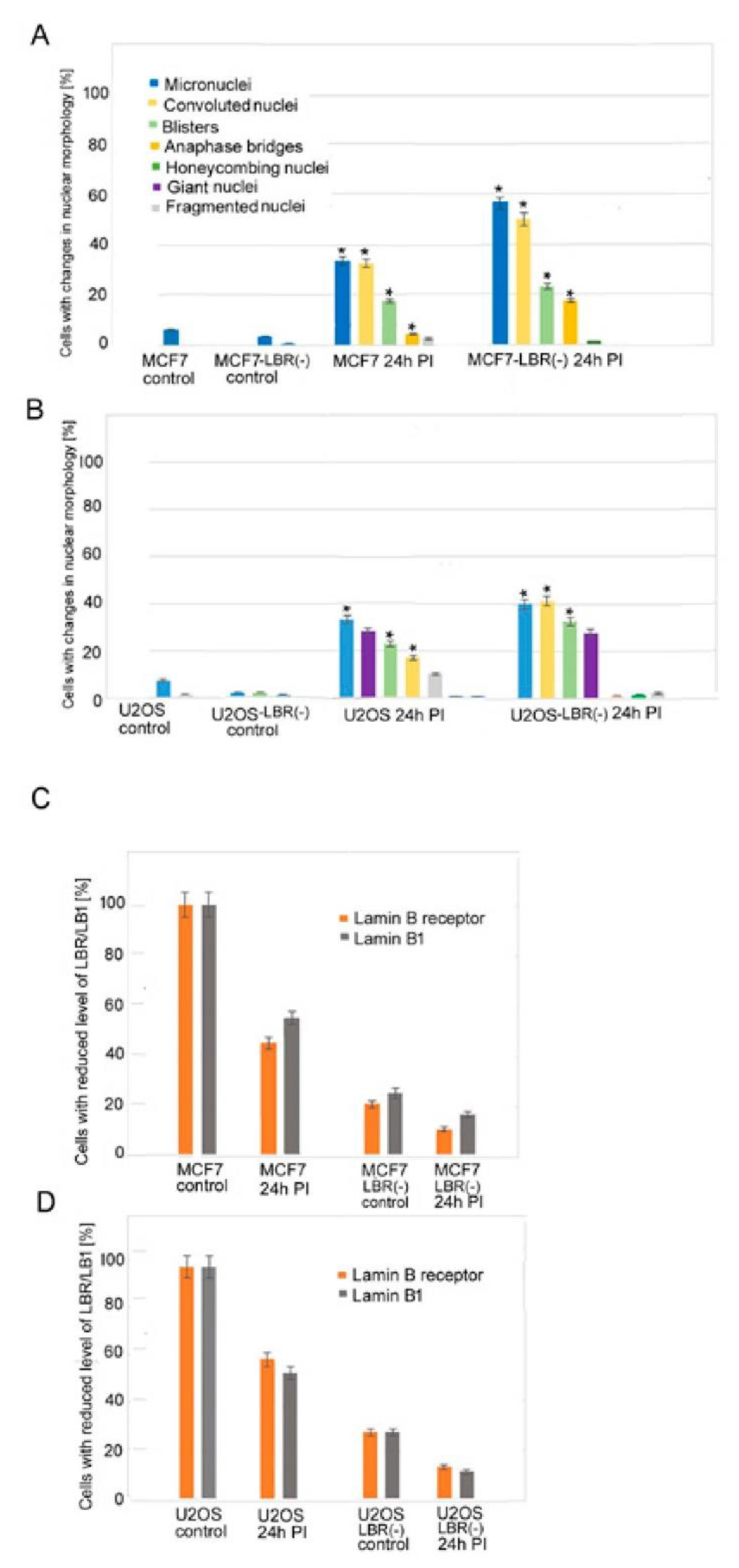
3.1. Changes in Nuclear Morphology of MCF7 and U2OS Cancer Cell Lines after Irradiation
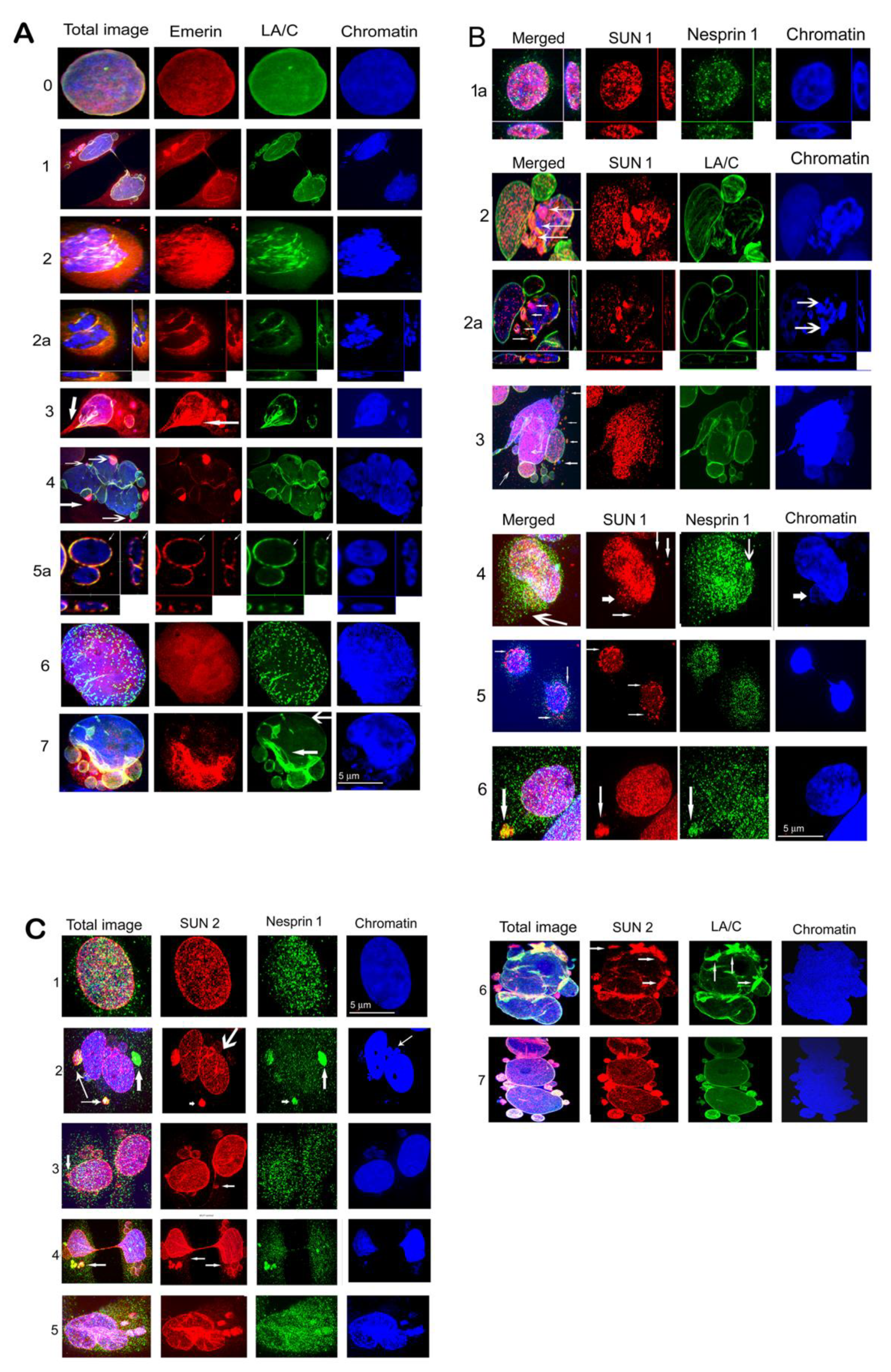
3.2. Defects in Localization of Emerin, LA/C, SUN1, SUN2, and Nesprin-1 Cells Exposed to γ-Irradiation.
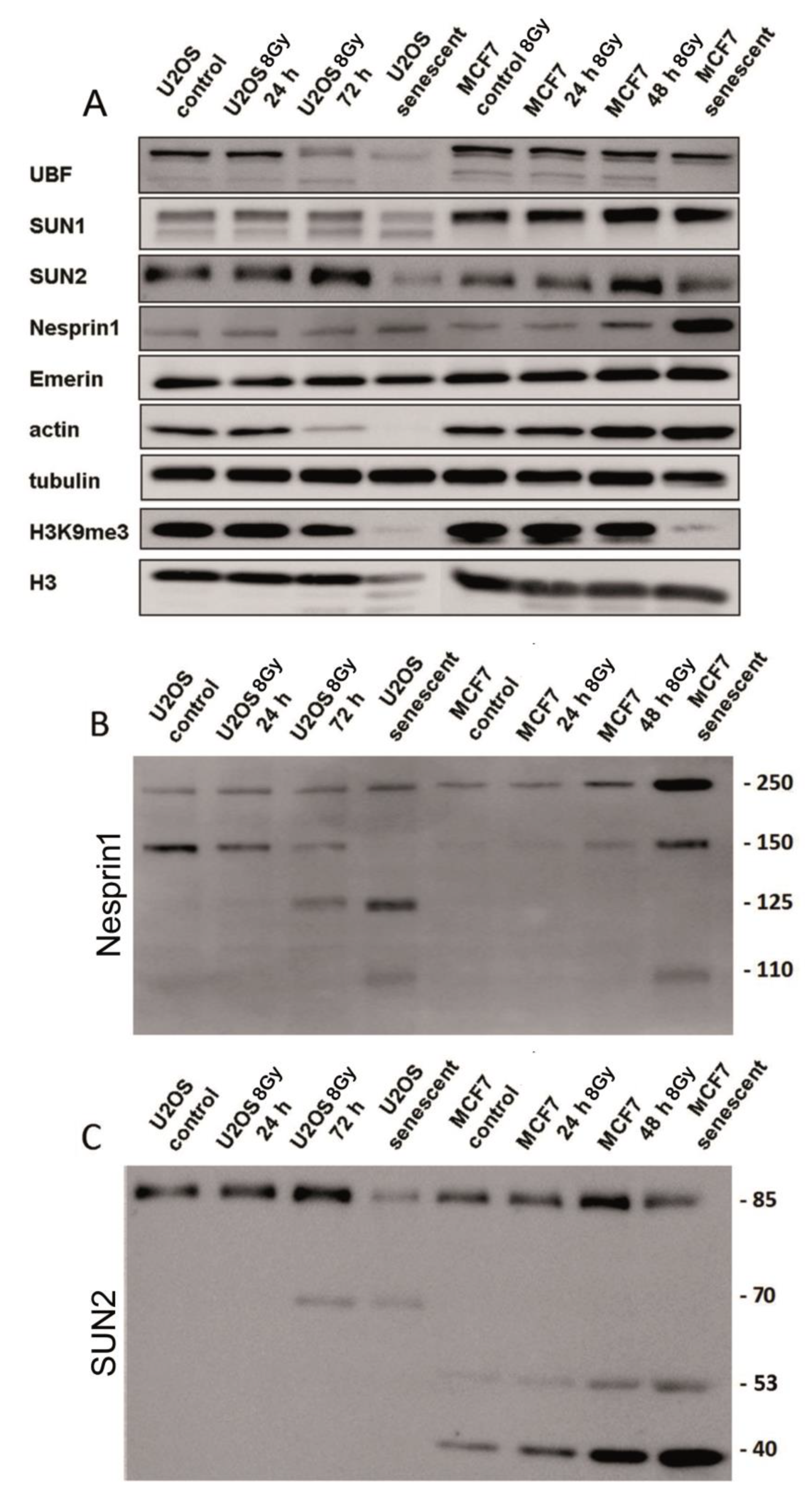
3.3. Expression and Integrity of LINC Proteins Analyzed by Western Blot
4. Discussion
4.1. Early Changes in Localization of LINC Proteins in Irradiated Cells
4.2. LBR/LB1 Deficiency May Enhance LINC Proteins Mobilization in Irradiated Cells
4.3. Possible Clinical Implications of Elevated LINC Protein Mobility in Irradiated Cells
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, Q.; Skepper, J.N.; Yang, F.; Davies, J.D.; Hegyi, L.; Roberts, L.G.; Weissberg, P.L.; Ellis, J.A.; Shanahan, C.M. Nesprins: A novel family of spectrin-repeat-containing proteins that localize to the nuclear membrane in multiple tissues. J. Cell Sci. 2001, 114, 4485–4498. [Google Scholar] [PubMed]
- Rajgor, D.; Mellad, J.A.; Autore, F.; Zhang, Q.; Shanahan, C.M. Multiple novel nesprin-1and nesprin-2 variants act as versatile tissue-specific intracellular scaffolds. PLos ONE 2012, 7, e40098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reigor, D.; Shanahan, C.M. Nesprins: From the nuclear envelope and beyond. Expert Rev. Mol. Med. 2013, 15. [Google Scholar] [CrossRef]
- Crisp, M.; Liu, Q.; Roux, Q.; Rattner, J.B.; Shanahan, C.; Burke, B.; Stahl, P.D.; Hodzic, D. Coupling of the nucleus and the cytoplasm: Role of the LINC complex. J. Cell Biol. 2006, 172, 41–53. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Li, R.; Shanahan, C.M.; Zhang, Q. Nesprin-1/2: Roles in nuclear envelope organization, myogenesis and muscle disease. Biochem. Soc. Trans. 2018, 46, 311–320. [Google Scholar] [CrossRef] [Green Version]
- Dupin, I.; Etienne-Manneville, S. Nuclear positioning mechanisms and functions. Int. J. Biochem. Cell Biol. 2011, 42, 1698–1707. [Google Scholar] [CrossRef] [PubMed]
