Identification of Novel Interaction Partners of Ets-1: Focus on DNA Repair


Abstract
:1. Introduction
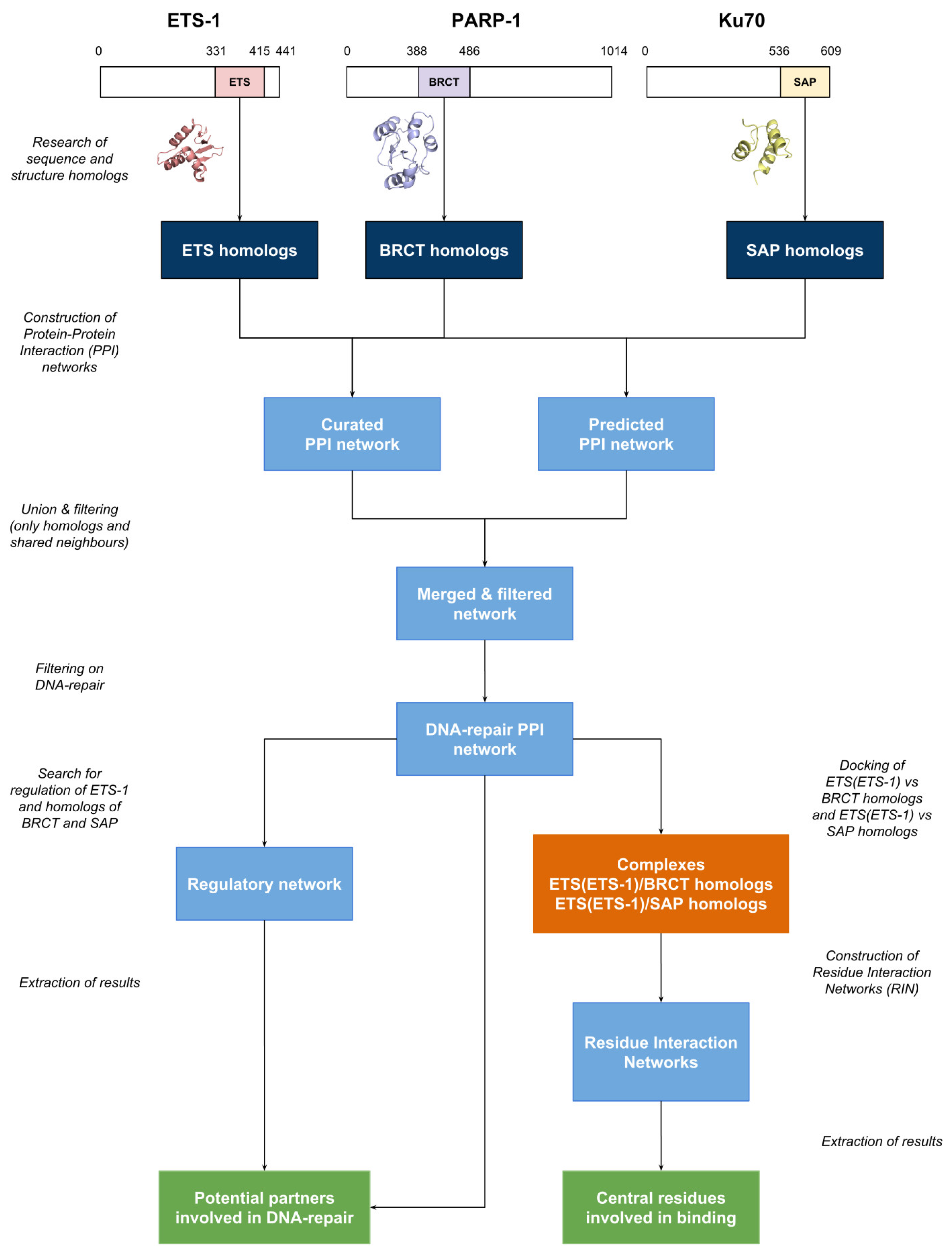
2. Materials and Methods
2.1. Homologs
- -
- ETS: P14921, residues 331 to 415
- -
- BRCT: P09874, residues 388 to 486
- -
- SAP: P12956, residues 536 to 609
- -
- ETS: 1GVJ (331–415)
- -
- BRCT: I-Tasser [7] model (388–486) based on the 2COK structure (BRCT domain which bears a mutation P480S)
- -
- SAP: I-Tasser model (536–609) based on the 1JEQ structure (limited SAP domain)
2.2. Domain-Domain Docking
2.3. Biological Networks
2.4. Expression Data
3. Results
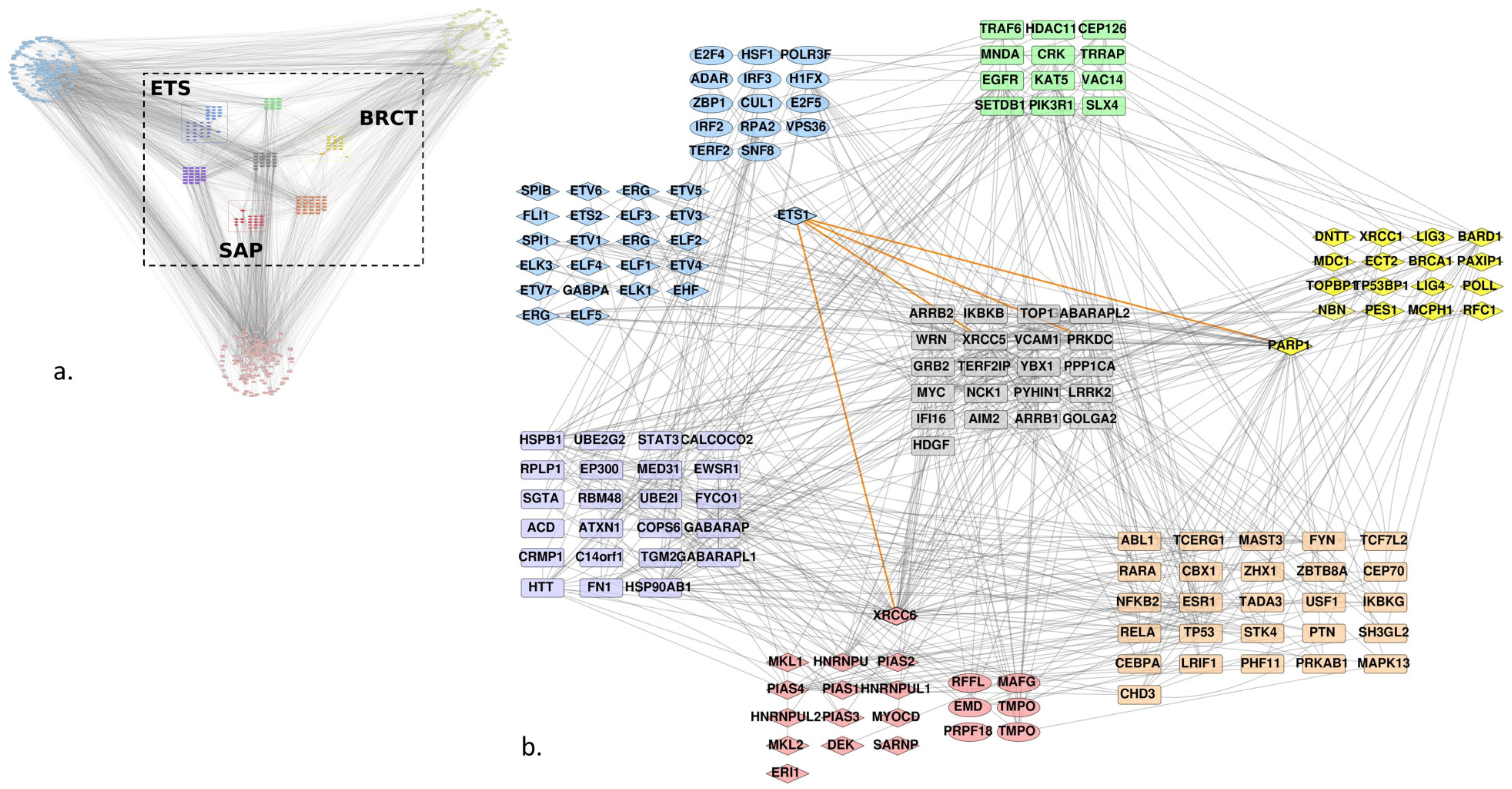
3.1. Homologs
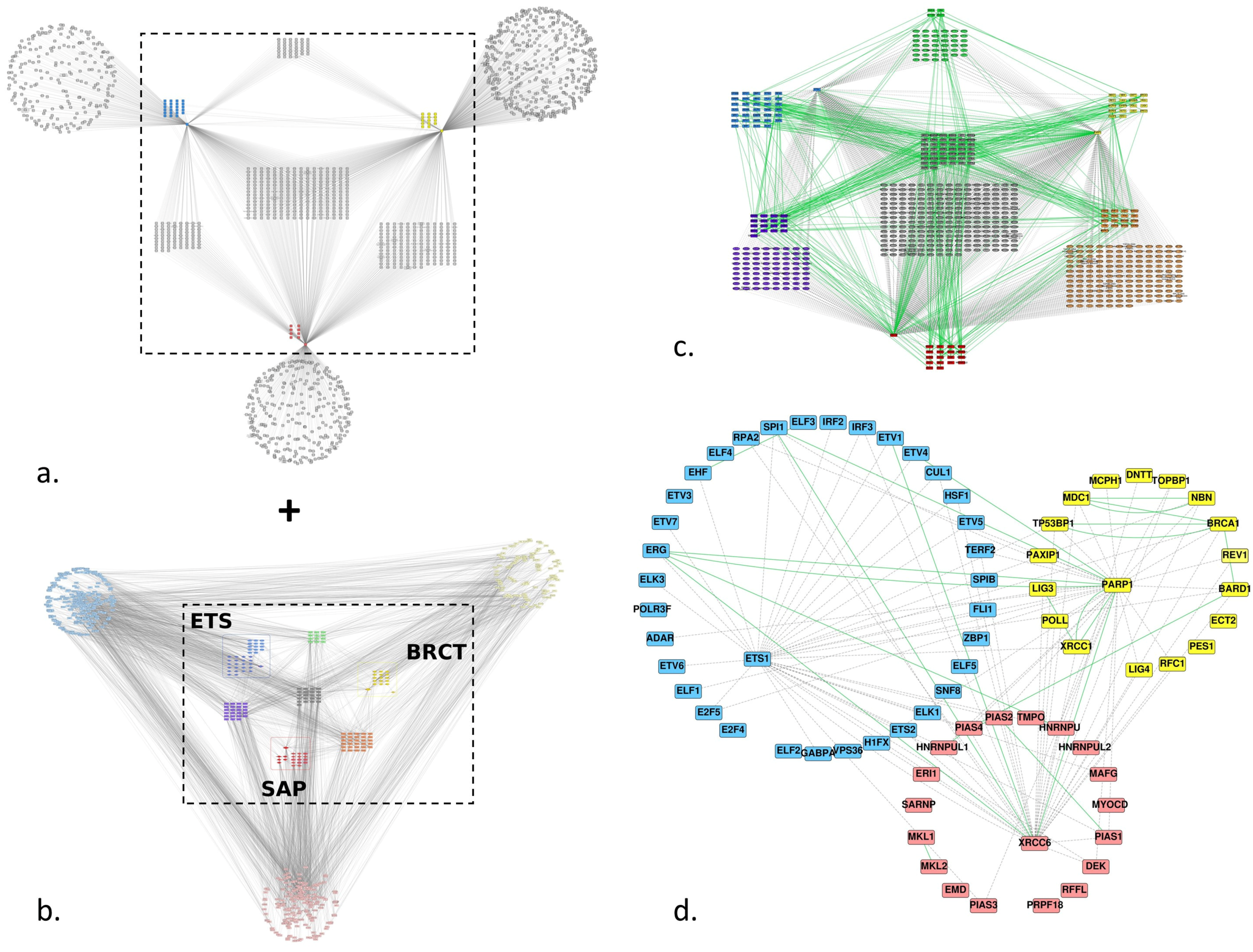
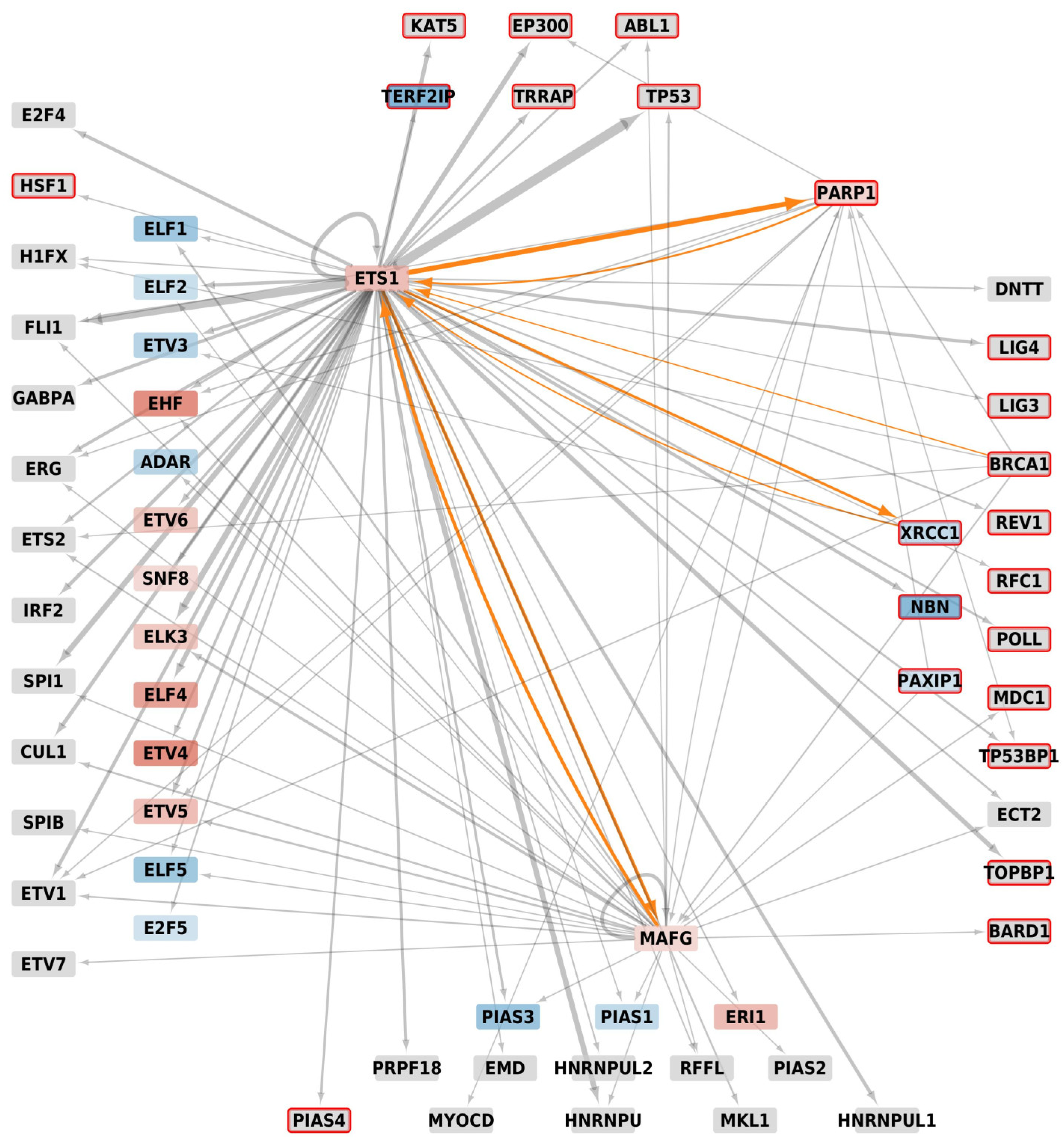
3.2. Protein-Protein Interaction Network
3.2.1. Experimentally Characterized Interactions
3.2.2. Predicted Interactions
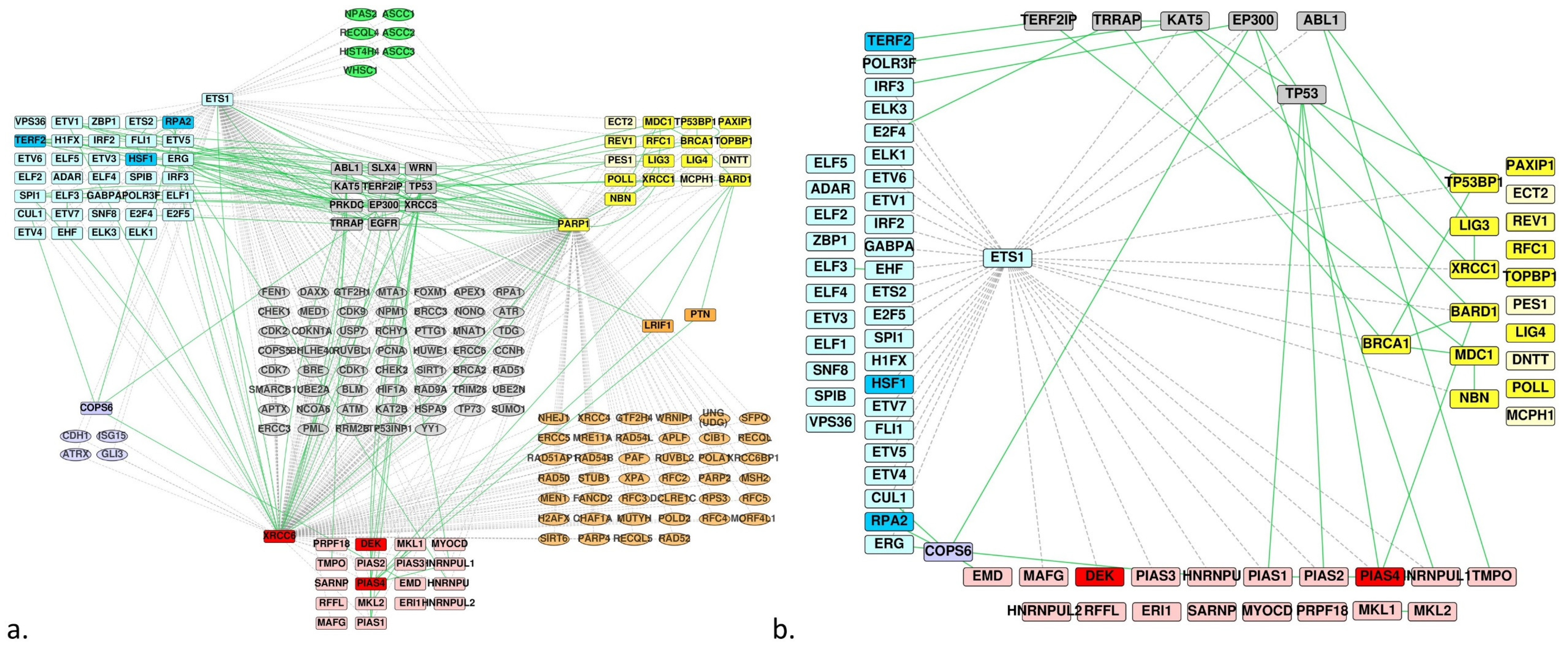
3.3. DNA Repair
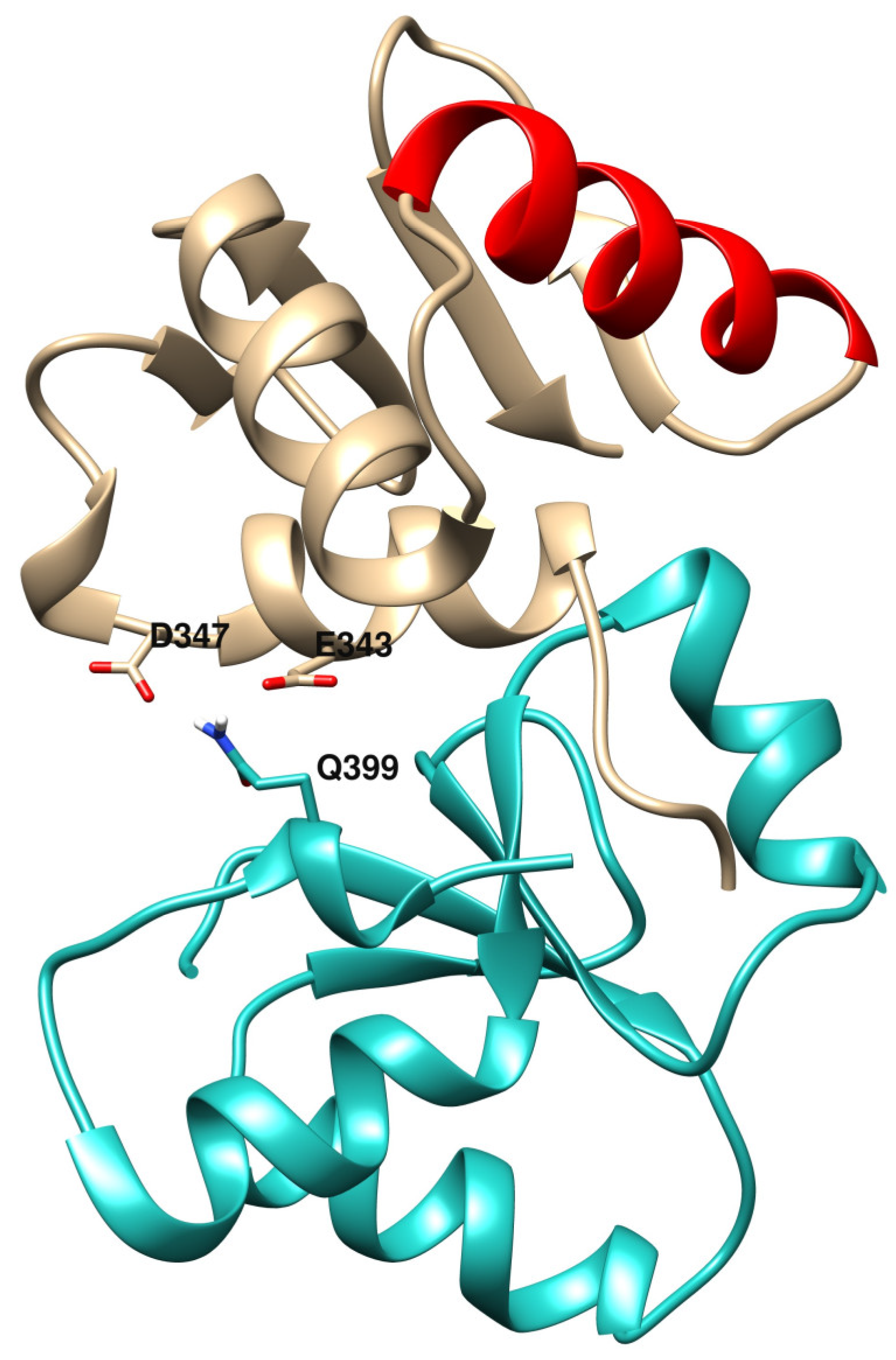
3.4. Protein-Protein Docking and Residue Interaction Networks
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

References
- Legrand, A.J.; Choul-Li, S.; Spriet, C.; Idziorek, T.; Vicogne, D.; Drobecq, H.; Dantzer, F.; Villeret, V.; Aumercier, M. The level of Ets-1 protein is regulated by poly(ADP-ribose) polymerase-1 (PARP-1) in cancer cells to prevent DNA damage. PLoS ONE 2013, 8, e55883. [Google Scholar] [CrossRef] [PubMed]
- Choul-Li, S.; Legrand, A.J.; Vicogne, D.; Villeret, V.; Aumercier, M. Ets-1 interacts through a similar binding interface with Ku70 and Poly (ADP-Ribose) Polymerase-1. Biosci. Biotechnol. Biochem. 2018, 82, 1753–1759. [Google Scholar] [CrossRef] [PubMed]
