Impact of Chromosomal Rearrangements on the Interpretation of Lupin Karyotype Evolution

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. BAC DNA Isolation, Sequencing, and Labeling
2.3. Fluorescence In Situ Hybridization
2.4. Microscopy, Image Acquisition and Processing, Data Presentation
3. Results
3.1. Assignment of Chromosomes to Pseudomolecules of the L. angustifolius Genome
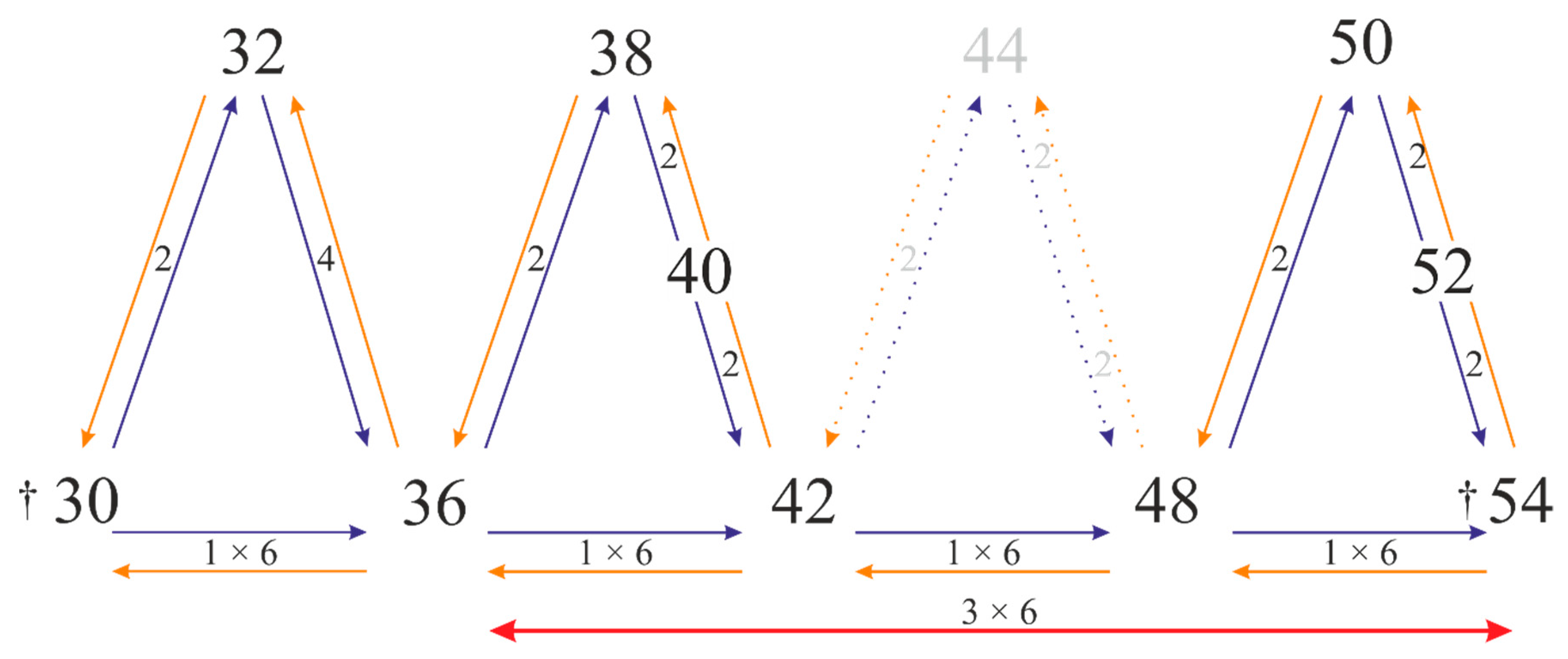
3.2. Chromosome Variation in Lupins
3.3. Chromosome Rearrangements in Lupins
4. Discussion
4.1. Lupin Chromosomes Reveal Their Puzzling Evolution
4.2. Cytogenetic Resources for Lupin Genome Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schubert, I.; Lysak, M.A. Interpretation of karyotype evolution should consider chromosome structural constraints. Trends Genet. 2011, 27, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Doležel, J.; Vrána, J.; Cápal, P.; Kubaláková, M.; Burešová, V.; Šimková, H. Advances in plant chromosome genomics. Biotechnol. Adv. 2014, 32, 122–136. [Google Scholar] [CrossRef] [PubMed]
- Iwata-Otsubo, A.; Lin, J.Y.; Gill, N.; Jackson, S.A. Highly distinct chromosomal structures in cowpea (Vigna unguiculata), as revealed by molecular cytogenetic analysis. Chromosome Res. 2016, 24, 197–216. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Choi, S.R.; Bae, J.; Hong, C.P.; Lee, S.Y.; Hossain, M.J.; Van Nguyen, D.; Jin, M.; Park, B.-S.; Bang, J.-W.; et al. Sequenced BAC anchored reference genetic map that reconciles the ten individual chromosomes of Brassica rapa. BMC Genom. 2009, 10, 432. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhang, Z.; Zong, X.; Huang, S.; Li, Z.; Han, Y. A high-resolution cucumber cytogenetic map integrated with the genome assembly. BMC Genom. 2013, 14, 461. [Google Scholar] [CrossRef]
- Chao, Y.-T.; Chen, W.-C.; Chen, C.-Y.; Ho, H.-Y.; Yeh, C.-H.; Kuo, Y.-T.; Su, C.-L.; Yen, S.-H.; Hsueh, H.-Y.; Yeh, J.-H.; et al. Chromosome-level assembly, genetic and physical mapping of Phalaenopsis aphrodite genome provides new insights into species adaptation and resources for orchid breeding. Plant Biotechnol. J. 2018, 16, 2027–2041. [Google Scholar] [CrossRef]
- Shearer, L.A.; Anderson, L.K.; de Jong, H.; Smit, S.; Goicoechea, J.L.; Roe, B.A.; Hua, A.; Giovannoni, J.J.; Stack, S.M. Fluorescence in situ hybridization and optical mapping to correct scaffold arrangement in the tomato genome. G3 (Bethesda) 2014, 4, 1395–1405. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, X.; Tian, L.; Chen, L.; Yu, W. Identification of peanut (Arachis hypogaea) chromosomes using a fluorescence in situ hybridization system reveals multiple hybridization events during tetraploid peanut formation. New Phytol. 2016, 211, 1424–1439. [Google Scholar] [CrossRef]
- Findley, S.D.; Cannon, S.; Varala, K.; Du, J.; Ma, J.; Hudson, M.E.; Birchler, J.A.; Stacey, G. A fluorescence in situ hybridization system for karyotyping soybean. Genetics 2010, 185, 727–744. [Google Scholar] [CrossRef] [PubMed]
- Fonsêca, A.; Ferreira, J.; dos Santos, T.R.B.; Mosiolek, M.; Bellucci, E.; Kami, J.; Gepts, P.; Geffroy, V.; Schweizer, D.; dos Santos, K.G.B.; et al. Cytogenetic map of common bean (Phaseolus vulgaris L.). Chromosome Res. 2010, 18, 487–502. [Google Scholar] [CrossRef]
