Sildenafil Recovers Burn-Induced Cardiomyopathy



Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals and Burn Model
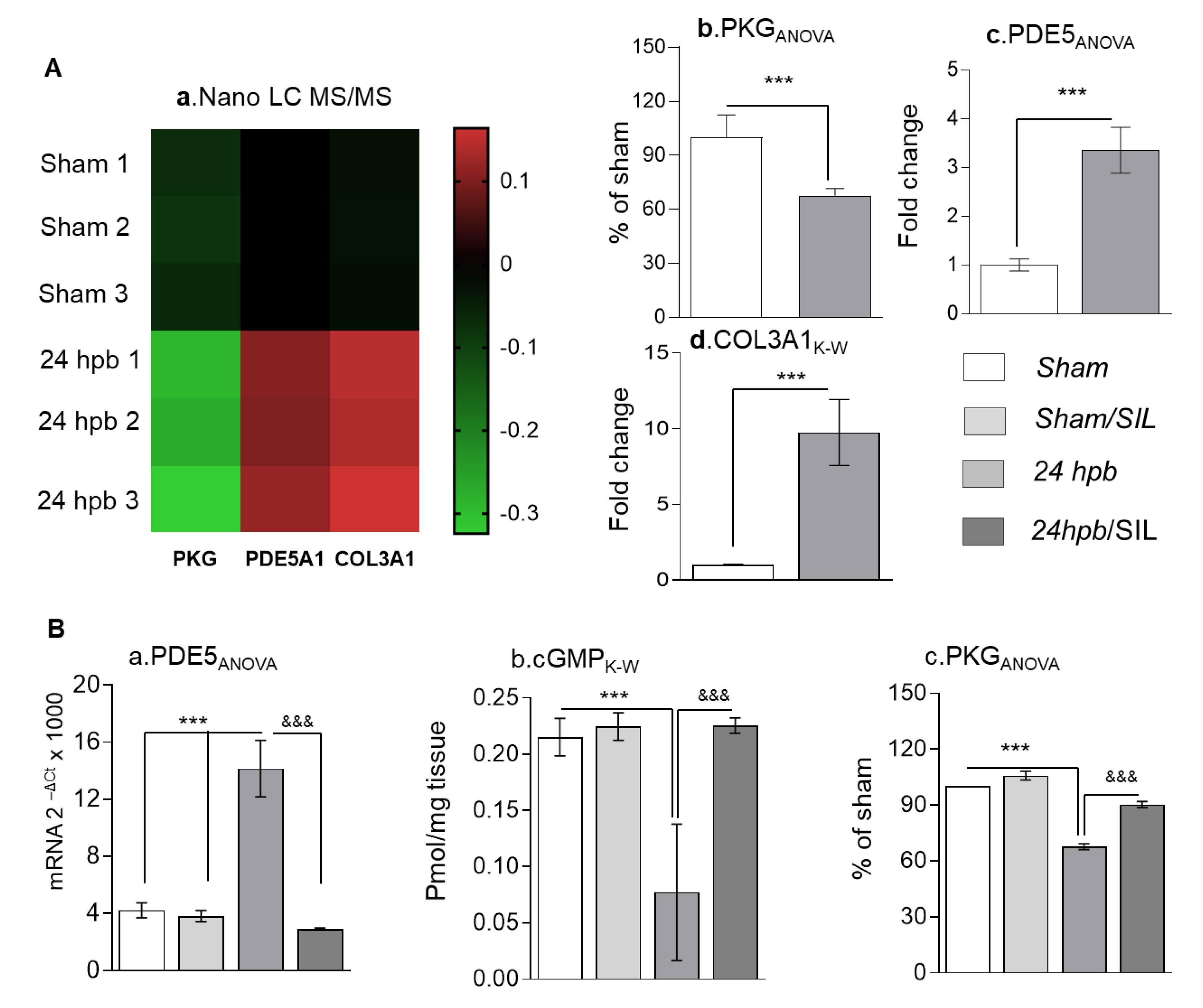
2.3. Nano LC-MS/MS
2.4. cGMP Level
2.5. The cGMP Dependent Protein Kinase (PKG) Activity
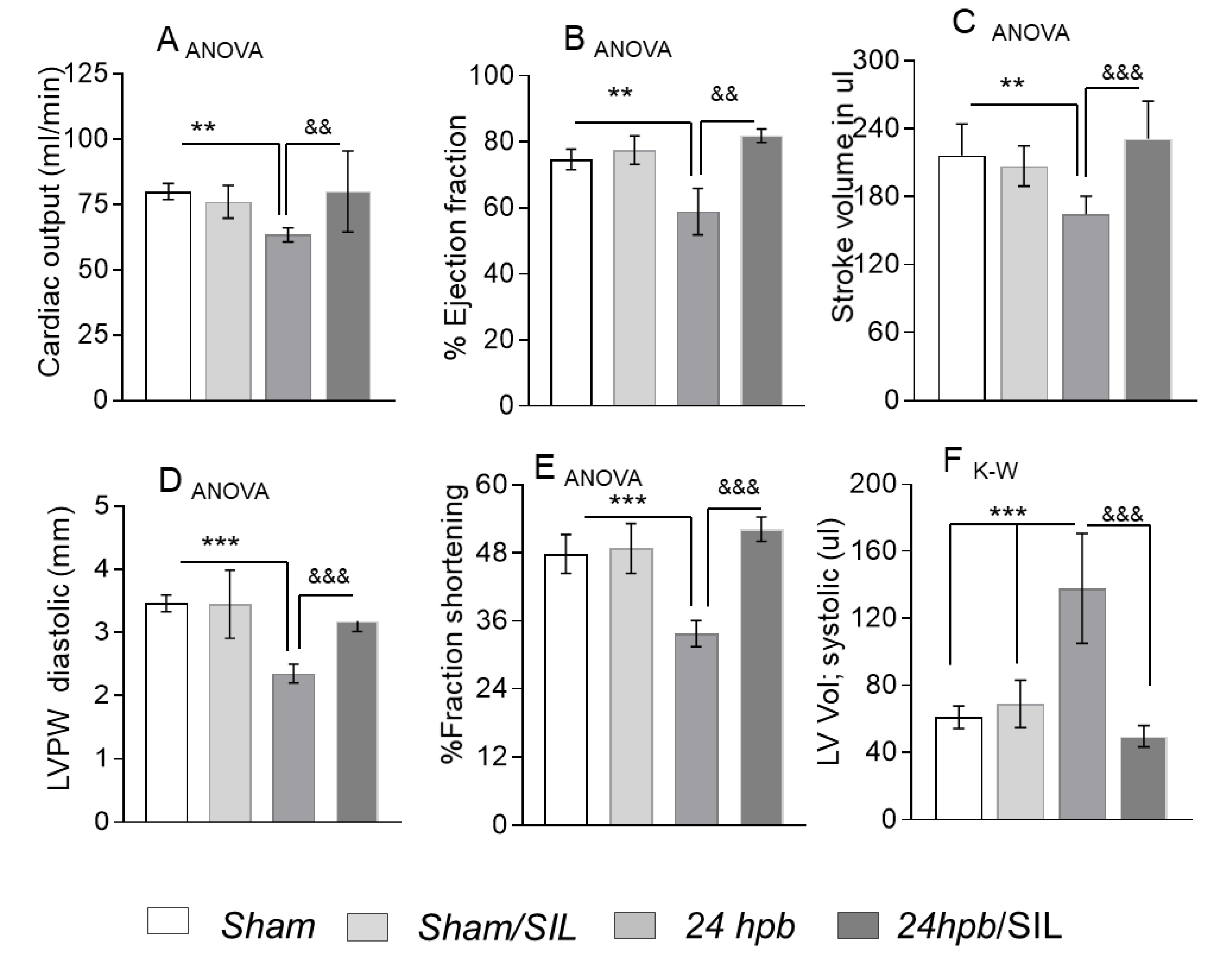
2.6. Echocardiography (ECHO)
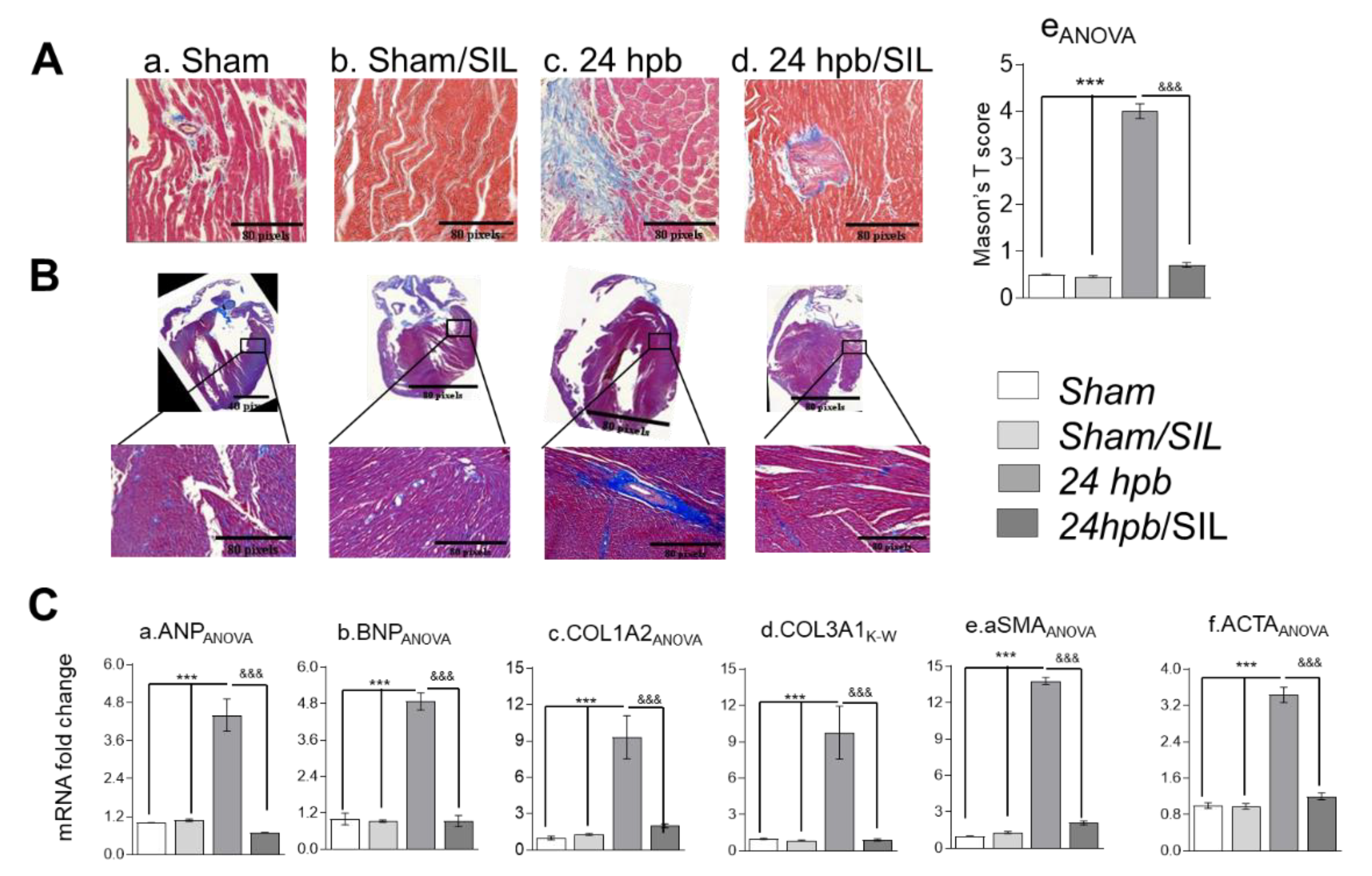