- Gundersen, G.G.; Worman, H.J. Nuclear positioning. Cell 2013, 152, 1376–1389. [Google Scholar] [CrossRef] [Green Version]
- Shanahan, C.M.; Weissberg, P.L.; Metcalfe, J.C. Isolation of gene markers differentiating and proliferating vascular smooth muscle cells. Circ. Res. 1993, 73, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Gotzman, J.; Foisner, R.P. Lamins and Emerin in muscular dystrophy: The nuclear envelope connection. In Madame Curie Bioscience Database; Landes Bioscience: Austin, TX, USA, 2013; Volume 222. [Google Scholar]
- Zhang, J.; Felder, A.; Liu, Y.; Guo, L.T.; Lange, S.; Dalton, N.D. Nesprin 1 is critical for nuclear positioning and anchorage. Hum. Mol. Genet. 2010, 19, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.K.; Haraguchi, T.; Lee, R.S.; Koujin, T.; Hiaoka, Y.; Wilson, K.L. Distinct functional domains in emerin bind lamin A and DNA binding protein BAF. J. Cell. Sci. 2001, 114, 4567–4573. [Google Scholar] [PubMed]
- Stierlé, V.; Couprie, J.; Ostlund, C.; Krimm, I.; Zinn-Justin, S.; Hossenlopp, P. Worman, H.J.; Courvalin, J.C.; Duband-Goulet, I. The carboxyl-terminal region common to lamins A and C contains a DNA binding domain. Biochemistry 2003, 42, 4819–4828. [Google Scholar] [CrossRef] [PubMed]
- Duong, N.T.; Morris, G.E.; Lam, L.T.; Zhang, Q.; Sewry, C.A.; Shanahan, C.M.; Holt, I. Nesprins: tissue-specific expression of epsilon and other short isoforms. PLoS ONE 2014, 9, e94380. [Google Scholar] [CrossRef] [Green Version]
- Meinke, P.; Nguyen, T.D.; Wehnert, M.S. The LINC complex and human disease. Biochem. Soc. Trans. 2011, 39, 1693–1697. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Bethmann, C.; Worth, N.F.; Davies, J.D.; Wasner, C.; Feuer, A.; Ragnauth, C.D.; Yi, Q.; Mellad, J.A.; Warren, D.T.; et al. Nesprins-1 and -2 are involved in the pathogenesis of Emery-Dreifuss muscular dystrophy and are critical for nuclear envelope integrity. Hum. Mol. Genet. 2007, 16, 2816–2833. [Google Scholar] [CrossRef] [PubMed]