- De Ruyck, J.; Brysbaert, G.; Villeret, V.; Aumercier, M.; Lensink, M.F. Computational characterization of the binding mode between oncoprotein Ets-1 and DNA repair enzymes. Proteins 2018, 86, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Chae, Y.K.; Anker, J.F.; Carneiro, B.A.; Chandra, S.; Kaplan, J.; Kalyan, A.; Santa-Maria, C.A.; Platanias, L.C.; Giles, F.J. Genomic landscape of DNA repair genes in cancer. Oncotarget 2016, 7, 23312–23321. [Google Scholar] [CrossRef] [PubMed]
- Chacko, P.; Rajan, B.; Joseph, T.; Mathew, B.S.; Pillai, M.R. Polymorphisms in DNA repair gene XRCC1 and increased genetic susceptibility to breast cancer. Breast Cancer Res. Treat. 2005, 89, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Nickoloff, J.A.; Jones, D.; Lee, S.-H.; Williamson, E.A.; Hromas, R. Drugging the Cancers Addicted to DNA Repair. J. Natl. Cancer Inst. 2017, 109. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Ye, J.; McGinnis, S.; Madden, T.L. BLAST: Improvements for better sequence analysis. Nucleic Acids Res. 2006, 34, W6–W9. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef] [PubMed]
- Holm, L.; Laakso, L.M. Dali server update. Nucleic Acids Res. 2016, 44, W351–W355. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, D.W.; Ghoorah, A.W.; Mavridis, L.; Venkatraman, V. Fast protein structure alignment using Gaussian overlap scoring of backbone peptide fragment similarity. Bioinformatics 2012, 28, 3274–3281. [Google Scholar] [CrossRef] [PubMed]
- Dawson, N.L.; Lewis, T.E.; Das, S.; Lees, J.G.; Lee, D.; Ashford, P.; Orengo, C.A.; Sillitoe, I. CATH: An expanded resource to predict protein function through structure and sequence. Nucleic Acids Res. 2017, 45, D289–D295. [Google Scholar] [CrossRef] [PubMed]
