Inhibition of ABCC6 Transporter Modifies Cytoskeleton and Reduces Motility of HepG2 Cells via Purinergic Pathway

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatments
2.2. Generation of Stable Abcc6 Knockdown HepG2 Cells
2.3. Real-Time Reverse Transcription Polymerase Chain Reaction (RT-qPCR)
2.4. Western Blot Analysis
2.5. Extracellular ATP Bioluminescence Assay
2.6. Cell Cycle Analysis
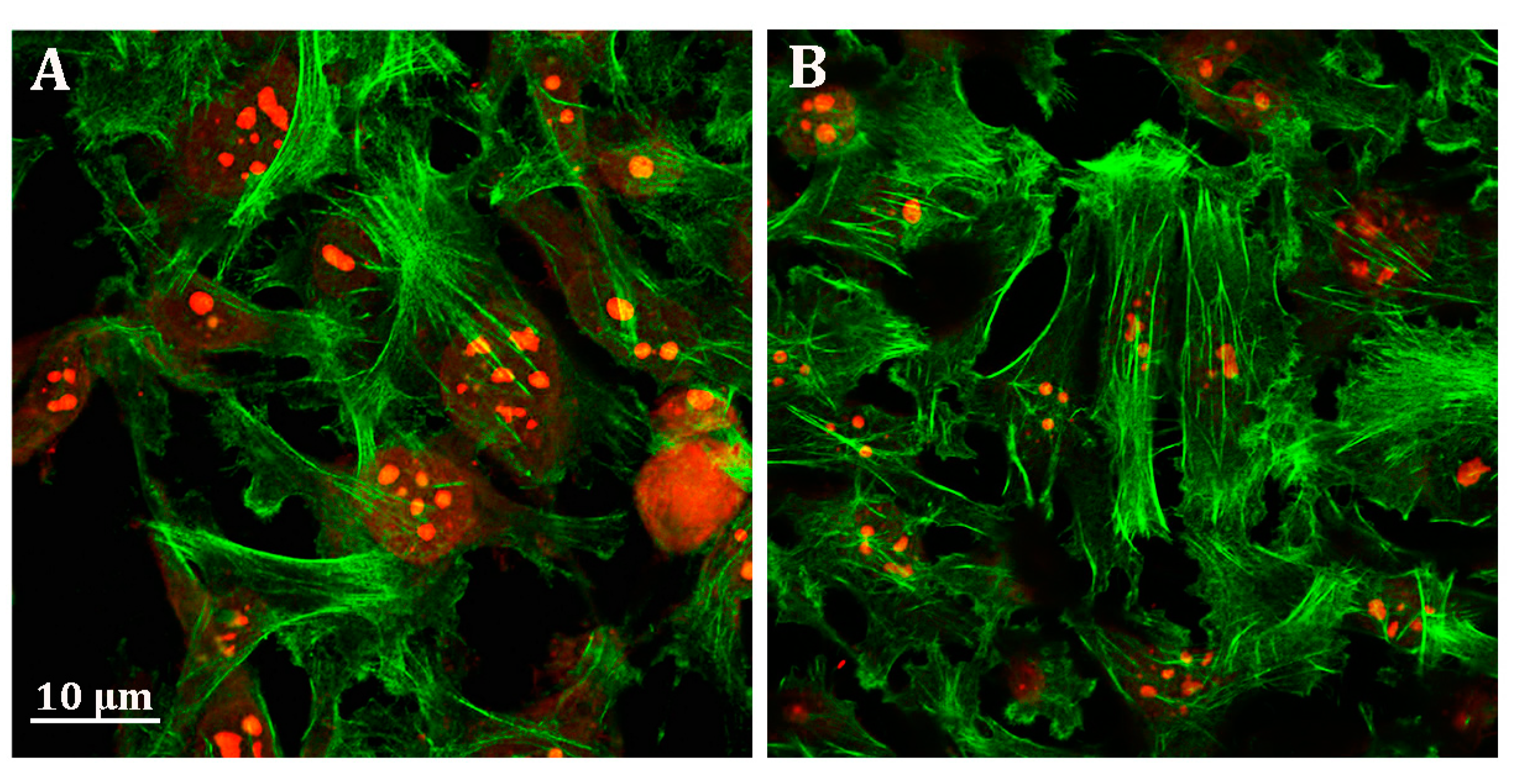
2.7. Confocal Fluorescence Microscopy
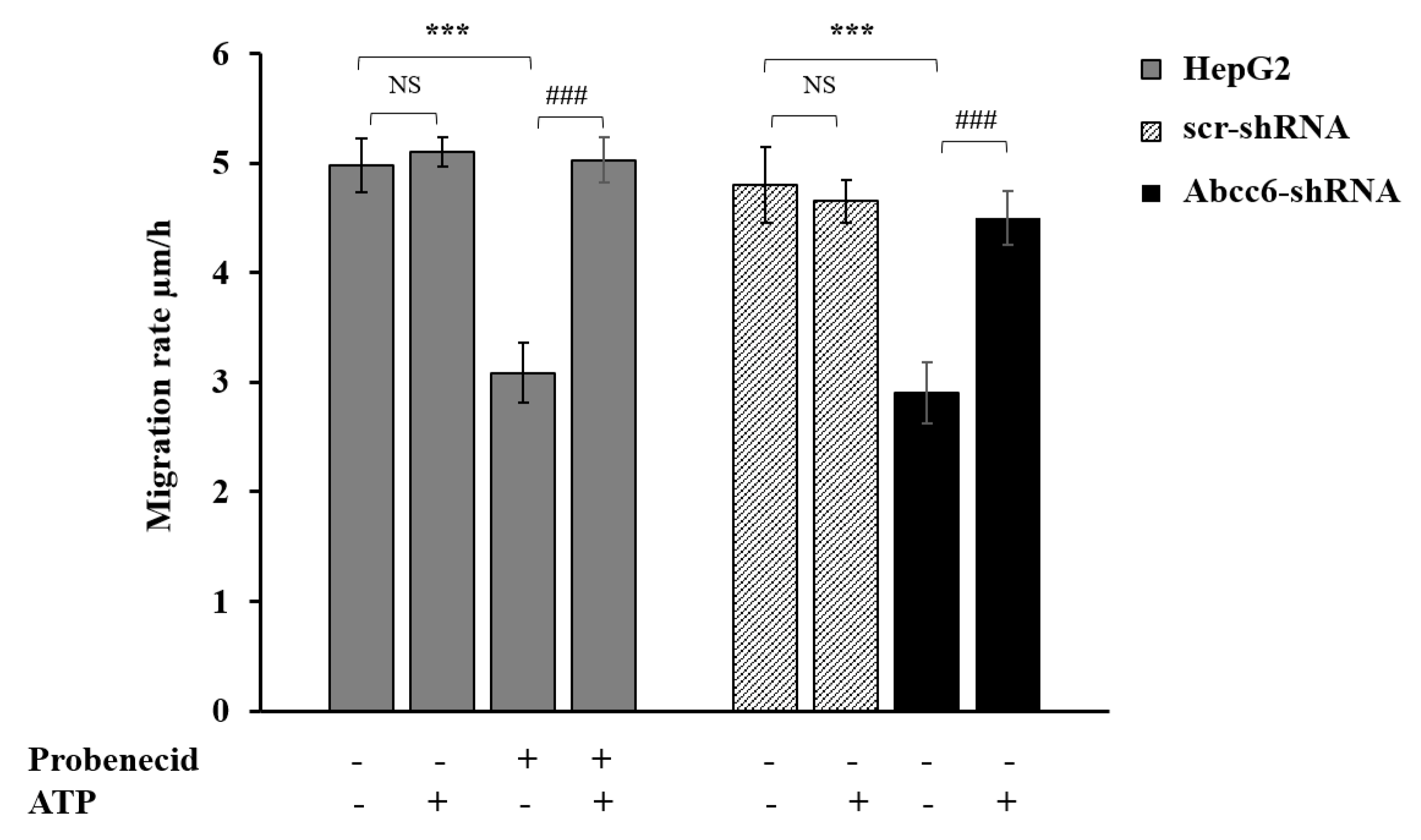
2.8. Migration Assay
2.9. Statistical Analysis
3. Results
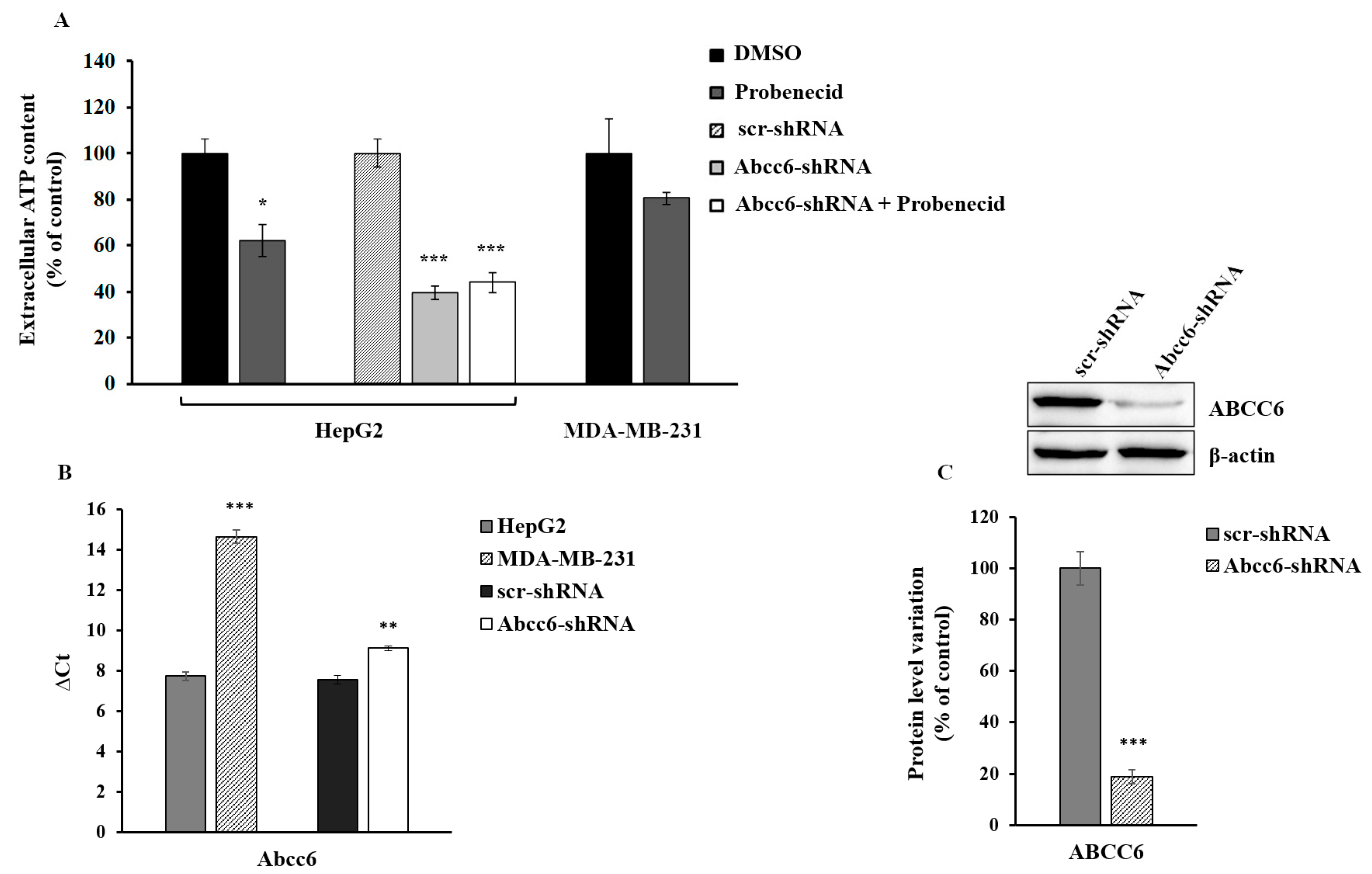
3.1. Probenecid Affects the Extracellular ATP Released by HepG2 Cells
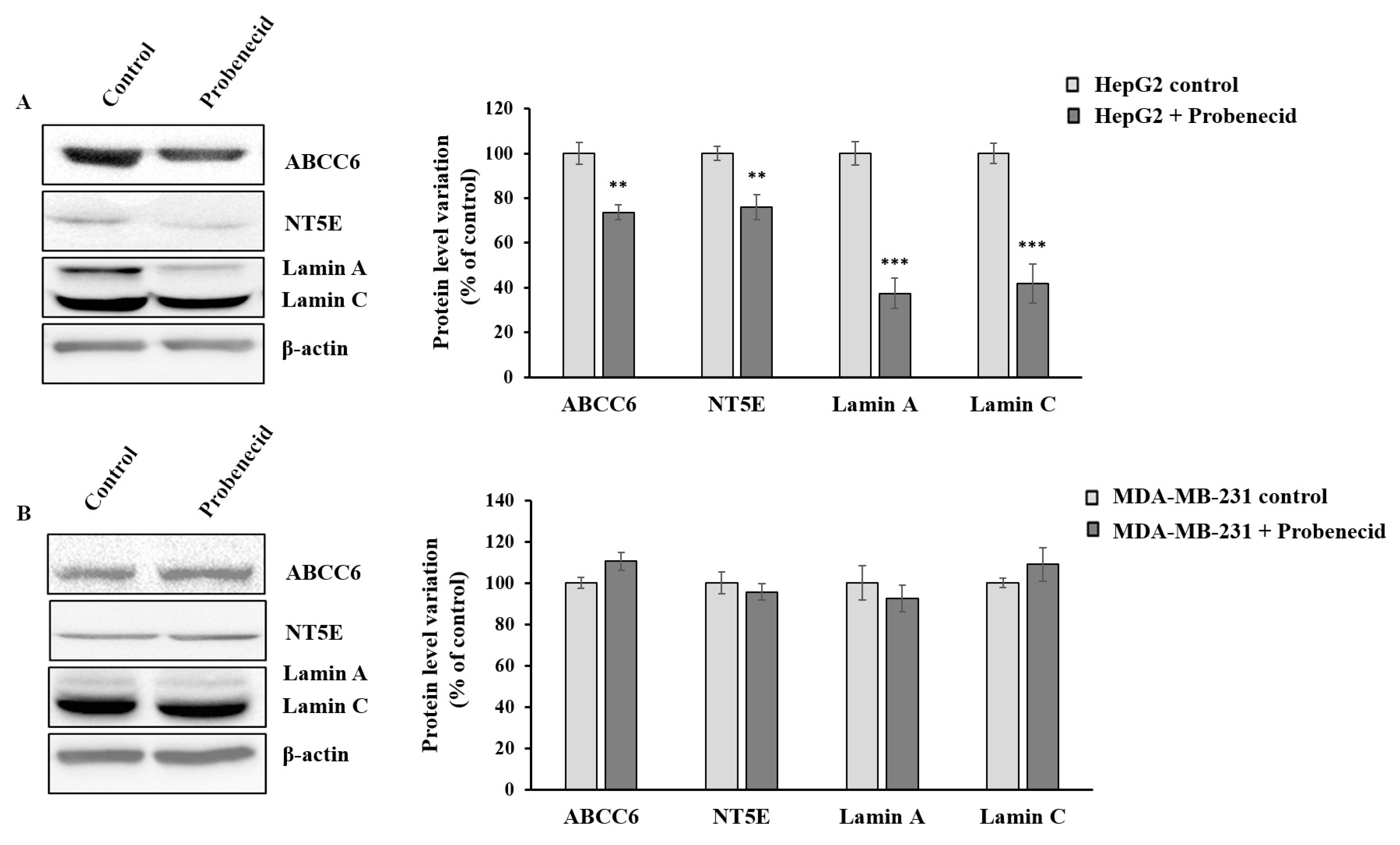
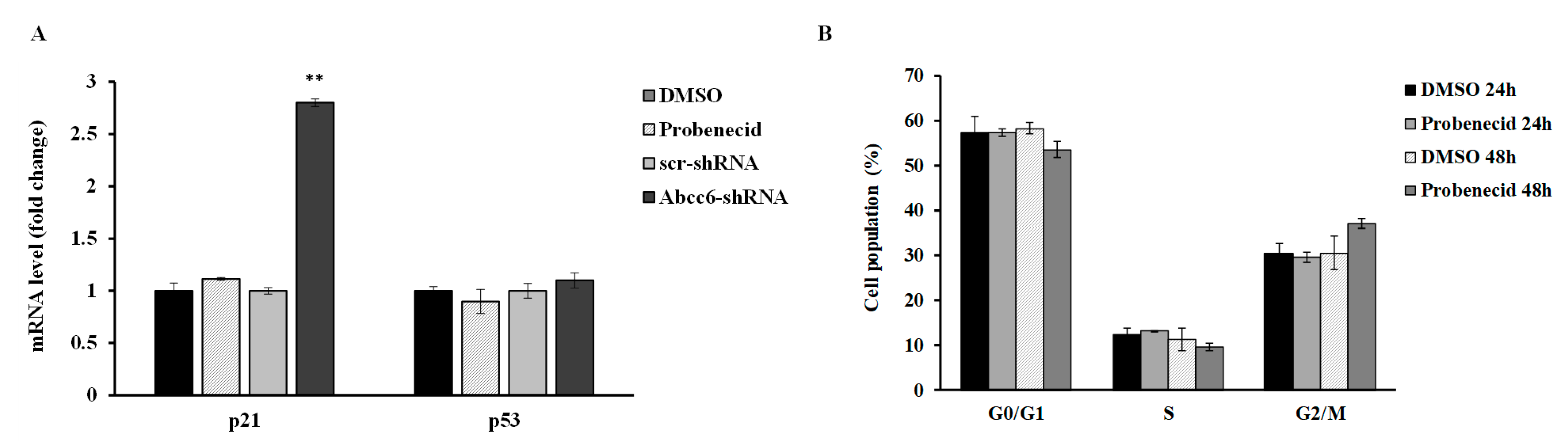
3.2. Probenecid Affects the Expression of Some Genes in HepG2 Cells
3.3. Probenecid Does Not Affect HepG2 Cell Cycle
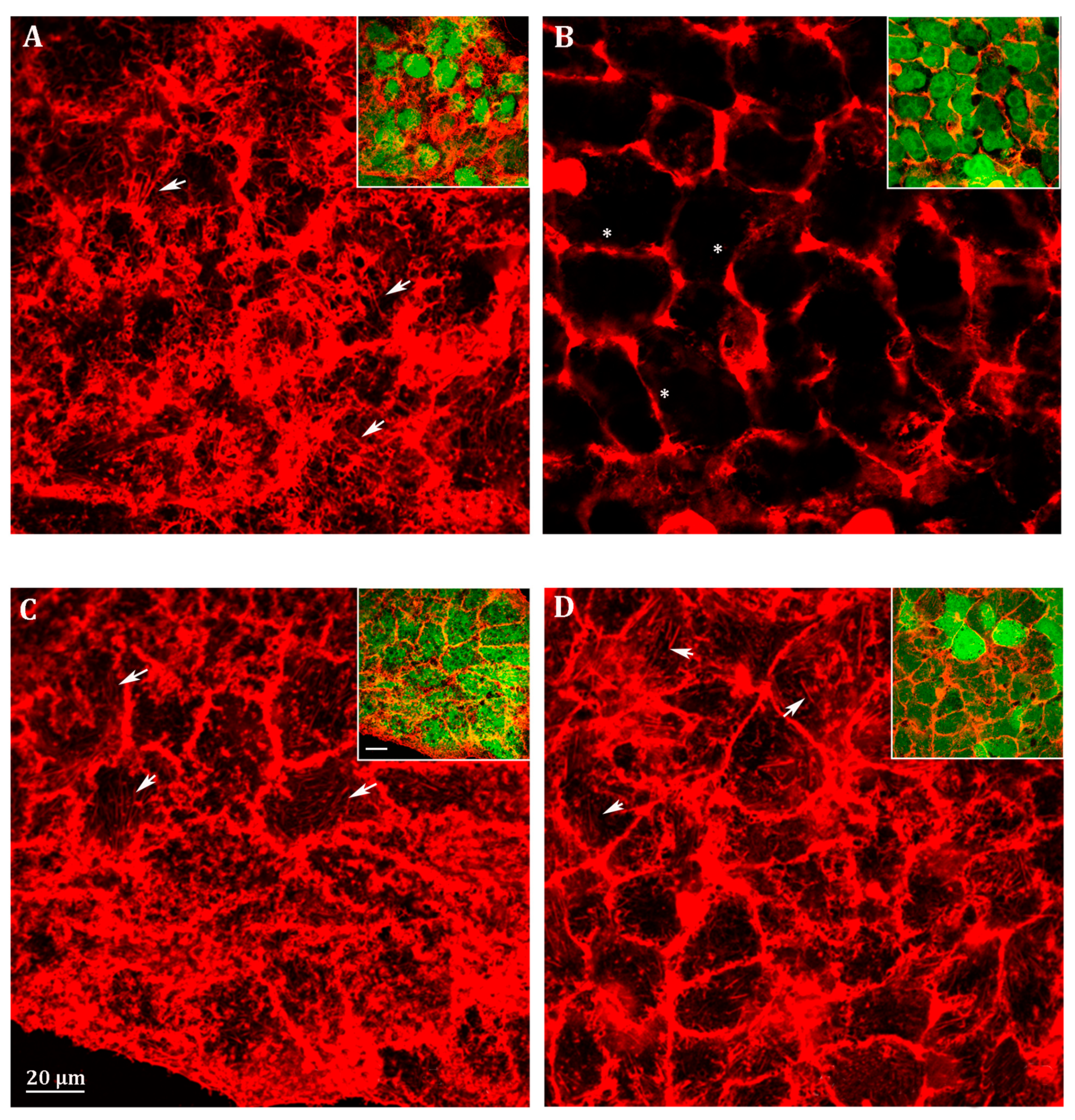
3.4. Probenecid Induces Cytoskeletal Rearrangement of HepG2 Cells
3.5. Probenecid Affects HepG2 Cell Migration
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Giuliani, A.L.; Sarti, A.C.; Di Virgilio, F. Extracellular Nucleotides and Nucleosides as Signalling Molecules. Immunol. Lett. 2019, 205, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Dosch, M.; Gerber, J.; Jebbawi, F.; Beldi, G. Mechanisms of ATP Release by Inflammatory Cells. Int. J. Mol. Sci. 2018, 19, 1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taruno, A. ATP Release Channels. Int. J. Mol. Sci. 2018, 19, 808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pomozi, V.; Le Saux, O.; Brampton, C.; Apana, A.; Iliás, A.; Szeri, F.; Martin, L.; Monostory, K.; Paku, S.; Sarkadi, B.; et al. ABCC6 is a Basolateral Plasma Membrane Protein. Circ. Res. 2013, 112, e148–e151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Favre, G.; Laurain, A.; Aranyi, T.; Szeri, F.; Fulop, K.; Le Saux, O.; Duranton, C.; Kauffenstein, G.; Martin, L.; Lefthériotis, G. The ABCC6 Transporter: A New Player in Biomineralization. Int. J. Mol. Sci. 2017, 18, 1941. [Google Scholar] [CrossRef]