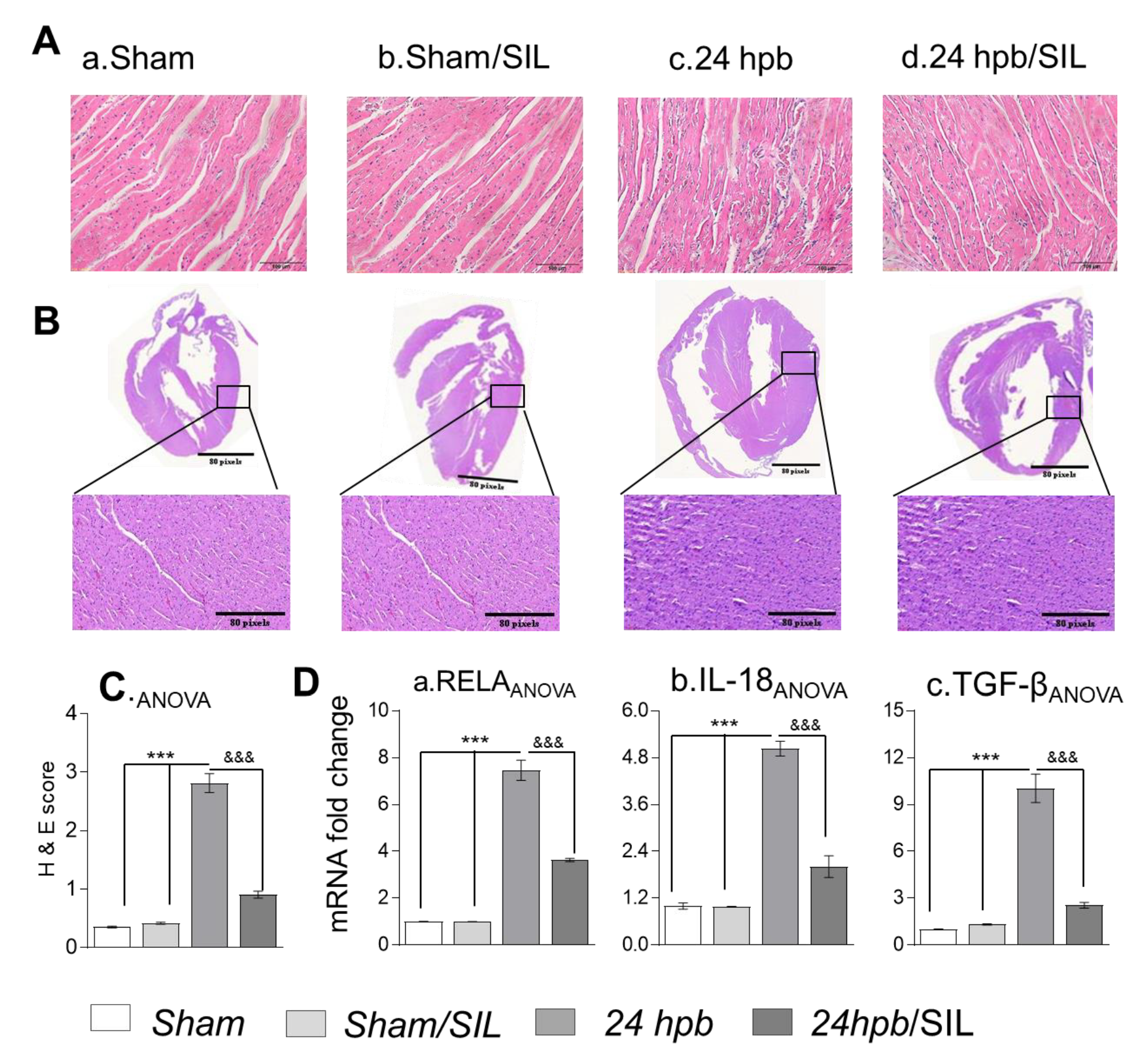
2.7. Histology
2.8. Gene Expression Analysis
2.9. Oxidative Stress
2.10. Antioxidant Levels
2.11. ATP Colorimetric Assay
2.12. Statistical Analysis
3. Results
3.1. The Importance of the PDE5A-cGMP-PKG Pathway in Cardiomyocytes after a Burn
3.2. Effect of PDE5A Inhibition on Burn-Induced Cardiac Dysfunction