- Fox, N.K.; Brenner, S.E.; Chandonia, J.-M. SCOPe: Structural Classification of Proteins—Extended, integrating SCOP and ASTRAL data and classification of new structures. Nucleic Acids Res. 2014, 42, D304–D309. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhang, Y. How significant is a protein structure similarity with TM-score = 0.5? Bioinformatics 2010, 26, 889–895. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Attwood, T.K.; Babbitt, P.C.; Bateman, A.; Bork, P.; Bridge, A.J.; Chang, H.-Y.; Dosztányi, Z.; El-Gebali, S.; Fraser, M.; et al. InterPro in 2017-beyond protein family and domain annotations. Nucleic Acids Res. 2017, 45, D190–D199. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- Kozakov, D.; Hall, D.R.; Xia, B.; Porter, K.A.; Padhorny, D.; Yueh, C.; Beglov, D.; Vajda, S. The ClusPro web server for protein-protein docking. Nat. Protoc. 2017, 12, 255–278. [Google Scholar] [CrossRef] [PubMed]
- Orchard, S.; Ammari, M.; Aranda, B.; Breuza, L.; Briganti, L.; Broackes-Carter, F.; Campbell, N.H.; Chavali, G.; Chen, C.; del-Toro, N.; et al. The MIntAct project—IntAct as a common curation platform for 11 molecular interaction databases. Nucleic Acids Res. 2014, 42, D358–D363. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Su, G.; Morris, J.H.; Demchak, B.; Bader, G.D. Biological network exploration with Cytoscape 3. Curr. Protoc. Bioinform. 2014, 47, 8.13.1–8.13.24. [Google Scholar] [CrossRef] [PubMed]
