Increased Leaf Nicotine Content by Targeting Transcription Factor Gene Expression in Commercial Flue-Cured Tobacco (Nicotiana tabacum L.)



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Transformation
2.2. DNA Cloning and Vector Construction
2.3. RNA Isolation and Quantitative Reverse-Transcription Polymerase Chain Reaction (qRT-PCR)
2.4. Alkaloid Extraction and Thin Layer Chromatography (TLC)
2.5. Gas Chromatography-Mass Spectrometry (GC-MS) Measurement of Nicotine
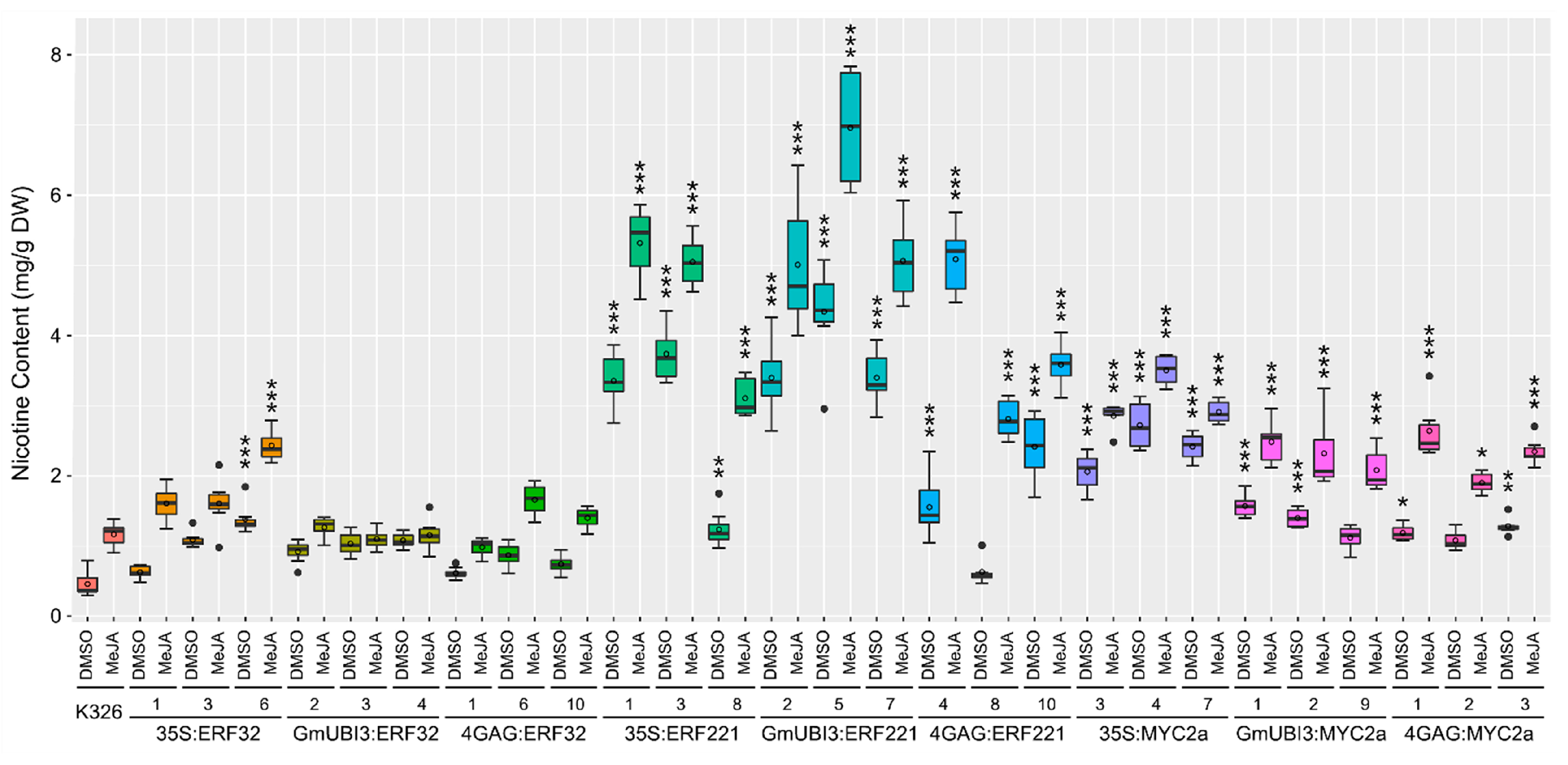
3. Results
3.1. Constitutive and Inducible Overexpression of Transcription factors NtERF32, NtERF221, and NtMYC2a Increases Nicotine Contents
3.2. Effects of Transcription Factors (TF) Overexpression on the Expression of Genes Involved in Nicotine Biosynthesis and Transport
3.3. Effects of the Overexpression of Biosynthetic Enzymes on Nicotine Accumulation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kessler, A.; Baldwin, I.T. Plant responses to insect herbivory: The emerging molecular analysis. Annu. Rev. Plant Biol. 2002, 53, 299–328. [Google Scholar] [CrossRef] [PubMed]
- Kessler, A.; Baldwin, I.T. Herbivore-induced plant vaccination. Part I. The orchestration of plant defenses in nature and their fitness consequences in the wild tobacco Nicotiana attenuata. Plant J. 2004, 38, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Steppuhn, A.; Gase, K.; Krock, B.; Halitschke, R.; Baldwin, I.T. Nicotine’s defensive function in nature. PLoS Biol. 2004, 2, e217. [Google Scholar] [CrossRef] [PubMed]
- Dewey, R.E.; Xie, J. Molecular genetics of alkaloid biosynthesis in Nicotiana tabacum. Phytochemistry 2013, 94, 10–27. [Google Scholar] [CrossRef]
- Saitoh, F.; Noma, M.; Kawashima, N. The Alkaloid Contents of 60 Nicotiana Species. Phytochemistry 1985, 24, 477–480. [Google Scholar] [CrossRef]
- Saunders, J.W.; Bush, L.P. Nicotine Biosynthetic Enzyme Activities in Nicotiana tabacum L. Genotypes with Different Alkaloid Levels. Plant Physiol. 1979, 64, 236–240. [Google Scholar] [CrossRef]
- Baldwin, I.T. Mechanism of damage-induced alkaloid production in wild tobacco. J. Chem. Ecol. 1989, 15, 1661–1680. [Google Scholar] [CrossRef]
- Baldwin, I.T. Jasmonate-induced responses are costly but benefit plants under attack in native populations. Proc. Natl. Acad. Sci. USA 1998, 95, 8113–8118. [Google Scholar] [CrossRef]
- Baldwin, I.T.; Schmelz, E.A.; Ohnmeiss, T.E. Wound-induced changes in root and shoot jasmonic acid pools correlate with induced nicotine synthesis in Nicotiana sylvestris spegazzini and comes. J. Chem. Ecol. 1994, 20, 2139–2157. [Google Scholar] [CrossRef]
- Baldwin, I.T. Methyl jasmonate-induced nicotine production in Nicotiana attenuata: Inducing defenses in the field without wounding. Entomol. Exp. Appl. 1996, 80, 213–220. [Google Scholar] [CrossRef]