- Borst, P.; Váradi, A.; van de Wetering, K. PXE, a Mysterious Inborn Error Clarified. Trends Biochem. Sci. 2019, 44, 125–140. [Google Scholar] [CrossRef]
- Sánchez-Tévar, A.M.; García-Fernández, M.; Murcia-Casas, B.; Rioja-Villodres, J.; Luis Carrillo, J.; Camacho, M.; Van Gils, M.; Sánchez-Chaparro, M.A.; Vanakker, O.; Valdivielso, P. Plasma Inorganic Pyrophosphate and Alkaline Phosphatase in Patients with Pseudoxanthoma Elasticum. Ann. Transl. Med. 2019, 7, 798. [Google Scholar] [CrossRef]
- Jansen, R.S.; Küçükosmanoǧlu, A.; De Haas, M.; Sapthu, S.; Otero, J.A.; Hegman, I.E.M.; Bergen, A.A.B.; Gorgels, T.G.M.F.; Borst, P.; Van De Wetering, K. ABCC6 Prevents Ectopic Mineralization Seen in Pseudoxanthoma Elasticum by Inducing Cellular Nucleotide Release. Proc. Natl. Acad. Sci. USA 2013, 110, 20206–20211. [Google Scholar] [CrossRef] [Green Version]
- Jansen, R.S.; Duijst, S.; Mahakena, S.; Sommer, D.; Szeri, F.; Váradi, A.; Plomp, A.; Bergen, A.A.; Oude Elferink, R.P.J.; Borst, P.; et al. ABCC6-Mediated ATP Secretion by the Liver Is the Main Source of the Mineralization Inhibitor Inorganic Pyrophosphate in the Systemic Circulation—Brief Report. Arteri. Thromb. Vasc. Biol. 2014, 34, 1985–1989. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, H.; Zebisch, M.; Sträter, N. Cellular Function and Molecular Structure of Ecto-Nucleotidases. Purinergic Signal. 2012, 8, 437–502. [Google Scholar] [CrossRef] [Green Version]