- Kotlyar, M.; Pastrello, C.; Pivetta, F.; Lo Sardo, A.; Cumbaa, C.; Li, H.; Naranian, T.; Niu, Y.; Ding, Z.; Vafaee, F.; et al. In silico prediction of physical protein interactions and characterization of interactome orphans. Nat. Methods 2015, 12, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Janky, R.; Verfaillie, A.; Imrichová, H.; Van de Sande, B.; Standaert, L.; Christiaens, V.; Hulselmans, G.; Herten, K.; Naval Sanchez, M.; Potier, D.; et al. iRegulon: From a gene list to a gene regulatory network using large motif and track collections. PLoS Comput. Biol. 2014, 10, e1003731. [Google Scholar] [CrossRef] [PubMed]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [PubMed]
- Brysbaert, G.; Lorgouilloux, K.; Vranken, W.; Lensink, M.F. RINspector: A Cytoscape app for centrality analyses and DynaMine flexibility prediction. Bioinformatics 2017. [Google Scholar] [CrossRef] [PubMed]
- Brysbaert, G.; Mauri, T.; Lensink, M.F. Comparing protein structures with RINspector automation in Cytoscape. F1000Res 2018, 7, 563. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Gautier, L.; Cope, L.; Bolstad, B.M.; Irizarry, R.A. affy—analysis of Affymetrix GeneChip data at the probe level. Bioinformatics 2004, 20, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.; Chipitsyna, G.; Cui, J.; Liao, B.; Liu, S.; Aysola, K.; Yezdani, M.; Reddy, E.S.; Rao, V.N. c-Fos oncogene regulator Elk-1 interacts with BRCA1 splice variants BRCA1a/1b and enhances BRCA1a/1b-mediated growth suppression in breast cancer cells. Oncogene 2001, 20, 1357–1367. [Google Scholar] [CrossRef] [PubMed]