- Fonsêca, A.; Pedrosa-Harand, A. Karyotype stability in the genus Phaseolus evidenced by the comparative mapping of the wild species Phaseolus microcarpus. Genome 2013, 56, 335–343. [Google Scholar] [CrossRef]
- Szinay, D.; Wijnker, E.; van den Berg, R.; Visser, R.G.F.; Jong, H.D.; Bai, Y. Chromosome evolution in Solanum traced by cross-species BAC-FISH. New Phytol. 2012, 195, 688–698. [Google Scholar] [CrossRef]
- Iovene, M.; Cavagnaro, P.F.; Senalik, D.; Buell, C.R.; Jiang, J.; Simon, P.W. Comparative FISH mapping of Daucus species (Apiaceae family). Chromosome Res. 2011, 19, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Gladstones, J.S. Distribution, origin, taxonomy, history and importance. In Lupins as Crop Plants: Biology, Production, and Utilization; Gladstones, J.S., Atkins, C.A., Hamblin, J., Eds.; CAB International: Wallingford, UK, 1998; pp. 1–36. [Google Scholar]
- Aïnouche, A.; Bayer, R.J.; Misset, M.T. Molecular phylogeny, diversification and character evolution in Lupinus (Fabaceae) with special attention to Mediterranean and African lupines. Plant Syst. Evol. 2004, 246, 211–222. [Google Scholar] [CrossRef]
- Naganowska, B.; Wolko, B.; Sliwinska, E.; Kaczmarek, Z. Nuclear DNA content variation and species relationships in the genus Lupinus (Fabaceae). Ann. Bot. 2003, 92, 349–355. [Google Scholar] [CrossRef]
- Susek, K.; Bielski, W.K.; Hasterok, R.; Naganowska, B.; Wolko, B. A first glimpse of wild lupin karyotype variation as revealed by comparative cytogenetic mapping. Front. Plant Sci. 2016, 7, 1152. [Google Scholar] [CrossRef] [PubMed]
- Susek, K.; Braszewska-Zalewska, A.; Bewick, A.J.; Hasterok, R.; Schmitz, R.J.; Naganowska, B. Epigenomic diversification within the genus Lupinus. PLoS ONE 2017, 12, e0179821. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.N.; Moolhuijzen, P.M.; Boersma, J.G.; Chudy, M.; Lesniewska, K.; Bellgard, M.; Oliver, R.P.; Swiecicki, W.; Wolko, B.; Cowling, W.A.; et al. Aligning a new reference genetic map of Lupinus angustifolius with the genome sequence of the model legume, Lotus japonicus. DNA Res. 2010, 17, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Hane, J.K.; Ming, Y.; Kamphuis, L.G.; Nelson, M.N.; Garg, G.; Atkins, C.A.; Bayer, P.E.; Bravo, A.; Bringans, S.; Cannon, S.; et al. A comprehensive draft genome sequence for lupin (Lupinus angustifolius), an emerging health food: Insights into plant–microbe interactions and legume evolution. Plant Biotechnol. J. 2017, 15, 318–330. [Google Scholar] [CrossRef] [PubMed]
- Książkiewicz, M.; Nazzicari, N.; Yang, H.A.; Nelson, M.N.; Renshaw, D.; Rychel, S.; Ferrari, B.; Carelli, M.; Tomaszewska, M.; Stawiński, S.; et al. A high-density consensus linkage map of white lupin highlights synteny with narrow-leafed lupin and provides markers tagging key agronomic traits. Sci. Rep. 2017, 7, 15335. [Google Scholar] [CrossRef]
- Parra-González, L.B.; Aravena-Abarzúa, G.A.; Navarro-Navarro, C.S.; Udall, J.; Maughan, J.; Peterson, L.M.; Salvo-Garrido, H.E.; Maureira-Butler, I.J. Yellow lupin (Lupinus luteus L.) transcriptome sequencing: Molecular marker development and comparative studies. BMC Genom. 2012, 13, 1–15. [Google Scholar] [CrossRef]
- Lesniewska, K.; Ksiazkiewicz, M.; Nelson, M.N.; Mahe, F.; Ainouche, A.; Wolko, B.; Naganowska, B. Assignment of 3 genetic linkage groups to 3 chromosomes of narrow-leafed lupin. J. Hered. 2011, 102, 228–236. [Google Scholar] [CrossRef]