- Morita, M.; Shitan, N.; Sawada, K.; Van Montagu, M.C.E.; Inze, D.; Rischer, H.; Goossens, A.; Oksman-Caldentey, K.M.; Moriyama, Y.; Yazaki, K. Vacuolar transport of nicotine is mediated by a multidrug and toxic compound extrusion (MATE) transporter in Nicotiana tabacum. Proc. Natl. Acad. Sci. USA 2009, 106, 2447–2452. [Google Scholar] [CrossRef] [PubMed]
- Shoji, T.; Inai, K.; Yazaki, Y.; Sato, Y.; Takase, H.; Shitan, N.; Yazaki, K.; Goto, Y.; Toyooka, K.; Matsuoka, K.; et al. Multidrug and Toxic Compound Extrusion-Type Transporters Implicated in Vacuolar Sequestration of Nicotine in Tobacco Roots. Plant Physiol. 2009, 149, 708–718. [Google Scholar] [CrossRef] [PubMed]
- Shitan, N.; Minami, S.; Morita, M.; Hayashida, M.; Ito, S.; Takanashi, K.; Omote, H.; Moriyama, Y.; Sugiyama, A.; Goossens, A.; et al. Involvement of the Leaf-Specific Multidrug and Toxic Compound Extrusion (MATE) Transporter Nt-JAT2 in Vacuolar Sequestration of Nicotine in Nicotiana tabacum. PLoS ONE 2014, 9, e108789. [Google Scholar] [CrossRef] [PubMed]
- Bush, L.; Hempfling, P.W.; Burton, H. Chapter 2—Biosynthesis of nicotine and related compounds. In Analytical Determination of Nicotine and Related Compounds and their Metabolites; John, W., Gorrod, P.J., Eds.; Elsevier Science: Amsterdam, The Netherlands, 1999; pp. 13–44. [Google Scholar]
- Ziegler, J.; Facchini, P.J. Alkaloid Biosynthesis: Metabolism and Trafficking. Annu. Rev. Plant Biol. 2008, 59, 735–769. [Google Scholar] [CrossRef] [PubMed]
- Shoji, T.; Hashimoto, T. Recruitment of a duplicated primary metabolism gene into the nicotine biosynthesis regulon in tobacco. Plant J. 2011, 67, 949–959. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Yamada, Y. Alkaloid Biogenesis: Molecular Aspects. Annu. Rev. Plant Biol. 1994, 45, 257–285. [Google Scholar] [CrossRef]
- Imanishi, S.; Hashizume, K.; Nakakita, M.; Kojima, H.; Matsubayashi, Y.; Hashimoto, T.; Sakagami, Y.; Yamada, Y.; Nakamura, K. Differential induction by methyl jasmonate of genes encoding ornithine decarboxylase and other enzymes involved in nicotine biosynthesis in tobacco cell cultures. Plant Mol. Biol. 1998, 38, 1101–1111. [Google Scholar] [CrossRef]
- Riechers, D.E.; Timko, M.P. Structure and expression of the gene family encoding putrescine N-methyltransferase in Nicotiana tabacum: New clues to the evolutionary origin of cultivated tobacco. Plant Mol. Biol. 1999, 41, 387–401. [Google Scholar] [CrossRef]
- Bortolotti, C.; Cordeiro, A.; Alcazar, R.; Borrell, A.; Culianez-Macia, F.A.; Tiburcio, A.F.; Altabella, T. Localization of arginine decarboxylase in tobacco plants. Physiol. Plant. 2004, 120, 84–92. [Google Scholar] [CrossRef]
- Xu, B.F.; Timko, M.P. Methyl jasmonate induced expression of the tobacco putrescine N-methyltransferase genes requires both G-box and GCC-motif elements. Plant Mol. Biol. 2004, 55, 743–761. [Google Scholar] [CrossRef]