- Fredholm, B.B. Adenosine, an Endogenous Distress Signal, Modulates Tissue Damage and Repair. Cell Death Differ. 2007, 14, 1315–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z.W.; Dong, K.; Zhang, H.Z. The Roles of CD73 in Cancer. BioMed Res. Int. 2014, 1–9. [Google Scholar] [CrossRef]
- Shali, S.; Yu, J.; Zhang, X.; Wang, X.; Jin, Y.; Su, M.; Liao, X.; Yu, Y.; Zhi, X.; Zhou, P. Ecto-5’-nucleotidase (CD73) is a potential target of hepatocellular carcinoma. J. Cell Physiol. 2019, 234, 10248–10259. [Google Scholar] [CrossRef] [PubMed]
- Vijayan, D.; Barkauskas, D.S.; Stannard, K.; Sult, E.; Buonpane, R.; Takeda, K.; Teng, M.W.L.; Sachsenmeier, K.; Hay, C.; Smyth, M.J. Selective Activation of Anti-CD73 Mechanisms in Control of Primary Tumors and Metastases. Oncoimmunology 2017, 6, e1312044–e1312055. [Google Scholar] [CrossRef] [PubMed]
- Dumontet, C.; Peyrottes, S.; Rabeson, C.; Cros-Perrial, E.; Géant, P.Y.; Chaloin, L.; Jordheim, L.P. CD73 Inhibition by Purine Cytotoxic Nucleoside Analogue-Based Diphosphonates. Eur. J. Med. Chem. 2018, 157, 1051–1055. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Gu, Z.; Fan, Y.; Zhai, G.; Zhao, X.; Sun, Q.; Shi, Y.; Lin, G. Inhibition of the Adenosinergic Pathway: The Indispensable Part of Oncological Therapy in the Future. Purinergic Signal. 2019, 15, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Miglionico, R.; Armentano, M.F.; Carmosino, M.; Salvia, A.M.; Cuviello, F.; Bisaccia, F.; Ostuni, A. Dysregulation of Gene Expression in ABCC6 Knockdown HepG2 Cells. Cell. Mol. Biol. Lett. 2014, 19, 517–526. [Google Scholar] [CrossRef] [Green Version]
- Martinelli, F.; Cuviello, F.; Pace, M.C.; Armentano, M.F.; Miglionico, R.; Ostuni, A.; Bisaccia, F. Extracellular ATP Regulates CD73 and ABCC6 Expression in HepG2 Cells. Front. Mol. Biosci. 2018, 5, 75. [Google Scholar] [CrossRef] [Green Version]