3.3. Effect of PDE5 Inhibition on Burn-Induced Cardiac Fibrogenesis
3.4. Effect of PDE5A Inhibition on Myocardial Inflammation after a Burn
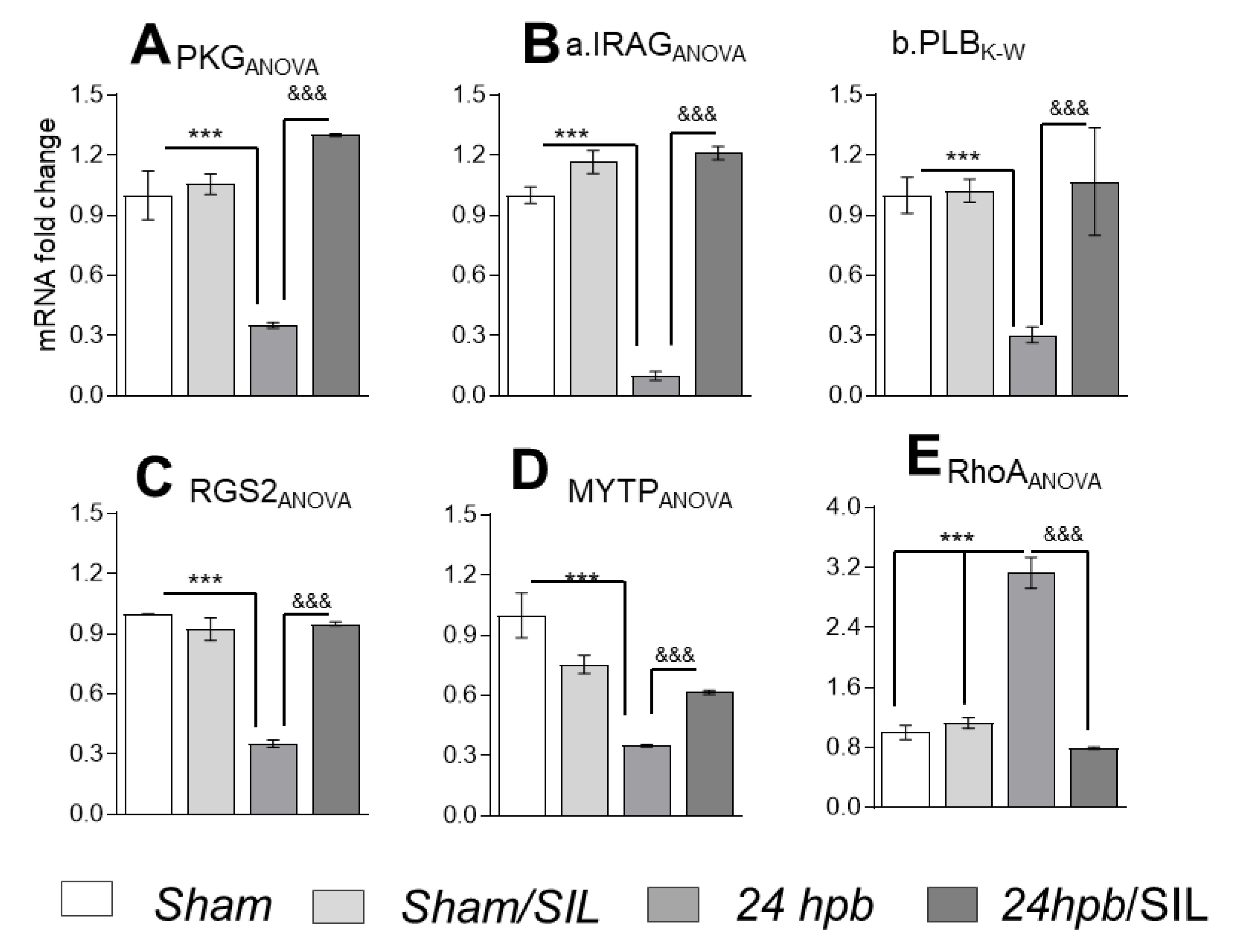
3.5. The Effect of PDE5A Inhibition on Downstream Gene Expression after a Burn
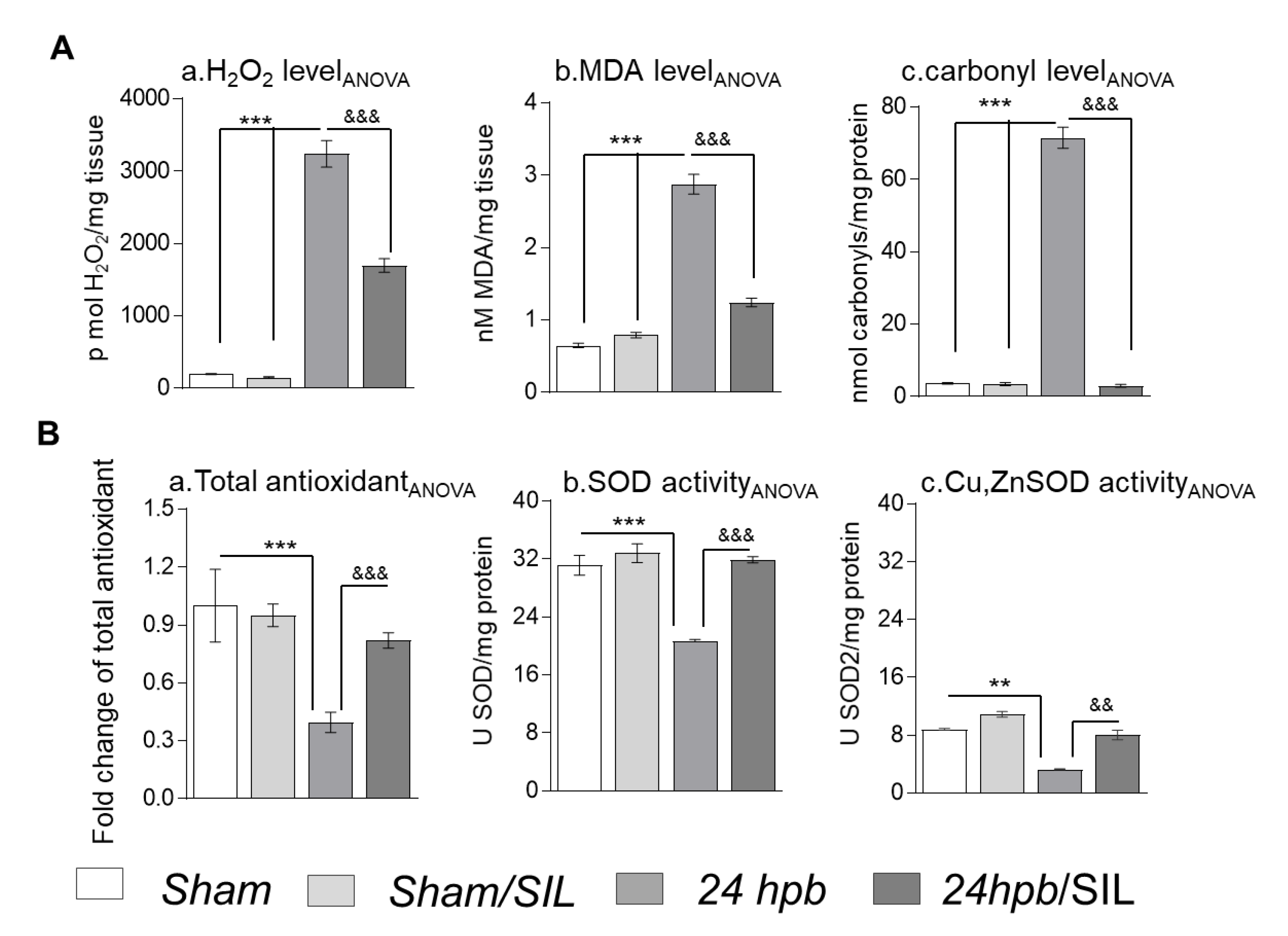
3.6. The Effect of PDE5A Inhibition on Oxidant/Antioxidant Imbalance
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. Burns. Available online: https://www.who.int/violence_injury_prevention/other_injury/burns/en/ (accessed on 2 June 2020).
- CDC. Burn Prevention. Available online: https://www.cdc.gov/safechild/burns/index.html (accessed on 2 June 2020).
- Colohan, S.M. Predicting prognosis in thermal burns with associated inhalational injury: A systematic review of prognostic factors in adult burn victims. J. Burn Care Res. 2010, 31, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, B.A.; Zaloshnja, E.; Miller, T.R.; Jones, P.R. Estimates of the Incidence and Costs of Fire-Related Injuries; The U.S. Consumer Product Safety Commission: Calveston, MD, USA, 2009; pp. 1–51.