- Heim, W.G.; Sykes, K.A.; Hildreth, S.B.; Sun, J.; Lu, R.H.; Jelesko, J.G. Cloning and characterization of a Nicotiana tabacum methylputrescine oxidase transcript. Phytochemistry 2007, 68, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Katoh, A.; Shoji, T.; Hashimoto, T. Molecular Cloning of N-methylputrescine Oxidase from Tobacco. Plant Cell Physiol. 2007, 48, 550–554. [Google Scholar] [CrossRef]
- Sinclair, S.J.; Murphy, K.J.; Birch, C.D.; Hamill, J.D. Molecular characterization of quinolinate phosphoribosyltransferase (QPRtase) in Nicotiana. Plant Mol. Biol. 2000, 44, 603–617. [Google Scholar] [CrossRef] [PubMed]
- Katoh, A.; Uenohara, K.; Akita, M.; Hashimoto, T. Early Steps in the Biosynthesis of NAD in Arabidopsis Start with Aspartate and Occur in the Plastid. Plant Physiol. 2006, 141, 851–857. [Google Scholar] [CrossRef] [PubMed]
- Ryan, S.M.; Cane, K.A.; DeBoer, K.D.; Sinclair, S.J.; Brimblecombe, R.; Hamill, J.D. Structure and expression of the quinolinate phosphoribosyltransferase (QPT) gene family in Nicotiana. Plant Sci. 2012, 188, 102–110. [Google Scholar] [CrossRef]
- DeBoer, K.D.; Lye, J.C.; Aitken, C.D.; Su, A.K.K.; Hamill, J.D. The A622 gene in Nicotiana glauca (tree tobacco): Evidence for a functional role in pyridine alkaloid synthesis. Plant Mol. Biol. 2009, 69, 299–312. [Google Scholar] [CrossRef]
- Kajikawa, M.; Hirai, N.; Hashimoto, T. A PIP-family protein is required for biosynthesis of tobacco alkaloids. Plant Mol. Biol. 2009, 69, 287–298. [Google Scholar] [CrossRef]
- Kajikawa, M.; Shoji, T.; Kato, A.; Hashimoto, T. Vacuole-Localized Berberine Bridge Enzyme-Like Proteins Are Required for a Late Step of Nicotine Biosynthesis in Tobacco. Plant Physiol. 2011, 155, 2010–2022. [Google Scholar] [CrossRef]
- De Sutter, V.; Vanderhaeghen, R.; Tilleman, S.; Lammertyn, F.; Vanhoutte, I.; Karimi, M.; Inze, D.; Goossens, A.; Hilson, P. Exploration of jasmonate signalling via automated and standardized transient expression assays in tobacco cells. Plant J. 2005, 44, 1065–1076. [Google Scholar] [CrossRef]
- Rushton, P.J.; Bokowiec, M.T.; Han, S.C.; Zhang, H.B.; Brannock, J.F.; Chen, X.F.; Laudeman, T.W.; Timko, M.P. Tobacco transcription factors: Novel insights into transcriptional regulation in the Solanaceae. Plant Physiol. 2008, 147, 280–295. [Google Scholar] [CrossRef]
- Shoji, T.; Kajikawa, M.; Hashimoto, T. Clustered transcription factor genes regulate nicotine biosynthesis in tobacco. Plant Cell 2010, 22, 3390–3409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todd, A.T.; Liu, E.; Polvi, S.L.; Pammett, R.T.; Page, J.E. A functional genomics screen identifies diverse transcription factors that regulate alkaloid biosynthesis in Nicotiana benthamiana. Plant J. 2010, 62, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Shoji, T.; Hashimoto, T. DNA-binding and transcriptional activation properties of tobacco NIC2-locus ERF189 and related transcription factors. Plant Biotechnol. 2012, 29, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Sears, M.T.; Zhang, H.B.; Rushton, P.J.; Wu, M.; Han, S.C.; Spano, A.J.; Timko, M.P. NtERF32: A non-NIC2 locus AP2/ERF transcription factor required in jasmonate-inducible nicotine biosynthesis in tobacco. Plant Mol. Biol. 2014, 84, 49–66. [Google Scholar] [CrossRef] [PubMed]