- Wyrwa, K.; Ksiazkiewicz, M.; Szczepaniak, A.; Susek, K.; Podkowinski, J.; Naganowska, B. Integration of Lupinus angustifolius L. (narrow-leafed lupin) genome maps and comparative mapping within legumes. Chromosome Res. 2016, 24, 355–378. [Google Scholar] [CrossRef]
- Kasprzak, A.; Safar, J.; Janda, J.; Dolezel, J.; Wolko, B.; Naganowska, B. The bacterial artificial chromosome (BAC) library of the narrow-leafed lupin (Lupinus angustifolius L.). Cell. Mol. Biol. Lett. 2006, 11, 396–407. [Google Scholar] [CrossRef]
- Janečka, J.; Lysak, M.A. chromDraw: An R package for visualization of linear and circular karyotypes. Chromosome Res. 2016, 24, 217–223. [Google Scholar] [CrossRef]
- Schubert, I. Chromosome evolution. Curr. Opin. Plant Biol. 2007, 10, 109–115. [Google Scholar] [CrossRef]
- Vasconcelos, E.V.; de Andrade Fonsêca, A.F.; Pedrosa-Harand, A.; de Andrade Bortoleti, K.C.; Benko-Iseppon, A.M.; da Costa, A.F.; Brasileiro-Vidal, A.C. Intra- and interchromosomal rearrangements between cowpea [Vigna unguiculata (L.) Walp.] and common bean (Phaseolus vulgaris L.) revealed by BAC-FISH. Chromosome Res. 2015, 23, 253–266. [Google Scholar] [CrossRef]
- Wolny, E.; Lesniewska, K.; Hasterok, R.; Langdon, T. Compact genomes and complex evolution in the genus Brachypodium. Chromosoma 2011, 120, 199–212. [Google Scholar] [CrossRef]
- Lusinska, J.; Majka, J.; Betekhtin, A.; Susek, K.; Wolny, E.; Hasterok, R. Chromosome identification and reconstruction of evolutionary rearrangements in Brachypodium distachyon, B. stacei and B. hybridum. Ann. Bot. 2018, 122, 445–459. [Google Scholar] [CrossRef]
- Lysak, M.A.; Mandáková, T.; Schranz, M.E. Comparative paleogenomics of crucifers: Ancestral genomic blocks revisited. Curr. Opin. Plant Biol. 2016, 30, 108–115. [Google Scholar] [CrossRef]
- Fonsêca, A.; Ferraz, M.E.; Pedrosa-Harand, A. Speeding up chromosome evolution in Phaseolus: Multiple rearrangements associated with a one-step descending dysploidy. Chromosoma 2016, 125, 413–421. [Google Scholar] [CrossRef]
- Doyle, J.J. Polyploidy in Legumes. In Polyploidy and Genome Evolution; Soltis, P.S., Soltis, D.E., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 147–180. [Google Scholar] [CrossRef]
- Leal-Bertioli, S.C.M.; Santos, S.P.; Dantas, K.M.; Inglis, P.W.; Nielen, S.; Araujo, A.C.G.; Silva, J.P.; Cavalcante, U.; Guimarães, P.M.; Brasileiro, A.C.M.; et al. Arachis batizocoi: A study of its relationship to cultivated peanut (A. hypogaea) and its potential for introgression of wild genes into the peanut crop using induced allotetraploids. Ann. Bot. 2015, 115, 237–249. [Google Scholar] [CrossRef]
- Talukdar, D. Meiotic consequences of selfing in grass pea (Lathyrus sativus L.) autotetraploids in the advanced generations: Cytogenetics of chromosomal rearrangement and detection of aneuploids. Nucleus 2012, 55, 73–82. [Google Scholar] [CrossRef]
- Steele, K.P.; Ickert-Bond, S.M.; Zarre, S.; Wojciechowski, M.F. Phylogeny and character evolution in Medicago (Leguminosae): Evidence from analyses of plastid trnK/matK and nuclear GA3ox1 sequences. Am. J. Bot. 2010, 97, 1142–1155. [Google Scholar] [CrossRef]
- Murat, F.; Zhang, R.; Guizard, S.; Gavranovic, H.; Flores, R.; Steinbach, D.; Quesneville, H.; Tannier, E.; Salse, J. Karyotype and gene order evolution from reconstructed extinct ancestors highlight contrasts in genome plasticity of modern rosid crops. Genome Biol. Evol. 2015, 7, 735–749. [Google Scholar] [CrossRef]