- Hoesel, L.M.; Niederbichler, A.D.; Schaefer, J.; Ipaktchi, K.R.; Gao, H.; Rittirsch, D.; Pianko, M.J.; Vogt, P.M.; Sarma, J.V.; Su, G.L.; et al. C5a-blockade improves burn-induced cardiac dysfunction. J. Immunol. 2007, 178, 7902–7910. [Google Scholar] [CrossRef]
- Tsai, E.J.; Kass, D.A. Cyclic GMP signaling in cardiovascular pathophysiology and therapeutics. Pharmacol. Ther. 2009, 122, 216–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Layland, J.; Li, J.M.; Shah, A.M. Role of cyclic GMP-dependent protein kinase in the contractile response to exogenous nitric oxide in rat cardiac myocytes. J. Physiol. 2002, 540, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Kass, D.A. Phosphodiesterases and cardiac cGMP: Evolving roles and controversies. Trends Pharmacol. Sci. 2011, 32, 360–365. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.I.; Kass, D.A. Phosphodiesterases and cyclic GMP regulation in heart muscle. Physiology (Bethesda) 2012, 27, 248–258. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.E.; Kass, D.A. Cardiac Phosphodiesterases and Their Modulation for Treating Heart Disease. Handb. Exp. Pharmacol. 2017, 243, 249–269. [Google Scholar] [CrossRef] [Green Version]
- Nagayama, T.; Zhang, M.; Hsu, S.; Takimoto, E.; Kass, D.A. Sustained soluble guanylate cyclase stimulation offsets nitric-oxide synthase inhibition to restore acute cardiac modulation by sildenafil. J. Pharmacol. Exp. Ther. 2008, 326, 380–387. [Google Scholar] [CrossRef] [Green Version]
- Andersson, K.E. PDE5 inhibitors-pharmacology and clinical applications 20 years after sildenafil discovery. Br. J. Pharmacol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Wen, J.J.; Cummins, C.B.; Szczesny, B.; Radhakrishnan, R.S. Cardiac Dysfunction after Burn Injury: Role of the AMPK-SIRT1-PGC1alpha-NFE2L2-ARE Pathway. J. Am. Coll. Surg. 2020. [Google Scholar] [CrossRef]
- Wen, J.J.; Cummins, C.B.; Radhakrishnan, R.S. Burn-Induced Cardiac Mitochondrial Dysfunction via Interruption of the PDE5A-cGMP-PKG Pathway. Int. J. Mol. Sci. 2020, 21, 2350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mascarenhas, D.D.; Elayadi, A.; Singh, B.K.; Prasai, A.; Hegde, S.D.; Herndon, D.N.; Finnerty, C.C. Nephrilin peptide modulates a neuroimmune stress response in rodent models of burn trauma and sepsis. Int. J. Burns Trauma 2013, 3, 190–200. [Google Scholar] [PubMed]
- Bohanon, F.J.; Nunez Lopez, O.; Herndon, D.N.; Wang, X.; Bhattarai, N.; Ayadi, A.E.; Prasai, A.; Jay, J.W.; Rojas-Khalil, Y.; Toliver-Kinsky, T.E.; et al. Burn Trauma Acutely Increases the Respiratory Capacity and Function of Liver Mitochondria. Shock 2018, 49, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Bhan, A.; Sirker, A.; Zhang, J.; Protti, A.; Catibog, N.; Driver, W.; Botnar, R.; Monaghan, M.J.; Shah, A.M. High-frequency speckle tracking echocardiography in the assessment of left ventricular function and remodeling after murine myocardial infarction. Am. J. Physiol. Heart Circ. Physiol. 2014, 306, H1371–H1383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, J.J.; Porter, C.; Garg, N.J. Inhibition of NFE2L2-Antioxidant Response Element Pathway by Mitochondrial Reactive Oxygen Species Contributes to Development of Cardiomyopathy and Left Ventricular Dysfunction in Chagas Disease. Antioxid. Redox Signal. 2017, 27, 550–566. [Google Scholar] [CrossRef]
- Wen, J.J.; Garg, N.J. Manganese superoxide dismutase deficiency exacerbates the mitochondrial ROS production and oxidative damage in Chagas disease. PLoS Negl. Trop. Dis. 2018, 12, e0006687. [Google Scholar] [CrossRef]
- Hernandez, P.; Buller, D.; Mitchell, T.; Wright, J.; Liang, H.; Manchanda, K.; Welch, T.; Huebinger, R.M.; Carlson, D.L.; Wolf, S.E.; et al. Severe Burn-Induced Inflammation and Remodeling of Achilles Tendon in a Rat Model. Shock 2018, 50, 346–350. [Google Scholar] [CrossRef]
- Nakamura, T.; Ranek, M.J.; Lee, D.I.; Shalkey Hahn, V.; Kim, C.; Eaton, P.; Kass, D.A. Prevention of PKG1alpha oxidation augments cardioprotection in the stressed heart. J. Clin. Investig. 2015, 125, 2468–2472. [Google Scholar] [CrossRef] [Green Version]