- Rahajro, W.H.; Enarson, P.; Sullivan, T.; Stewart, C.L.; Burke, B. Nuclear envelope defects associated with LMNA mutations cause dilated cardiomyopathy and Emery-Dreifuss muscular dystrophy. J. Cell Sci. 2001, 114, 4447–4457. [Google Scholar]
- Puckelwartz, M.J.; Kessler, E.J.; Kim, G.; deWitt, M.M.; Zhang, Y.; Earley, J.U.; Depreux, F.F.; Holaska, J.; Mewborn, S.K.; Pytel, P.; et al. Neprin-1 mutations in human and murine cardiomyopathy. J. Mol. Cell Cardiol. 2010, 48, 600–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Meinke, P.; Huomg, L.T.T.; Wehnert, M.; Noegel, A.A. Contribution of SUN1 mutations to the pathomechanism in muscular dystrophies. Hum. Mutat. 2014, 35, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Meinke, P.; Mattioli, E.; Haque, F.; Antoku, S.; Columbaro, M.; Straatman, K.R.; Worman, H.J.; Gundersen, G.G.; Lattanzi, G.; Wehnert, M.; et al. Muscular dystrophy associated SUN1 and SUN2 variants disrupt nuclear-cytoskeletal connections and myonuclear organization. PLoS Genet. 2014, 10, e1004114. [Google Scholar] [CrossRef] [Green Version]
- Razafsky, D.; Hodzic, D. Binding KASH under SUN: The many faces of nucleo-skeletal connections. J. Cell Biol. 2009, 186, 461–472. [Google Scholar] [CrossRef]
- Banerjee, I.; Zhang, J.; Moore-Morris, T.; Pfeiffer, E.; Buchholz, K.S.; Liu, A.; Ouyang, K.; Stroud, M.J.; Gerace, L.; Evans, S.M.; et al. Targeted ablation of nesprin 1 and nesprin 2 from murine myocardium results in cardiomyopathy, altered nuclear morphology and inhibition of biomechanical gene response. PLoS Genet. 2014, 10, e1004114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, C.L.; Burke, B. The missing LINC: A mammalian KASH protein coupling meiotic chromosomes to the cytoskeleton. Nucleus 5 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukášová, E.; Kovařík, A.; Bačíková, A.; Falk, M.; Kozubek, S. Loss of lamin B receptor is necessary to induce cellular senescence. Biochem. J. 2017, 474, 281–300. [Google Scholar] [CrossRef] [PubMed]
- Lukášová, E.; Řezáčová, M.; Bačíková, A.; Šebejová, L.; Vávrová, J.; Kozubek, S. Distinct cellular responses to replication stress leading to apoptosis or senescence. FEBS Open Biol. 2019, 9, 870–890. [Google Scholar] [CrossRef] [PubMed]