- Helleday, T. The underlying mechanism for the PARP and BRCA synthetic lethality: Clearing up the misunderstandings. Mol. Oncol. 2011, 5, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Klonowska, K.; Ratajska, M.; Czubak, K.; Kuzniacka, A.; Brozek, I.; Koczkowska, M.; Sniadecki, M.; Debniak, J.; Wydra, D.; Balut, M.; et al. Analysis of large mutations in BARD1 in patients with breast and/or ovarian cancer: The Polish population as an example. Sci. Rep. 2015, 5, 10424. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Das, H.K. Inhibition of basal activity of c-jun-NH2-terminal kinase (JNK) represses the expression of presenilin-1 by a p53-dependent mechanism. Brain Res. 2008, 1207, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Günther, W.; Yoshizato, K.; Meissner, H.; Zapf, S.; Nüsing, R.M.; Yamamoto, H.; Van Meir, E.G.; Deppert, W.; Giese, A. Tumor suppressor p53 inhibits transcriptional activation of invasion gene thromboxane synthase mediated by the proto-oncogenic factor Ets-1. Oncogene 2003, 22, 7716–7727. [Google Scholar] [CrossRef] [PubMed]
- Joo, W.S.; Jeffrey, P.D.; Cantor, S.B.; Finnin, M.S.; Livingston, D.M.; Pavletich, N.P. Structure of the 53BP1 BRCT region bound to p53 and its comparison to the Brca1 BRCT structure. Genes Dev. 2002, 16, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Uzunoglu, H.; Korak, T.; Ergul, E.; Uren, N.; Sazci, A.; Utkan, N.Z.; Kargi, E.; Triyaki, Ç.; Yirmibesoglu, O. Association of the nibrin gene (NBN) variants with breast cancer. Biomed. Rep. 2016, 4, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Macías-Gómez, N.M.; Peralta-Leal, V.; Meza-Espinoza, J.P.; Gutiérrez-Angulo, M.; Durán-González, J.; Ramírez-González, J.M.; Gaspar-Del Toro, A.; Norberto-Rodríguez, A.; Leal-Ugarte, E. Polymorphisms of the XRCC1 gene and breast cancer risk in the Mexican population. Fam. Cancer 2015, 14, 349–354. [Google Scholar] [CrossRef] [PubMed]