- Drummond, C.S. Diversification of Lupinus (Leguminosae) in the western New World: Derived evolution of perennial life history and colonization of montane habitats. Mol. Phylogenet. Evol. 2008, 48, 408–421. [Google Scholar] [CrossRef]
- Kroc, M.; Koczyk, G.; Swiecicki, W.; Kilian, A.; Nelson, M.N. New evidence of ancestral polyploidy in the Genistoid legume Lupinus angustifolius L. (narrow-leafed lupin). Theor. Appl. Genet. 2014, 127, 1237–1249. [Google Scholar] [CrossRef]
- Conterato, I.F.; Schifino-Wittmann, M.T. New chromosome numbers, meiotic behaviour and pollen fertility in American taxa of Lupinus (Leguminosae): Contributions to taxonomic and evolutionary studies. Bot. J. Linn. Soc. 2006, 150, 229–240. [Google Scholar] [CrossRef]
- Naganowska, B.; Wolko, B.; Śliwińska, E.; Kaczmarek, Z.; Schifino-Wittmann, M. 2C DNA variation and relationships among New World species of the genus Lupinus (Fabaceae). Plant Syst. Evol. 2006, 256, 147–157. [Google Scholar] [CrossRef]
- Murat, F.; Zhang, R.; Guizard, S.; Flores, R.; Armero, A.; Pont, C.; Steinbach, D.; Quesneville, H.; Cooke, R.; Salse, J. Shared subgenome dominance following polyploidization explains grass genome evolutionary plasticity from a seven protochromosome ancestor with 16K protogenes. Genome Biol. Evol. 2014, 6, 12–33. [Google Scholar] [CrossRef]
- Cannon, S.B.; McKain, M.R.; Harkess, A.; Nelson, M.N.; Dash, S.; Deyholos, M.K.; Peng, Y.; Joyce, B.; Stewart, C.N., Jr.; Rolf, M.; et al. Multiple polyploidy events in the early radiation of nodulating and nonnodulating legumes. Mol. Biol. Evol. 2015, 32, 193–210. [Google Scholar] [CrossRef]
- Lin, J.Y.; Stupar, R.M.; Hans, C.; Hyten, D.L.; Jackson, S.A. Structural and functional divergence of a 1-Mb duplicated region in the soybean (Glycine max) genome and comparison to an orthologous region from Phaseolus vulgaris. Plant Cell 2010, 22, 2545–2561. [Google Scholar] [CrossRef] [PubMed]
- Mandáková, T.; Guo, X.; Özüdoğru, B.; Mummenhoff, K.; Lysak, M.A. Hybridization-facilitated genome merger and repeated chromosome fusion after 8 million years. Plant J. 2018, 96, 748–760. [Google Scholar] [CrossRef]
- Mandáková, T.; Kovařík, A.; Zozomová-Lihová, J.; Shimizu-Inatsugi, R.; Shimizu, K.K.; Mummenhoff, K.; Marhold, K.; Lysak, M.A. The more the merrier: Recent hybridization and polyploidy in Cardamine. Plant Cell 2013, 25, 3280–3295. [Google Scholar] [CrossRef] [PubMed]
- Drummond, C.S.; Eastwood, R.J.; Miotto, S.T.S.; Hughes, C.E. Multiple continental radiations and correlates of diversification in Lupinus (Leguminosae): Testing for key innovation with incomplete taxon sampling. Syst. Biol. 2012, 61, 443–460. [Google Scholar] [CrossRef] [PubMed]
- Mahe, F.; Markova, D.; Pasquet, R.; Misset, M.T.; Ainouche, A. Isolation, phylogeny and evolution of the SymRK gene in the legume genus Lupinus L. Mol. Phylogenet. Evol. 2011, 60, 49–61. [Google Scholar] [CrossRef]
- International Brachypodium Initiative. Genome sequencing and analysis of the model grass Brachypodium distachyon. Nature 2010, 463, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Saski, C.A.; Scheffler, B.E.; Hulse-Kemp, A.M.; Liu, B.; Song, Q.; Ando, A.; Stelly, D.M.; Scheffler, J.A.; Grimwood, J.; Jones, D.C.; et al. Sub genome anchored physical frameworks of the allotetraploid Upland cotton (Gossypium hirsutum L.) genome, and an approach toward reference-grade assemblies of polyploids. Sci. Rep. 2017, 7, 15274. [Google Scholar] [CrossRef] [PubMed]