- Hirano, Y.; Hizume, K.; Kimura, H.; Takeyasu, K.; Haraguchi, T.; Hiraoka, Y. Lamin B receptor recognizes specific modifications of Histone 4 in heterochromatin formation. J. Biol. Chem. 2012, 287, 4265–42663. [Google Scholar] [CrossRef] [Green Version]
- Solovei, I.; Wang, A.S.; Thanisch, K.; Schmidt, C.S.; Krebs, S.; Zwerger, M.; Cohen, T.V.; Devys, D.; Foisner, R.; Peichl, L.; et al. LBR and lamin A/C sequentially tether peripheral heterochromatin and inversely regulate differentiation. Cell 2013, 152, 584–598. [Google Scholar] [CrossRef] [Green Version]
- Campisi, J.; d’Adda di Fagagna, F. Cellular senescence: When bad things happen to good cells. Nat. Rev. Mol. Cell Biol. 2007, 8, 729–740. [Google Scholar] [CrossRef]
- Fumagalli, M.; Rossiello, F.; Mondello, C.; d’Adda di Fagagna, F. Stable cellular senescence is associated with persistent DDR activation. PLoS ONE 2014. [Google Scholar] [CrossRef]
- Kosar, M.; Bartkova, J.; Hubackova, S.; Hodny, Z.; Lukas, J.; Bartek, J. Senescence-associated heterochromatin foci are dispensable for cellular senescence, occur in a cell type- and insult-dependent manner, and follow expression of p16ink4a. Cell Cycle 2011, 10, 457–468. [Google Scholar] [CrossRef] [Green Version]
- Matula, P.A.; Danek, O.; Maska, M.; Vinkler, M.; Kozubek, M. Acquiarium: Free software for acquisition and analysis of 3D images of cells Fluoresence Microscopy. In Proceedings of the IEEE International Symposium on Biomedical Imaging, Boston, MA, USA, 28 June–1 July 2009. [Google Scholar] [CrossRef]
- Dimri, G.P.; Lee, X.; Basile, G.; Acosta, M.; Scott, G.; Roskelley, C.; Medrano, E.E.; Linskens, M.; Rubelj, I.; Pereira-Smith, O. A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc. Natl. Acad. Sci. USA 1995, 92, 9363–9367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franek, E.; Haluzík, M.; Canecki-Varzic, S.; Sargin, M.; Macura, S.; Zacho, J.; Christiansen, J.S. Twice-daily