- Miglionico, R.; Ostuni, A.; Armentano, M.F.; Milella, L.; Crescenzi, E.; Carmosino, M.; Bisaccia, F. ABCC6 Knockdown in HepG2 Cells Induces a Senescent-like Cell Phenotype. Cell. Mol. Biol. Lett. 2017, 22, 517–526. [Google Scholar] [CrossRef] [Green Version]
- Gillet, J.P.; Schneider, J.; Bertholet, V.; DE Longueville, F.; Remacle, J.; Efferth, T. Microarray Expression Profiling of ABC Transporters in Human Breast Cancer. Cancer Genom. Proteom. 2006, 3, 97–106. [Google Scholar]
- Soleymani Abyaneh, H.; Gupta, N.; Radziwon-Balicka, A.; Jurasz, P.; Seubert, J.; Lai, R.; Lavasanifar, A. STAT3 but Not HIF-1α Is Important in Mediating Hypoxia-Induced Chemoresistance in MDA-MB-231, a Triple Negative Breast Cancer Cell Line. Cancers (Basel) 2017, 9, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, H.; Daddysman, M.K.; Rankin, G.O.; Jiang, B.H.; Chen, Y.C. Kaempferol enhances cisplatin’s effect on ovarian cancer cells through promoting apoptosis caused by down regulation of cMyc. Cancer Cell Int. 2010, 10, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, H.-M.; Sohn, Y.-W.; Oh, S.-Y.; Kim, S.-H.; Beck, S.; Kim, S.; Kim, H. ID4 imparts chemoresistance and cancer stemness to glioma cells by derepressing miR-9*-mediated suppression of SOX2. Cancer Res. 2011, 71, 3410–3421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eadie, L.N.; Dang, P.; Goyne, J.M.; Hughes, T.P.; White, D.L. ABCC6 plays a significant role in the transport of nilotinib and dasatinib, and contributes to TKI resistance in vitro, in both cell lines and primary patient mononuclear cells. PLoS ONE 2018, 13, e0192180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deeken, J.F.; Cormier, T.; Price, D.K.; Sissung, T.M.; Steinberg, S.M.; Tran, K.; Liewehr, D.J.; Figg, D. A pharmacogenetic study of docetaxel and thalidomide in patients with castration-resistant prostate cancer using the DMET genotyping platform. Pharm. J. 2010, 10, 191–199. [Google Scholar] [CrossRef]