- DeBoer, K.D.; Tilleman, S.; Pauwels, L.; Vanden Bossche, R.; De Sutter, V.; Vanderhaeghen, R.; Hilson, P.; Hamill, J.D.; Goossens, A. APETALA2/ETHYLENE RESPONSE FACTOR and basic helix-loop-helix tobacco transcription factors cooperatively mediate jasmonate-elicited nicotine biosynthesis. Plant J. 2011, 66, 1053–1065. [Google Scholar] [CrossRef] [PubMed]
- Shoji, T.; Mishima, M.; Hashimoto, T. Divergent DNA-Binding Specificities of a Group of ETHYLENE RESPONSE FACTOR Transcription Factors Involved in Plant Defense. Plant Physiol. 2013, 162, 977–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheard, L.B.; Tan, X.; Mao, H.B.; Withers, J.; Ben-Nissan, G.; Hinds, T.R.; Kobayashi, Y.; Hsu, F.F.; Sharon, M.; Browse, J.; et al. Jasmonate perception by inositol phosphate-potentiated COI1-JAZ co-receptor. Nature 2010, 468, 400–405. [Google Scholar] [CrossRef]
- Zhang, F.; Yao, J.; Ke, J.Y.; Zhang, L.; Lam, V.Q.; Xin, X.F.; Zhou, X.E.; Chen, J.; Brunzelle, J.; Griffin, P.R.; et al. Structural basis of JAZ repression of MYC transcription factors in jasmonate signalling. Nature 2015, 525, 269–273. [Google Scholar] [CrossRef] [Green Version]
- Chini, A.; Fonseca, S.; Fernandez, G.; Adie, B.; Chico, J.M.; Lorenzo, O.; Garcia-Casado, G.; Lopez-Vidriero, I.; Lozano, F.M.; Ponce, M.R.; et al. The JAZ family of repressors is the missing link in jasmonate signalling. Nature 2007, 448, 666–671. [Google Scholar] [CrossRef]
- Thines, B.; Katsir, L.; Melotto, M.; Niu, Y.; Mandaokar, A.; Liu, G.H.; Nomura, K.; He, S.Y.; Howe, G.A.; Browse, J. JAZ repressor proteins are targets of the SCFCO11 complex during jasmonate signalling. Nature 2007, 448, U661–U662. [Google Scholar] [CrossRef]
- Browse, J. Jasmonate Passes Muster: A Receptor and Targets for the Defense Hormone. Annu. Rev. Plant Biol. 2009, 60, 183–205. [Google Scholar] [CrossRef] [PubMed]
- Pauwels, L.; Barbero, G.F.; Geerinck, J.; Tilleman, S.; Grunewald, W.; Perez, A.C.; Chico, J.M.; Bossche, R.V.; Sewell, J.; Gil, E.; et al. NINJA connects the co-repressor TOPLESS to jasmonate signalling. Nature 2010, 464, 788–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.; Jiang, H.; Li, L.; Zhai, Q.; Qi, L.; Zhou, W.; Liu, X.; Li, H.; Zheng, W.; Sun, J.; et al. The Arabidopsis mediator subunit MED25 differentially regulates jasmonate and abscisic acid signaling through interacting with the MYC2 and ABI5 transcription factors. Plant Cell 2012, 24, 2898–2916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, C.; Li, L.; Zhai, Q.; You, Y.; Deng, L.; Wu, F.; Chen, R.; Jiang, H.; Wang, H.; Chen, Q.; et al. Mediator subunit MED25 links the jasmonate receptor to transcriptionally active chromatin. Proc. Natl. Acad. Sci. USA 2017, 114, E8930–E8939. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.-B.; Bokowiec, M.T.; Rushton, P.J.; Han, S.-C.; Timko, M.P. Tobacco Transcription Factors NtMYC2a and NtMYC2b Form Nuclear Complexes with the NtJAZ1 Repressor and Regulate Multiple Jasmonate-Inducible Steps in Nicotine Biosynthesis. Mol. Plant 2012, 5, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.P.; Guo, J.; Yan, P.C.; Li, Y.S.; Liu, K.; Gao, P.; Zhao, H.P.; Chen, Y.B.; Wang, Y.D.; Timko, M.P.; et al. Transcriptome Profiling Identified Multiple Jasmonate ZIM-Domain Proteins Involved in the Regulation of Alkaloid Biosynthesis in Tobacco BY-2 Cells. Plant Mol. Biol. Report. 2015, 33, 153–166. [Google Scholar] [CrossRef]
- Chadick, J.Z.; Asturias, F.J. Structure of eukaryotic Mediator complexes. Trends Biochem. Sci. 2005, 30, 264–271. [Google Scholar] [CrossRef]
- Malik, S.; Roeder, R.G. Dynamic regulation of pol II transcription by the mammalian Mediator complex. Trends Biochem. Sci. 2005, 30, 256–263. [Google Scholar] [CrossRef]
- Liu, Y.; Du, M.; Deng, L.; Shen, J.; Fang, M.; Chen, Q.; Lu, Y.; Wang, Q.; Li, C.; Zhai, Q. MYC2 Regulates the Termination of Jasmonate Signaling via an Autoregulatory Negative Feedback Loop. Plant Cell 2019, 31, 106–127. [Google Scholar] [CrossRef] [Green Version]
- Shoji, T.; Hashimoto, T. Tobacco MYC2 Regulates Jasmonate-Inducible Nicotine Biosynthesis Genes Directly and By Way of the NIC2-Locus ERF Genes. Plant Cell Physiol. 2011, 52, 1117–1130. [Google Scholar] [CrossRef] [Green Version]
- Shoji, T.; Hashimoto, T. Stress-induced expression of NICOTINE2-locus genes and their homologs encoding Ethylene Response Factor transcription factors in tobacco. Phytochemistry 2015, 113, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Voelckel, C.; Krugel, T.; Gase, K.; Heidrich, N.; van Dam, N.M.; Winz, R.; Baldwin, I.T. Anti-sense expression of putrescine N-methyltransferase confirms defensive role of nicotine in Nicotiana sylvestris against Manduca sexta. Chemoecology 2001, 11, 121–126. [Google Scholar] [CrossRef]
- Chintapakorn, Y.; Hamill, J.D. Antisense-mediated down-regulation of putrescine N-methyltransferase activity in transgenic Nicotiana tabacum L. can lead to elevated levels of anatabine at the expense of nicotine. Plant Mol. Biol. 2003, 53, 87–105. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zeng, J.; Liang, Z.F.; Miao, Z.Q.; Sun, X.F.; Tang, K.X. Silencing of PMT expression caused a surge of anatabine accumulation in tobacco. Mol. Biol. Rep. 2009, 36, 2285–2289. [Google Scholar] [CrossRef]
- DeBoer, K.D.; Dalton, H.L.; Edward, F.J.; Hamill, J.D. RNAi-mediated down-regulation of ornithine decarboxylase (ODC) leads to reduced nicotine and increased anatabine levels in transgenic Nicotiana tabacum L. Phytochemistry 2011, 72, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Dalton, H.L.; Blomstedt, C.K.; Neale, A.D.; Gleadow, R.; DeBoer, K.D.; Hamill, J.D. Effects of down-regulating ornithine decarboxylase upon putrescine-associated metabolism and growth in Nicotiana tabacum L. J. Exp. Bot. 2016, 67, 3367–3381. [Google Scholar] [CrossRef]