- Takimoto, E. Cyclic GMP-dependent signaling in cardiac myocytes. Circ. J. 2012, 76, 1819–1825. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Osanai, T.; Nakano, T.; Wakui, M.; Okumura, K. Enhanced activities and gene expression of phosphodiesterase types 3 and 4 in pressure-induced congestive heart failure. Heart Vessels 2002, 16, 249–256. [Google Scholar] [CrossRef]
- West, T.M.; Wang, Q.; Deng, B.; Zhang, Y.; Barbagallo, F.; Reddy, G.R.; Chen, D.; Phan, K.S.; Xu, B.; Isidori, A.; et al. Phosphodiesterase 5 Associates With beta2 Adrenergic Receptor to Modulate Cardiac Function in Type 2 Diabetic Hearts. J. Am. Heart Assoc. 2019, 8, e012273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, J.J.; Wan, X.; Thacker, J.; Garg, N.J. Chemotherapeutic efficacy of phosphodiesterase inhibitors in chagasic cardiomyopathy. JACC Basic Transl. Sci. 2016, 1, 235–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutchings, D.C.; Anderson, S.G.; Caldwell, J.L.; Trafford, A.W. Phosphodiesterase-5 inhibitors and the heart: Compound cardioprotection? Heart 2018, 104, 1244–1250. [Google Scholar] [CrossRef]
- Barnes, H.; Brown, Z.; Burns, A.; Williams, T. Phosphodiesterase 5 inhibitors for pulmonary hypertension. Cochrane Database Syst. Rev. 2019, 1, CD012621. [Google Scholar] [CrossRef]
- Guillory, A.N.; Clayton, R.P.; Herndon, D.N.; Finnerty, C.C. Cardiovascular Dysfunction Following Burn Injury: What We Have Learned from Rat and Mouse Models. Int. J. Mol. Sci. 2016, 17, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klinger, J.R.; Kadowitz, P.J. The Nitric Oxide Pathway in Pulmonary Vascular Disease. Am. J. Cardiol. 2017, 120, S71–S79. [Google Scholar] [CrossRef] [Green Version]
- Kass, D.A.; Champion, H.C.; Beavo, J.A. Phosphodiesterase type 5: Expanding roles in cardiovascular regulation. Circ. Res. 2007, 101, 1084–1095. [Google Scholar] [CrossRef]
- Gao, J.; Xu, Y.; Lei, M.; Shi, J.; Gong, Q. Icariside II, a PDE5 inhibitor from Epimedium brevicornum, promotes neuron-like pheochromocytoma PC12 cell proliferation via activating NO/cGMP/PKG pathway. Neurochem. Int. 2018, 112, 18–26. [Google Scholar] [CrossRef]
- Hundeshagen, G.; Herndon, D.N.; Clayton, R.P.; Wurzer, P.; McQuitty, A.; Jennings, K.; Branski, L.; Collins, V.N.; Marques, N.R.; Finnerty, C.C.; et al. Long-term effect of critical illness after severe paediatric burn injury on cardiac function in adolescent survivors: An observational study. Lancet Child Adolesc. Health 2017, 1, 293–301. [Google Scholar] [CrossRef]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb Perspect Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [Green Version]
- Sekiyama, A.; Ueda, H.; Kashiwamura, S.; Sekiyama, R.; Takeda, M.; Rokutan, K.; Okamura, H. A stress-induced, superoxide-mediated caspase-1 activation pathway causes plasma IL-18 upregulation. Immunity 2005, 22, 669–677. [Google Scholar] [CrossRef] [Green Version]
- Vidal-Vanaclocha, F.; Fantuzzi, G.; Mendoza, L.; Fuentes, A.M.; Anasagasti, M.J.; Martin, J.; Carrascal, T.; Walsh, P.; Reznikov, L.L.; Kim, S.H.; et al. IL-18 regulates IL-1beta-dependent hepatic melanoma metastasis via vascular cell adhesion molecule-1. Proc. Natl. Acad. Sci. USA 2000, 97, 734–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Darawish, Y.; Li, W.; Yamanishi, K.; Pencheva, M.; Oka, N.; Yamanishi, H.; Matsuyama, T.; Tanaka, Y.; Minato, N.; Okamura, H. Frontline Science: IL-18 primes murine NK cells for proliferation by promoting protein synthesis, survival, and autophagy. J. Leukoc. Biol. 2018, 104, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Pu, Z.; Che, Y.; Zhang, W.; Sun, H.; Meng, T.; Xie, H.; Cao, L.; Hao, H. Dual roles of IL-18 in colitis through regulation of the function and quantity of goblet cells. Int. J. Mol. Med. 2019, 43, 2291–2302. [Google Scholar] [CrossRef]
- Dinarello, C.A.; Novick, D.; Kim, S.; Kaplanski, G. Interleukin-18 and IL-18 binding protein. Front. Immunol. 2013, 4, 289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobaczewski, M.; Chen, W.; Frangogiannis, N.G. Transforming growth factor (TGF)-beta signaling in cardiac remodeling. J. Mol. Cell. Cardiol. 2011, 51, 600–606. [Google Scholar] [CrossRef] [Green Version]
- Klass, B.R.; Grobbelaar, A.O.; Rolfe, K.J. Transforming growth factor beta1 signalling, wound healing and repair: A multifunctional cytokine with clinical implications for wound repair, a delicate balance. Postgrad. Med. J. 2009, 85, 9–14. [Google Scholar] [CrossRef]