- Yu, J.; Zhu, T.; Wang, Z.R.; Zhang, H.Y.; Qain, Z.L.; Xu, H.L.; Gao, B.M.; Wang, W.; Gu, L.P.; Meng, J.; et al. A novel set of DNA methylation markers in urine sediments for sensitive/specific detection of bladder cancer. Clin. Cancer Res. 2007, 13, 7296–7304. [Google Scholar] [CrossRef] [Green Version]
- Fang, D.; He, S.; Xiong, G.; Singla, N.; Cao, Z.P.; Zhang, L.; Li, X.; Zhou, L.Q. Comparison of clinicopathologic characteristics, epigenetic biomarkers and prognosis between renal pelvic and ureteral tumors in upper tract urothelial carcinoma. BMC Urol. 2018, 18, 22. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; He, Z.; Zhou, F.; Huang, Y.; Zhu, G. Prognostic significance and molecular mechanisms of adenosine triphosphate-binding cassette subfamily C members in gastric cancer. Medicine (Baltimore) 2019, 98, e18347. [Google Scholar] [CrossRef]
- Ruiz-Pinto, S.; Pita, G.; Patiño-García, A.; García-Miguel, P.; Alonso, J.; Pérez-Martínez, A.; Sastre, A.; Gómez-Mariano, G.; Lissat, A.; Scotlandi, K.; et al. Identification of genetic variants in pharmacokinetic genes associated with Ewing Sarcoma treatment outcome. Ann. Oncol. 2016, 27, 1788–1793. [Google Scholar] [CrossRef]
- Salvia, A.M.; Cuviello, F.; Coluzzi, S.; Nuccorini, R.; Attolico, I.; Pascale, S.P.; Bisaccia, F.; Pizzuti, M.; Ostuni, A. Expression of Some ATP-Binding Cassette Transporters in Acute Myeloid Leukemia. Hematol. Rep. 2017, 9, 137–141. [Google Scholar] [CrossRef]
- Møller, J.V. The Tubular Site of Urate Transport in the Rabbit Kidney, and the Effect of Probenecid on Urate Secretion. Acta Pharmacol. Toxicol. (Copenh.) 1965, 23, 329–336. [Google Scholar] [CrossRef]