- Farsalinos, K.E.; Polosa, R. Safety evaluation and risk assessment of electronic cigarettes as tobacco cigarette substitutes: A systematic review. Ther. Adv. Drug Saf. 2014, 5, 67–86. [Google Scholar] [CrossRef] [Green Version]
- Glasser, A.M.; Collins, L.; Pearson, J.L.; Abudayyeh, H.; Niaura, R.S.; Abrams, D.B.; Villanti, A.C. Overview of Electronic Nicotine Delivery Systems: A Systematic Review. Am. J. Prev. Med. 2017, 52, e33–e66. [Google Scholar] [CrossRef] [Green Version]
- Quik, M.; Perez, X.A.; Bordia, T. Nicotine as a potential neuroprotective agent for Parkinson’s disease. Mov. Disord. 2012, 27, 947–957. [Google Scholar] [CrossRef] [Green Version]
- Quik, M.; Boyd, J.T.; Bordia, T.; Perez, X. Potential Therapeutic Application for Nicotinic Receptor Drugs in Movement Disorders. Nicotine Tob. Res. 2019, 21, 357–369. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Horsch, R.B.; Fry, J.E.; Hoffmann, N.L.; Eichholtz, D.; Rogers, S.G.; Fraley, R.T. A Simple and General-Method for Transferring Genes into Plants. Science 1985, 227, 1229–1231. [Google Scholar]
- Schmidt, G.W.; Delaney, S.K. Stable internal reference genes for normalization of real-time RT-PCR in tobacco (Nicotiana tabacum) during development and abiotic stress. Mol. Genet. Genom. 2010, 283, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Goossens, A.; Hakkinen, S.T.; Laakso, I.; Seppanen-Laakso, T.; Biondi, S.; De Sutter, V.; Lammertyn, F.; Nuutila, A.M.; Soderlund, H.; Zabeau, M.; et al. A functional genomics approach toward the understanding of secondary metabolism in plant cells. Proc. Natl. Acad. Sci. USA 2003, 100, 8595–8600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez-Garcia, C.M.; Bouchard, R.A.; Rushton, P.J.; Jones, M.L.; Chen, X.; Timko, M.P.; Finer, J.J. High level transgenic expression of soybean (Glycine max) GmERF and Gmubi gene promoters isolated by a novel promoter analysis pipeline. BMC Plant Biol. 2010, 10, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, F.; Hashimoto, T.; Hachiya, A.; Tamura, K.; Choi, K.B.; Morishige, T.; Fujimoto, H.; Yamada, Y. Metabolic engineering of plant alkaloid biosynthesis. Proc. Natl. Acad. Sci. USA 2001, 98, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Li, F.F.; Wang, W.D.; Zhao, N.; Xiao, B.G.; Cao, P.J.; Wu, X.F.; Ye, C.Y.; Shen, E.H.; Qiu, J.; Zhu, Q.H.; et al. Regulation of Nicotine Biosynthesis by an Endogenous Target Mimicry of MicroRNA in Tobacco. Plant Physiol. 2015, 169, 1062–1071. [Google Scholar] [CrossRef] [Green Version]
- Legg, P.D.; Collins, G.B. Inheritance of Per Cent Total Alkaloids in Nicotiana tabacum L. 2. Genetic Effects of 2 Loci in Burley 21 X La Burley 21 Populations. Can. J. Genet. Cytol. 1971, 13, 287–291. [Google Scholar] [CrossRef]