- London, R.E. The structural basis of XRCC1-mediated DNA repair. DNA Repair (Amst) 2015, 30, 90–103. [Google Scholar] [CrossRef] [PubMed]
- Cuneo, M.J.; Gabel, S.A.; Krahn, J.M.; Ricker, M.A.; London, R.E. The structural basis for partitioning of the XRCC1/DNA ligase III-α BRCT-mediated dimer complexes. Nucleic Acids Res. 2011, 39, 7816–7827. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.-Y.; Frit, P.; Meesala, S.; Dauvillier, S.; Modesti, M.; Andres, S.N.; Huang, Y.; Sekiguchi, J.; Calsou, P.; Salles, B.; et al. Structural and functional interaction between the human DNA repair proteins DNA ligase IV and XRCC4. Mol. Cell. Biol. 2009, 29, 3163–3172. [Google Scholar] [CrossRef] [PubMed]
- Capp, J.-P.; Boudsocq, F.; Bergoglio, V.; Trouche, D.; Cazaux, C.; Blanco, L.; Hoffmann, J.-S.; Canitrot, Y. The R438W polymorphism of human DNA polymerase lambda triggers cellular sensitivity to camptothecin by compromising the homologous recombination repair pathway. Carcinogenesis 2010, 31, 1742–1747. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Wu, X. DNA polymerase lambda can elongate on DNA substrates mimicking non-homologous end joining and interact with XRCC4-ligase IV complex. Biochem. Biophys. Res. Commun. 2004, 323, 1328–1333. [Google Scholar] [CrossRef] [PubMed]