- Boersma, J.G.; Li, C.; Leśniewska, K.; Sivasithamparam, K.; Yang, H. Identification of quantitative trait loci (QTLs) influencing early vigour, height, flowering date, and seed size and their implications for breeding of narrow-leafed lupin (Lupinus angustifolius L.). Aust. J. Agr. Res. 2008, 59, 527–535. [Google Scholar] [CrossRef]
- Yang, H.; Tao, Y.; Zheng, Z.; Zhang, Q.; Zhou, G.; Sweetingham, M.W.; Howieson, J.G.; Li, C. Draft genome sequence, and a sequence-defined genetic linkage map of the legume crop species Lupinus angustifolius L. PLoS ONE 2013, 8, e64799. [Google Scholar] [CrossRef]
- Visendi, P.; Berkman, P.J.; Hayashi, S.; Golicz, A.A.; Bayer, P.E.; Ruperao, P.; Hurgobin, B.; Montenegro, J.; Chan, C.-K.K.; Staňková, H.; et al. An efficient approach to BAC based assembly of complex genomes. Plant Methods 2016, 12, 2. [Google Scholar] [CrossRef]
- Kolmogorov, M.; Armstrong, J.; Raney, B.J.; Streeter, I.; Dunn, M.; Yang, F.; Odom, D.; Flicek, P.; Keane, T.M.; Thybert, D.; et al. Chromosome assembly of large and complex genomes using multiple references. Genome Res. 2018, 28, 1720–1732. [Google Scholar] [CrossRef]
- Danilova, T.V.; Birchler, J.A. Integrated cytogenetic map of mitotic metaphase chromosome 9 of maize: Resolution, sensitivity, and banding paint development. Chromosoma 2008, 117, 345–356. [Google Scholar] [CrossRef]
- Mandáková, T.; Lysak, M.A. Healthy roots and leaves: Comparative genome structure of horseradish and watercress. Plant Physiol. 2019, 179, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Mandakova, T.; Pouch, M.; Harmanova, K.; Zhan, S.H.; Mayrose, I.; Lysak, M.A. Multispeed genome diploidization and diversification after an ancient allopolyploidization. Mol. Ecol. 2017, 26, 6445–6462. [Google Scholar] [CrossRef] [PubMed]
- Lysak, M.A.; Berr, A.; Pecinka, A.; Schmidt, R.; McBreen, K.; Schubert, I. Mechanisms of chromosome number reduction in Arabidopsis thaliana and related Brassicaceae species. Proc. Natl. Acad. Sci. USA 2006, 103, 5224. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Zhang, T.; Thammapichai, P.; Weng, Y.; Jiang, J. Chromosome-specific painting in Cucumis species using bulked oligonucleotides. Genetics 2015, 200, 771–779. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Section | Species | Accession | Chromosome Number (2n) | Genome Size (pg/2C DNA) |
|---|---|---|---|---|---|
| Smooth-seeded | Angustifolius | L. angustifolius | cv. ‘Sonet’ * | 40 | 1.89 |
| L. cryptanthus | 96361 * | 40 | 1.86 | ||
| Luteus | L. luteus | cv. ‘Talar’ * | 52 | 2.44 | |
| L. hispanicus | 96385 * | 52 | 2.15 | ||
| Albus | L. albus | cv. ‘Boros’ * | 50 | 1.16 | |
| L. graecus | 95601 * | 50 | 1.13 | ||
| Micranthus | L. micranthus | 98552 * | 52 | 0.98 | |
| Rough-seeded | Atlanticus | L. atlanticus | 98401 * | 38 | 1.61 |
| L. digitatus | PI 660697 ** | 36 | 1.37 | ||
| L. cosentinii | 98452 * | 32 | 1.42 | ||
| Pilosus | L. pilosus | 98653 * | 42 | 1.36 | |
| L. palaestinus | 98605 * | 42 | 1.39 |
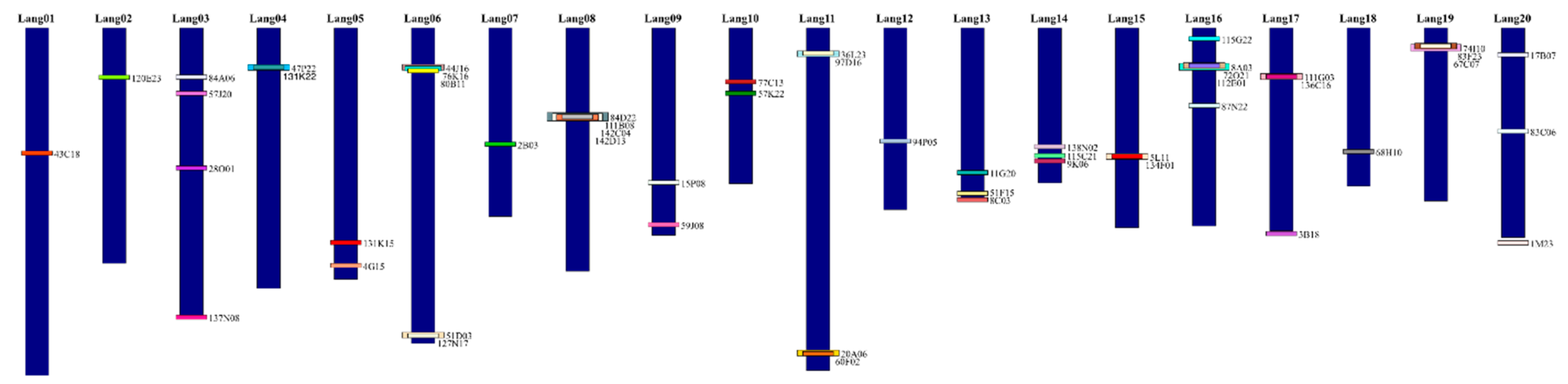
| Assignment | Chromosome (Lang) | Pseudomolecule (NLL) | Pseudomolecule (bp) | BAC ID | BAC Data | GenBank Accession |