- Hibi, N.; Higashiguchi, S.; Hashimoto, T.; Yamada, Y. Gene Expression in Tobacco Low-Nicotine Mutants. Plant Cell 1994, 6, 723–725. [Google Scholar]
- Cane, K.A.; Mayer, M.; Lidgett, A.J.; Michael, A.J.; Hamill, J.D. Molecular analysis of alkaloid metabolism in AABB v. aabb genotype Nicotiana tabacum in response to wounding of aerial tissues and methyl jasmonate treatment of cultured roots. Funct. Plant Biol. 2005, 32, 305–320. [Google Scholar] [CrossRef]
- Kidd, S.K.; Melillo, A.A.; Lu, R.H.; Reed, D.G.; Kuno, N.; Uchida, K.; Furuya, M.; Jelesko, J.G. The A and B Loci in Tobacco Regulate a Network of Stress Response Genes, Few of which are Associated with Nicotine Biosynthesis. Plant Mol. Biol. 2006, 60, 699–716. [Google Scholar] [CrossRef] [PubMed]
- Ohme-Takagi, M.; Shinshi, H. Ethylene-Inducible DNA Binding Proteins That Interact with an Ethylene-Responsive Element. Plant Cell 1995, 7, 173–182. [Google Scholar] [PubMed] [Green Version]
- Fischer, U.; Dröge-Laser, W. Overexpression of NtERF5, a New Member of the Tobacco Ethylene Response Transcription Factor Family Enhances Resistance to Tobacco mosaic virus. Mol. Plant-Microbe Interact. 2004, 17, 1162–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, T.; Nishiuchi, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Studies on Transcriptional Regulation of Endogenous Genes by ERF2 Transcription Factor in Tobacco Cells. Plant Cell Physiol. 2006, 47, 554–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsir, L.; Chung, H.S.; Koo, A.J.; Howe, G.A. Jasmonate signaling: A conserved mechanism of hormone sensing. Curr. Opin. Plant Biol. 2008, 11, 428–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Geyter, N.; Gholami, A.; Goormachtig, S.; Goossens, A. Transcriptional machineries in jasmonate-elicited plant secondary metabolism. Trends Plant Sci. 2012, 17, 349–359. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Kotova, T.I.; Timko, M.P. Increased Leaf Nicotine Content by Targeting Transcription Factor Gene Expression in Commercial Flue-Cured Tobacco (Nicotiana tabacum L.). Genes 2019, 10, 930. https://doi.org/10.3390/genes10110930
Liu H, Kotova TI, Timko MP. Increased Leaf Nicotine Content by Targeting Transcription Factor Gene Expression in Commercial Flue-Cured Tobacco (Nicotiana tabacum L.). Genes. 2019; 10(11):930. https://doi.org/10.3390/genes10110930
Chicago/Turabian StyleLiu, Hai, Tatyana I. Kotova, and Michael P. Timko. 2019. "Increased Leaf Nicotine Content by Targeting Transcription Factor Gene Expression in Commercial Flue-Cured Tobacco (Nicotiana tabacum L.)" Genes 10, no. 11: 930. https://doi.org/10.3390/genes10110930
APA StyleLiu, H., Kotova, T. I., & Timko, M. P. (2019). Increased Leaf Nicotine Content by Targeting Transcription Factor Gene Expression in Commercial Flue-Cured Tobacco (Nicotiana tabacum L.). Genes, 10(11), 930. https://doi.org/10.3390/genes10110930