- Forma, E.; Krzeslak, A.; Bernaciak, M.; Romanowicz-Makowska, H.; Brys, M. Expression of TopBP1 in hereditary breast cancer. Mol. Biol. Rep. 2012, 39, 7795–7804. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, P.; Lin, G.E.; Liu, K.; Song, Y.; Lin, F.-T.; Lin, W.-C. Targeting TopBP1 at a convergent point of multiple oncogenic pathways for cancer therapy. Nat. Commun. 2014, 5, 5476. [Google Scholar] [CrossRef] [PubMed]
- Brenner, J.C.; Ateeq, B.; Li, Y.; Yocum, A.K.; Cao, Q.; Asangani, I.A.; Patel, S.; Wang, X.; Liang, H.; Yu, J.; et al. Mechanistic rationale for inhibition of poly(ADP-ribose) polymerase in ETS gene fusion-positive prostate cancer. Cancer Cell 2011, 19, 664–678. [Google Scholar] [CrossRef] [PubMed]
- Perez-Borrajero, C.; Lin, C.S.-H.; Okon, M.; Scheu, K.; Graves, B.J.; Murphy, M.E.P.; McIntosh, L.P. The Biophysical Basis for Phosphorylation-Enhanced DNA-Binding Autoinhibition of the ETS1 Transcription Factor. J. Mol. Biol. 2019, 431, 593–614. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.M.; Donaldson, L.W.; Pufall, M.A.; Kang, H.-S.; Pot, I.; Graves, B.J.; McIntosh, L.P. The structural and dynamic basis of Ets-1 DNA binding autoinhibition. J. Biol. Chem. 2005, 280, 7088–7099. [Google Scholar] [CrossRef] [PubMed]
- Pufall, M.A.; Lee, G.M.; Nelson, M.L.; Kang, H.-S.; Velyvis, A.; Kay, L.E.; McIntosh, L.P.; Graves, B.J. Variable control of Ets-1 DNA binding by multiple phosphates in an unstructured region. Science 2005, 309, 142–145. [Google Scholar] [CrossRef] [PubMed]
- Brysbaert, G.; Mauri, T.; de Ruyck, J.; Lensink, M.F. Identification of Key Residues in Proteins Through Centrality Analysis and Flexibility Prediction with RINspector. Curr. Protoc. Bioinform. 2019, 65, e66. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Homologs | |||
|---|---|---|---|