|---|---|---|---|---|---|---|
| Lang01 | 01 | 01 | 1..36 457 581 | 43C18 | WBS | MK650088 |
| 13 | ||||||
| Lang02 | 02 | 02 | 1..24 697 652 | 120E23 | WBS | MK650090 |
| Lang03 | 03 | 03 | 1..30 153 019 | 84A06 | WBS | MK650076 |
| 03 | 03 | 57J20 | WBS | HE804810.1 | ||
| 03 | 03 | 28O01 | 3′ STS | GF112056.1 | ||
| 03 | 03 | 137N08 | WBS | MK650084 | ||
| Lang04 | 04 | 04 | 1..27 333 975 | 47P22 | WBS | MK650072 |
| 04 | 04 | 131K22 | 5′ BES | KU678257.1 | ||
| 04 | 04 | 3′ BES | KU678256.1 | |||
| Lang05 | 05 | 05 | 1..26376911 | 131K15 | WBS | MK650081 |
| 05 | 05 | 4G15 | WBS | HE804808.1 | ||
| Lang06 | 06 | 06 | 1..33 111 450 | 44J16 | WBS | KX298066 |
| 06 | 06 | 76K16 | WBS | MK650073 | ||
| 06 | 06 | 80B11 | WBS | HE804812.1 | ||
| 06 | 06 | 51D03 | WBS | MK650071 | ||
| 06 | 06 | 127N17 | WBS | KU678223.1 | ||
| Lang07 | Not assigned | 07 | 1..19 782 170 | 2B03 | WBS | KX298069 |
| Lang08 | 08 | 17 | 1...25 521 646 | 84D22 | WBS | KX298065 |
| 08 | 17 | 111B08 | WBS | KX298071 | ||
| 08 | 17 | 142C04 | WBS | KX298073 | ||
| 08 | 17 | 142D13 | WBS | KX298074 | ||
| Lang09 | 09 | 09 | 1..21 753 712 | 15P08 | 5′ BES | AB809174.1 |
| 09 | 3′ BES | AB809173.1 | ||||
| 09 | 09 | 59J08 | WBS | MK650082 | ||
| Lang10 | 10 | 10 | 1...16 341 955 | 77C13 | WBS | MK650077 |
| 10 | 10 | 57K22 | WBS | MK650069 | ||
| Lang11 | 11 | 11 | 1..35 963 958 | 97D16 | WBS | MK650085 |
| 11 | 11 | 36L23 | WBS | MK650075 | ||
| 11 | 11 | 20A06 | WBS | MK650080 | ||
| 11 | 11 | 60F02 | WBS | MK045265 | ||
| Lang12 | 12 | 12 | 1..19 065 701 | 94P05 | WBS | MK650086 |
| Lang13 | Not analyzed | Many | 1..17 820 680 | 11G20 | WBS | MK650079 |
| Not assigned | 13 | 8C03 | WBS | KX298063 | ||
| Not analyzed | 13 | 51F15 | WBS | MK045272 | ||
| Lang14 | 14 | 03 | 1...16 251 777 | 138N02 | WBS | MK650089 |
| 14 | 14 | 115C21 | 5′ BES | HR864196.1 | ||
| 12 | 3′ BES | AB809318.1 | ||||
| Not analyzed | 14 | 9K06 | WBS | MK045273 | ||
| Lang15 | 15 | 15 | 1...20 964 703 | 5L11 | 3′ STS | GF112057.1 |
| 15 | 134F01 | WBS | MK650070 | |||
| Lang16 | 16 | Many | 1..20 786 881 | 115G22 | 5′ BES | AB811328.1 |
| Many | 3′ BES | AB811323.1 | ||||
| 16 | Many | 112E01 | 5′ BES | AB809308.1 | ||
| Many | 3′ BES | AB809307.1 | ||||
| 16 | Many | 72O21 | 5′ BES | AB809272.1 | ||
| 16 | 20 | 8A03 | 5′ BES | AB809167.1 | ||
| Many | 3′ BES | AB809166.1 | ||||
| 16 | 16 | 87N22 | WBS | MK650087 | ||
| Lang17 | 17 | 17 | 1..21 299 880 | 111G03 | WBS | KX298064 |
| 17 | 17 | 136C16 | WBS | KX298072 | ||
| 17 | 17 | 3B18 | WBS | KX298070 | ||
| Lang18 | 18 | 18 | 1..16 588 007 | 68H10 | WBS | KU678221 |
| Lang19 | 19 | 19 | 1..18 159 812 | 67C07 | WBS | MK650078 |
| 19 | 19 | 83F23 | 5′ BES | KU678304.1 | ||
| 19 | 3′ BES | KU678303.1 | ||||
| 07 | 19 | 74I10 | WBS | MK650074 | ||
| Lang20 | 20 | 20 | 1..21 988 590 | 17B07 | WBS | HF937076.1 |
| 20 | 20 | 83C06 | WBS | MK650083 | ||
| Cluster-2 | 20 | 1M23 | WBS | KX298067 |
| No. | BAC ID | Lcry | Llut | Lhis | Lalb | Lgra | Lmic | Latl | Ldig | Lcos | Lpil | Lpal |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lang01 | 1-R | 1-R | 1-R | 1-R | 1-R | 1-R | 1-R | 1-R | 1-R | 1-R | 1-R | |
| 1 | 43C18 | R | R | R | R | R | R | R | R | R | R | R |
| Lang02 | S | S | S | 2-R | 2-R | S | S | 2-R | S | S | S | |