- Silverman, W.; Locovei, S.; Dahl, G. Probenecid, a Gout Remedy, Inhibits Pannexin 1 Channels. Am. J. Physiol. Cell Physiol. 2008, 295, C761–C767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuviello, F.; Bisaccia, F.; Spinelli, D.; Armentano, M.F.; Bufo, S.A.; Cosimelli, B.; Ostuni, A. The P-Glycoprotein Inhibitor Diltiazem-like 8-(4-Chlorophenyl)-5-Methyl-8-[(2Z)-Pent-2-En-1-Yloxy]-8H-[1,2,4]Oxadiazolo [3,4-c][1,4]Thiazin-3-One Inhibits Esterase Activity and H3 Histone Acetylation. Eur. J. Med. Chem. 2019, 164, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Laurenzana, I.; Caivano, A.; La Rocca, F.; Trino, S.; De Luca, L.; D’Alessio, F.; Schenone, S.; Falco, G.; Botta, M.; Del Vecchio, L.; et al. A Pyrazolo[3,4-d]Pyrimidine Compound Reduces Cell Viability and Induces Apoptosis in Different Hematological Malignancies. Front. Pharmacol. 2016, 7, 416. [Google Scholar] [CrossRef] [Green Version]
- Le Clainche, C.; Carlier, M.F. Regulation of Actin Assembly Associated with Protrusion and Adhesion in Cell Migration. Physiol. Rev. 2008, 88, 489–513. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Pei, Z.; Ji, C.; Zhang, X.; Xu, J.; Wang, J. Novel Insights into the Role of the Cytoskeleton in Cancer. Cytoskelet. Struct. Dyn. Funct. Dis. 2017, 64, 259–278. [Google Scholar] [CrossRef] [Green Version]
- Ran, Y.; Zheng, A.; Thibodeau, P.H. Structural analysis reveals pathomechanisms associated with pseudoxanthoma elasticum-causing mutations in the ABCC6 transporter. J. Biol. Chem. 2018, 293, 15855–15866. [Google Scholar] [CrossRef] [Green Version]
- Ostuni, A.; Miglionico, R.; Castiglione Morelli, M.A.; Bisaccia, F. Study of the Nucleotide-Binding Domain 1 of the Human Transporter Protein MRP6. Protein Pept. Lett. 2012, 17, 1553–1558. [Google Scholar] [CrossRef]
- Ran, Y.; Thibodeau, P.H. Stabilization of Nucleotide Binding Domain Dimers Rescues ABCC6 Mutants Associated with Pseudoxanthoma Elasticum. J. Biol. Chem. 2017, 292, 1559–1572. [Google Scholar] [CrossRef] [Green Version]