| Domain | Number of Genes | Number of PDB Structures | Number of Sequence Clusters |
| ETS | 42 | 23 | 38 |
| BRCT | 19 | 24 | 30 |
| SAP | 19 | 10 | 19 |
| ETS/SAP Homologs—All | ||
|---|---|---|
| Residue | Nb Interface | Nb Z-Score ≥ 2 |
| Trp361 | 10 | 10 |
| Leu342 | 10 | 5 |
| Trp338 | 10 | 5 |
| Trp356 | 10 | 3 |
| ETS/SAP homologs—DNA repair | ||
| Trp361 | 5 | 5 |
| Leu342 | 5 | 2 |
| Trp338 | 5 | 2 |
| Trp356 | 5 | 2 |
| ETS/BRCT homologs—All | ||
| Trp361 | 24 | 23 |
| Trp338 | 24 | 9 |
| Thr346 | 24 | 6 |
| Gln339 | 24 | 5 |
| Trp356 | 24 | 5 |
| Phe414 | 24 | 3 |
| Glu343 | 24 | 2 |
| ETS/BRCT homologs—DNA repair | ||
| Trp361 | 18 | 17 |
| Trp338 | 18 | 7 |
| Thr346 | 18 | 6 |
| Gln339 | 18 | 4 |
| Trp356 | 18 | 3 |
| Phe414 | 18 | 3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brysbaert, G.; de Ruyck, J.; Aumercier, M.; Lensink, M.F. Identification of Novel Interaction Partners of Ets-1: Focus on DNA Repair. Genes 2019, 10, 206. https://doi.org/10.3390/genes10030206
Brysbaert G, de Ruyck J, Aumercier M, Lensink MF. Identification of Novel Interaction Partners of Ets-1: Focus on DNA Repair. Genes. 2019; 10(3):206. https://doi.org/10.3390/genes10030206
Chicago/Turabian StyleBrysbaert, Guillaume, Jérôme de Ruyck, Marc Aumercier, and Marc F. Lensink. 2019. "Identification of Novel Interaction Partners of Ets-1: Focus on DNA Repair" Genes 10, no. 3: 206. https://doi.org/10.3390/genes10030206
APA StyleBrysbaert, G., de Ruyck, J., Aumercier, M., & Lensink, M. F. (2019). Identification of Novel Interaction Partners of Ets-1: Focus on DNA Repair. Genes, 10(3), 206. https://doi.org/10.3390/genes10030206