| 2 | 120E23 | S | S | S | R | R | S | S | R | S | S | S |
| Lang03 | 3=4=6; 5-R | 3-4-5-6-R | 3-4-5-6-R | 3=4=6; 5-R | 3=4; 5-6-R | 3=6; 4-5-R | 3-4-5-6-R | 3-5-6-R | 3=6; 4-5-R | 3=6; 4-5-R | 3=4=6; 5-R | |
| 3 | 84A06 | S | R | R | S | S | S | R | R | S | S | S |
| 4 | 57J20 | S | R | R | S | S | R | R | N/D | R | R | S |
| 5 | 28O01 | R | R | R | R | R | R | R | R | R | R | R |
| 6 | 137N08 | S | R | R | S | R | S | R | R | S | S | S |
| Lang04 | 7=8 | 7-8-R | 7=8 | 7; 8-R | 7=8 | 7; 8-R | 7-8-R | 7-8-R | 7; 8-R | 7; 8-R | 7=8 | |
| 7 | 47P22 | S | R | S | S | S | S | R | R | S | S | S |
| 8 | 131K22 | S | R | S | R | S | R | R | R | R | R | S |
| Lang05 | 9=10 | 9=10 | 9-10-R | 9=10 | 9=10 | 9=10 | 9=10 | 9=10 | 9=10 | 9=10 | 9=10 | |
| 9 | 131K15 | S | S | R | S | S | S | S | S | S | S | S |
| 10 | 4G15 | S | S | R | S | S | S | S | S | S | S | S |
| Lang06 | 11=12=13=15=16 | 11=13 #14; 12-15-R | 13; 11-12-14-15-R | 12=13 # 14; 11-15-R | 12=13 # 14; 11-15-R | 12=13 # 14=15 | 11=12=13=15; 14-R | 12=13; 11-14-15-R | 13; 12-14-15-R | 12 # 13 # 14=15 | 12=13#12′; 11-14-15-R | |
| 11 | 44J16 | S | S | R | R | R | N/D | S | R | N/D | N/D | R |
| 12 | 76K16 | S | R | R | S | S | S | S | S | R | S | S, S′ |
| 13 | 80B11 | S | S | S | S | S | S | S | S | S | S | S |
| 14 | 51D03 | S | S | R | S | S | S | R | R | R | S | R |
| 15 | 127N17 | S | R | R | R | N/D | S | S | R | R | S | R |
| Lang07 | S | S | S | S | S | 16-R | S | 16-R | S | S | S | |
| 16 | 2B03 | S | S | S | S | S | R | S | R | S | S | S |
| Lang08 | 17=18=19=20 | 18=20; 17-19-R | 17-18-19-20-R | 17-18-19-20-R | 17-18-19-20-R | 17=19 # 18; 20-R | 17-18-19-20-R | 17-18-19-20-R | 17=18; 19-20-R | 17=18; 19-20-R | 17-18-19-20-R | |
| 17 | 84D22 | S | R | R | R | R | S | R | R | S | S | R |
| 18 | 111B08 | S | S | R | R | R | S | R | R | S | S | R |
| 19 | 142C04 | S | R | R | R | R | S | R | R | R | R | R |
| 20 | 142D13 | S | S | R | R | R | R | R | R | R | R | R |
| Lang09 | 22; 21-R | 22; 21-R | 22; 21-R | 22; 21-R | 22; 21-R | 22; 21-R | 22; 21-R | 22; 21-R | 22; 21-R | 22; 21-R | 22; 21-R | |
| 21 | 15P08 | R | R | R | R | R | R | R | R | R | R | R |
| 22 | 59J08 | S | S | S | S | S | S | S | S | S | S | S |
| Lang10 | 23=24 | 23; 24-R | 23-24-R | 23; 24-R | 23-24-R | 23=24 | 23; 24-R | 23; 24-R | 23-24-R | 23-24-R | 23-24-R | |
| 23 | 77C13 | S | S | R | S | R | S | S | S | R | R | R |
| 24 | 57K22 | S | R | R | R | R | S | R | R | R | R | R |
| Lang11 | 25=26=27=28 | 26; 25-27-28-R | 25=26; 27-28-R | 25=27; 26-28-R | 25=26; 27-28-R | 26; 25-27-28-R | 25; 26-27-28-R | 26-27-28-R | 26; 25-27-28-R | 26; 25-27-28-R | 25; 27-28-R | |
| 25 | 97D16 | S | R | S | S | S | R | S | N/D | R | R | S |
| 26 | 36L23 | S | S | S | R | S | S | R | R | S | S | N/D |
| 27 | 20A06 | S | R | R | S | R | R | R | R | R | R | R |
| 28 | 60F02 | S | R | R | R | R | R | R | R | R | R | R |
| Lang12 | 29-R | S | 29-R | S | S | 29-R | 29-R | 29-R | 29-R | 29-R | S | |
| 29 | 94P05 | R | S | R | S | S | R | R | R | R | R | S |
| Lang13 | 31=32; 30-R | 32 # 32′; 30-31-R | 32; 30-31-R | 32; 30-31-R | 32; 30-31-R | 31; 30-32-R | 32; 30-31-R | 32; 30-31-R | 32; 30-31-R | 32; 30-31-R | 30-31-32-R | |
| 30 | 11G20 | R | R | R | R | R | R | R | R | R | R | R |
| 31 | 8C03 | S | R | R | R | R | S | R | N/D | R | R | R |
| 32 | 51F15 | S | S, S′ | S | S | S | R | S | S | S | S | R |