insulin degludec/insulin as part provides superior fasting plasma glucose control and a reduced rate of hypoglycemia compared with basic insulin aspart 30 in insulin-naïve adults with Type 2 diabetes. Diabet. Med. 2016, 33, 497–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feringa, F.M.; Raajimakers, M.A.; Hadders, C.; Vaarting, L.; Macurek, L.; Heiting, L.; Krenning, L.; Medema, R.H. Persistent repair intermediates induce senescence. Nat. Com. 2018, 2923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stixová, L.; Komůrková, D.; Svobodová Kovaříková, A.; Bártová, E. UVA irradiation strengthened an interaction between UBF1/2 proteins and H4K20 di-/tri-methylation. Chromosome Res. 2019, 27, 41–55. [Google Scholar] [CrossRef]
- Mellad, J.A.; Warren, D.T.; Shanahan, C.M. Nesprins LINC the nucleus and cytoskeleton. Curr. Oppin. Cell Biol. 2011, 23, 47–54. [Google Scholar] [CrossRef]
- Lei, K.L.; Zhu, R.; Xu, C.; Shao, T.; Xu, Y.; Zhuang, Y.; Han, M. Inner nuclear envelope proteins SUN1 and SUN2 play a prominent role in the DNA damage response. Curr. Biol. 2012, 22, 1609–1615. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, R.K.; Rodriguez, E.C.; King, M.C. A role for nuclear envelope-bridging complexes in homology-directed repair. Mol. Biol. Cell 2014, 25, 2461–2471. [Google Scholar] [CrossRef]
- Sur, I.; Neumann, S.; Noegel, A.A. Nesprin-1 role in DNA damage response. Nucleus 2014, 5, 173–191. [Google Scholar] [CrossRef] [Green Version]
- Lottersberger, F.; Karssemeijer, R.A.; Dimitrova, N.; de Lange, E. 53BP1 and LINC complex promote microtubule-dependent DSB mobility and DNA repair. Cell 2015, 163, 880–893. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Zheng, H.; Wang, Y.; Lian, F.; Hu, Z.; Xue, S. Nesprin-1 plays an important role in the proliferation and apoptosis of mesenchymal stem cells. Int. J. Mol. Med. 2013, 32, 805–813. [Google Scholar] [CrossRef]