- Hofmann, F.; Feil, R.; Kleppisch, T.; Schlossmann, J. Function of cGMP-dependent protein kinases as revealed by gene deletion. Physiol. Rev. 2006, 86, 1–23. [Google Scholar] [CrossRef]
- Lu, Z.; Xu, X.; Hu, X.; Lee, S.; Traverse, J.H.; Zhu, G.; Fassett, J.; Tao, Y.; Zhang, P.; dos Remedios, C.; et al. Oxidative stress regulates left ventricular PDE5 expression in the failing heart. Circulation 2010, 121, 1474–1483. [Google Scholar] [CrossRef] [Green Version]
- Koupparis, A.J.; Jeremy, J.Y.; Muzaffar, S.; Persad, R.; Shukla, N. Sildenafil inhibits the formation of superoxide and the expression of gp47 NAD[P]H oxidase induced by the thromboxane A2 mimetic, U46619, in corpus cavernosal smooth muscle cells. BJU Int. 2005, 96, 423–427. [Google Scholar] [CrossRef]
- Borlaug, B.A.; Melenovsky, V.; Marhin, T.; Fitzgerald, P.; Kass, D.A. Sildenafil inhibits beta-adrenergic-stimulated cardiac contractility in humans. Circulation 2005, 112, 2642–2649. [Google Scholar] [CrossRef] [PubMed]
- Lawless, M.; Caldwell, J.L.; Radcliffe, E.J.; Smith, C.E.R.; Madders, G.W.P.; Hutchings, D.C.; Woods, L.S.; Church, S.J.; Unwin, R.D.; Kirkwood, G.J.; et al. Phosphodiesterase 5 inhibition improves contractile function and restores transverse tubule loss and catecholamine responsiveness in heart failure. Sci. Rep. 2019, 9, 6801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Ayadi, A.; Prasai, A.; Wang, Y.; Herndon, D.N.; Finnerty, C.C. beta-Adrenergic Receptor Trafficking, Degradation, and Cell Surface Expression Are Altered in Dermal Fibroblasts from Hypertrophic Scars. J. Investig. Dermatol. 2018, 138, 1645–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivas, E.; Herndon, D.N.; Cambiaso-Daniel, J.; Rontoyanni, V.G.; Porter, C.; Glover, S.; Suman, O.E. Quantification of an Exercise Rehabilitation Program for Severely Burned Children: The Standard of Care at Shriners Hospitals for Children(R)-Galveston. J. Burn Care Res. 2018, 39, 889–896. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | 5′-Forward-3′ | 5′-Reverse-3′ | Ampliconsize (bp) | Accession # |
|---|---|---|---|---|
| α-tubulin | CAATTCCATCCTCACCACCCA | TCAACCTGTTTAAGTTAGTGTAGGT | 137 | AH002269.2 |
| β-actin | ACTGGCATTGTGATGGACTC | GCTCGGTCAGGATCTTCATG | 142 | V01217.1 |
| ACTA | AGGGAGTGATGGTTGGAATG | GGTGATGATGCCGTGTTCTA | 110 | BC158550.1 |
| ANP | CGAGAGTCAGGAAACGGAAAG | CTCAGACACACACACACATACA | 99 | M60731.1 |
| BNP | CACCTCTCAAGTGATCCTGTTT | GCAAGTTTGTGCTGGAAGATAAG | 99 | M25297.1 |
| COLIA2 | CCAGAGTGGAAGAGCGATTAC | ATGCAGGTTTCACCAGTAGAG | 101 | NM_053356.1 |
| COLIIIA1 | CAGGCCAATGGCAATGTAAAG | GCCATCCTCTAGAACTGTGTAAG | 108 | BC087039.1 |
| GAPDH | ACTCCCATTCTTCCACCTTTG | CCCTGTTGCTGTAGCCATATT | 105 | NM_017008.4 |
| IL-18 | GAATCCCAGACCAGACTGATAAT | GGTAGACATCCTTCCATCCTTC | 96 | NM_019165.1 |
| IRAG | GTACAACTGTCCTTGGCCTTTA | GTTCCTTCTCGGTGTTCTCTTC | 106 | NM_001105211.1 |
| MYTP | CCTTTCCAGCACAAGCACTA | GTACTATCCTCAGCCCACAAAC | 99 | AF110176.1 |
| PDE5 | GCCGCCACTATTATCTCCTTC | CTACTTCCTCCCACTCCATTTG | 112 | NM_133584.1 |
| PKG | GGGAAGGTCGAAGTCACAAA | CTGTCCGGGTACAGTTGTAAAG | 100 | EU251189.1 |
| PLB | TATCAGGAGAGCCTCGACTATT | CAGATCAGCAGCAGACATATCA | 105 | NM_022707.2 |
| RELA | GCTCAAGATCTGCCGAGTAAA | GTCCCGTGAAATACACCTCAA | 113 | AY307375.1 |
| RGS2 | GGAAGACCCGTTTGAGCTATT | TCCTCAGGAGAAGGCTTGATA | 106 | NM_053453.2 |
| RhoA | GACCAGTTCCCAGAGGTTTATG | GTCCCATAAAGCCAACTCTACC | 96 | D84477.1 |
| αSMA | CACCGCTGAACGTGAAATTG | CTTCTCCAGAGAGGAGGAAGAT | 109 | NM_019183.1 |
| TGF-β | CTGAACCAAGGAGACGGAATAC | GTTTGGGACTGATCCCATTGA | 106 | AY550025.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, J.J.; Cummins, C.; Radhakrishnan, R.S. Sildenafil Recovers Burn-Induced Cardiomyopathy. Cells 2020, 9, 1393. https://doi.org/10.3390/cells9061393
Wen JJ, Cummins C, Radhakrishnan RS. Sildenafil Recovers Burn-Induced Cardiomyopathy. Cells. 2020; 9(6):1393. https://doi.org/10.3390/cells9061393
Chicago/Turabian StyleWen, Jake J., Claire Cummins, and Ravi S. Radhakrishnan. 2020. "Sildenafil Recovers Burn-Induced Cardiomyopathy" Cells 9, no. 6: 1393. https://doi.org/10.3390/cells9061393
APA StyleWen, J. J., Cummins, C., & Radhakrishnan, R. S. (2020). Sildenafil Recovers Burn-Induced Cardiomyopathy. Cells, 9(6), 1393. https://doi.org/10.3390/cells9061393