| Lang14 | 33=35; 34-R | 33-34-35-R | 33-34-35-R | 33-34-35-R | 35; 33-34-R | 35; 33-34-R | 35; 33-34-R | 33-34-35-R | 35; 33-34-R | 35; 33-34-R | 33-34-35-R | |
| 33 | 138N02 | S | R | R | R | R | R | R | R | R | R | R |
| 34 | 115C21 | R | R | R | R | R | R | R | R | R | R | R |
| 35 | 9K06 | S | R | R | R | S | S | S | R | S | S | R |
| Lang15 | 37; 36-R | 36-37-R | 36-37-R | 36-37-R | 36-37-R | 37; 36-R | 36-37-R | 37; 36-R | 37; 36-R | 37; 36-R | 36-37-R | |
| 36 | 5L11 | R | R | R | R | R | R | R | R | R | R | R |
| 37 | 134F01 | S | R | R | R | R | S | R | S | S | S | R |
| Lang16 | 38=39=40=41; 42-R | 38=39=40=41; 42-R | 38=39=40=41; 42-R | 38=39=40=41; 42-R | 38=39=40=41; 42-R | 38=39=40=41; 42-R | 38=39=40=41; 42-R | 38=39=40=41; 42-R | 38=39=40=41; 42-R | 38=39=40=41; 42-R | 38=39=40=41; 42-R | |
| 38 | 115G22 | S | S | S | S | S | S | S | S | S | S | S |
| 39 | 112E01 | S | S | S | S | S | S | S | S | S | S | S |
| 40 | 72O21 | S | S | S | S | S | S | S | S | S | S | S |
| 41 | 8A03 | S | S | S | S | S | S | S | S | S | S | S |
| 42 | 87N22 | R | R | R | R | R | R | R | R | R | R | R |
| Lang17 | 43=45; 44-R | 43=44=45 | 43=44=45 | 43=44=45 | 43=44=45 | 43=44=45 | 43=44=45 | 43 # 44=45 # 45′ | 43=44=45 | 43 # 44=45 | 44=45; 43-R | |
| 43 | 3B18 | S | S | S | S | S | S | S | S | S | S | R |
| 44 | 111G03 | R | S | S | S | S | S | S | S | S | S | S |
| 45 | 136C16 | S | S | S | S | S | S | S | S, S′ | S | S | S |
| Lang18 | 46-R | 46-R | 46-R | 46-R | 46-R | 46-R | 46-R | 46-R | 46-R | 46-R | 46-R | |
| 46 | 68H10 | R | R | R | R | R | R | R | R | R | R | R |
| Lang19 | 47=48; 49-R | 47; 48-49-R | 47=48; 49-R | 47-49-R | 47-48-49-R | 47=48; 49-R | 47=48=49 | 47=48; 49-R | 47=48; 49-R | 47=48; 49-R | 47=48; 49-R | |
| 47 | 67C07 | S | S | S | R | R | S | S | S | S | S | S |
| 48 | 83F23 | S | R | S | N/D | R | S | S | S | S | S | S |
| 49 | 74I10 | R | R | R | R | R | R | S | R | R | R | R |
| Lang20 | 50=51=52 | 50=51=52 | 50; 51-52-R | 50=51; 52-R | 50; 51-52-R | 50=51; 52-R | 50=51; 52-R | 50=51; 52-R | 50=51; 52-R | 50=51; 52-R | 50=51; 52-R | |
| 50 | 17B07 | S | S | S | S | S | S | S | S | S | S | S |
| 51 | 83C06 | S | S | R | S | R | S | S | S | S | S | S |
| 52 | 1M23 | S | S | R | R | R | R | R | R | R | R | R |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Susek, K.; Bielski, W.; Czyż, K.B.; Hasterok, R.; Jackson, S.A.; Wolko, B.; Naganowska, B. Impact of Chromosomal Rearrangements on the Interpretation of Lupin Karyotype Evolution. Genes 2019, 10, 259. https://doi.org/10.3390/genes10040259
Susek K, Bielski W, Czyż KB, Hasterok R, Jackson SA, Wolko B, Naganowska B. Impact of Chromosomal Rearrangements on the Interpretation of Lupin Karyotype Evolution. Genes. 2019; 10(4):259. https://doi.org/10.3390/genes10040259
Chicago/Turabian StyleSusek, Karolina, Wojciech Bielski, Katarzyna B. Czyż, Robert Hasterok, Scott A. Jackson, Bogdan Wolko, and Barbara Naganowska. 2019. "Impact of Chromosomal Rearrangements on the Interpretation of Lupin Karyotype Evolution" Genes 10, no. 4: 259. https://doi.org/10.3390/genes10040259
APA StyleSusek, K., Bielski, W., Czyż, K. B., Hasterok, R., Jackson, S. A., Wolko, B., & Naganowska, B. (2019). Impact of Chromosomal Rearrangements on the Interpretation of Lupin Karyotype Evolution. Genes, 10(4), 259. https://doi.org/10.3390/genes10040259