- Ostuni, A.; Miglionico, R.; Monné, M.; Castiglione Morelli, M.A.; Bisaccia, F. The Nucleotide-Binding Domain 2 of the Human Transporter Protein MRP6. J. Bioenerg. Biomembr. 2011, 43, 465–471. [Google Scholar] [CrossRef]
- Cuviello, F.; Tellgren-Roth, Å.; Lara, P.; Ruud Selin, F.; Monné, M.; Bisaccia, F.; Nilsson, I.; Ostuni, A. Membrane Insertion and Topology of the Amino-Terminal Domain TMD0 of Multidrug-Resistance Associated Protein 6 (MRP6). FEBS Lett. 2015, 589, 3921–3928. [Google Scholar] [CrossRef] [PubMed]
- Ostuni, A.; Castiglione Morelli, M.A.; Cuviello, F.; Bavoso, A.; Bisaccia, F. Structural characterization of the L0 cytoplasmic loop of human multidrug resistance protein 6 (MRP6). Biochim. Biophys. Acta Biomembr. 2019, 1861, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Boison, D.; Yegutkin, G.G. Adenosine Metabolism: Emerging Concepts for Cancer Therapy. Cancer Cell. 2019, 36, 582–596. [Google Scholar] [CrossRef] [PubMed]
- Di Virgilio, F.; Sarti, A.C.; Falzoni, S.; De Marchi, E.; Adinolfi, E. Extracellular ATP and P2 purinergic signalling in the tumour microenvironment. Nat. Rev. Cancer 2018, 18, 601–618. [Google Scholar] [CrossRef] [PubMed]
- Miglionico, R.; Gerbino, A.; Ostuni, A.; Armentano, M.F.; Monne, M.; Carmosino, M.; Bisaccia, F. New insights into the roles of the N-terminal region of the ABCC6 transporter. J. Bioenerg. Biomembr. 2016, 48, 259–267. [Google Scholar] [CrossRef]
- Sakthivel, K.M.; Sehgal, P. A novel role of lamins from genetic disease to cancer biomarkers. Oncol. Rev. 2016, 10, 309. [Google Scholar] [CrossRef]