- Muchir, A.; van Engelen, B.G.; Lammens, M.; Mislow, J.M.; McNally, E.; Schwartz, K.; Bonne, G. Nuclear envelope alterations in fibroblasts from LGMD1B patients carrying nonsense Y259X heterozygous or homozygous mutation in lamin A/C gene. Exp. Cell Res. 2003, 291, 352–362. [Google Scholar] [CrossRef] [PubMed]
- Lammerding, J.; Hsiao, J.; Schulze, P.C.; Kozlov, S.; Stewart, C.L.; Lee, R.T. Abnormal nuclear shape and impaired mechanotransduction in emerin deficient cells. J. Cell Biol. 2005, 170, 781–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.; Li, C.; Zhou, B.; Sun, H.; Koullourou, V.; Holt, I.; Puckelwartz, M.J.; Warren, D.T.; Hayward, R.; Lin, Z.; et al. Novel nesprin-1 mutations associated with dilated cardiomyopathy cause nuclear envelope disruption and defects in myogenesis. Hum. Mol. Genet. 2017, 26, 2258–2276. [Google Scholar] [CrossRef] [PubMed]
- Ognibene, A.; Sabatelli, T.; Petrini, S.; Squarzoni, S.; Riccio, M.; Santi, S.; Villanova, M.; Palmeri, S.; Merlini, L.; Maraldi, M.N. Nuclear changes in a case of X-linked Emery-Dreifuss muscular dystrophy. Muscle Nerve 1999, 22, 864–869. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Svobodová Kovaříková, A.; Bártová, E.; Kovařík, A.; Lukášová, E. Spatiotemporal Mislocalization of Nuclear Membrane-Associated Proteins in γ-Irradiation-Induced Senescent Cells. Cells 2020, 9, 999. https://doi.org/10.3390/cells9040999
Svobodová Kovaříková A, Bártová E, Kovařík A, Lukášová E. Spatiotemporal Mislocalization of Nuclear Membrane-Associated Proteins in γ-Irradiation-Induced Senescent Cells. Cells. 2020; 9(4):999. https://doi.org/10.3390/cells9040999
Chicago/Turabian StyleSvobodová Kovaříková, Alena, Eva Bártová, Aleš Kovařík, and Emilie Lukášová. 2020. "Spatiotemporal Mislocalization of Nuclear Membrane-Associated Proteins in γ-Irradiation-Induced Senescent Cells" Cells 9, no. 4: 999. https://doi.org/10.3390/cells9040999
APA StyleSvobodová Kovaříková, A., Bártová, E., Kovařík, A., & Lukášová, E. (2020). Spatiotemporal Mislocalization of Nuclear Membrane-Associated Proteins in γ-Irradiation-Induced Senescent Cells. Cells, 9(4), 999. https://doi.org/10.3390/cells9040999