- Gao, Z.W.; Wang, H.P.; Lin, F.; Wang, X.; Long, M.; Zhang, H.Z.; Dong, K. CD73 Promotes Proliferation and Migration of Human Cervical Cancer Cells Independent of Its Enzyme Activity. BMC Cancer 2017, 17, 135. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Forward Primer | Reverse Primer |
|---|---|---|---|
| β-actin | NM 001101.3 | 5′-CCTGGCACCCAGCACAAT-3′ | 5′-GCCGATCCACACGGAGTACT-3′ |
| Abcc6 | NM_001171.5 | 5′-AAGGAACCACCATCAGGAGGAG-3′ | 5′-ACCAGCGACACAGAGAAGAGG3′ |
| p21 | NM_000389 | 5′-CTGTCTTGTACCCTTGTGCCT-3′ | 5′-CGTTTGGAGTGGTAGAAATCTGTC-3′ |
| p53 | NM_001276760.1 | 5′-TGAATGAGGCCTTGGAACTC-3′ | 5′-ACTTCAGGTGGCTGGAGTG-3′ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ostuni, A.; Carmosino, M.; Miglionico, R.; Abruzzese, V.; Martinelli, F.; Russo, D.; Laurenzana, I.; Petillo, A.; Bisaccia, F. Inhibition of ABCC6 Transporter Modifies Cytoskeleton and Reduces Motility of HepG2 Cells via Purinergic Pathway. Cells 2020, 9, 1410. https://doi.org/10.3390/cells9061410
Ostuni A, Carmosino M, Miglionico R, Abruzzese V, Martinelli F, Russo D, Laurenzana I, Petillo A, Bisaccia F. Inhibition of ABCC6 Transporter Modifies Cytoskeleton and Reduces Motility of HepG2 Cells via Purinergic Pathway. Cells. 2020; 9(6):1410. https://doi.org/10.3390/cells9061410
Chicago/Turabian StyleOstuni, Angela, Monica Carmosino, Rocchina Miglionico, Vittorio Abruzzese, Fabio Martinelli, Daniela Russo, Ilaria Laurenzana, Agata Petillo, and Faustino Bisaccia. 2020. "Inhibition of ABCC6 Transporter Modifies Cytoskeleton and Reduces Motility of HepG2 Cells via Purinergic Pathway" Cells 9, no. 6: 1410. https://doi.org/10.3390/cells9061410
APA StyleOstuni, A., Carmosino, M., Miglionico, R., Abruzzese, V., Martinelli, F., Russo, D., Laurenzana, I., Petillo, A., & Bisaccia, F. (2020). Inhibition of ABCC6 Transporter Modifies Cytoskeleton and Reduces Motility of HepG2 Cells via Purinergic Pathway. Cells, 9(6), 1410. https://doi.org/10.3390/cells9061410