A Premature Stop Codon in RAF1 Is the Priority Candidate Causative Mutation of the Inherited Chicken Wingless-2 Developmental Syndrome


Abstract
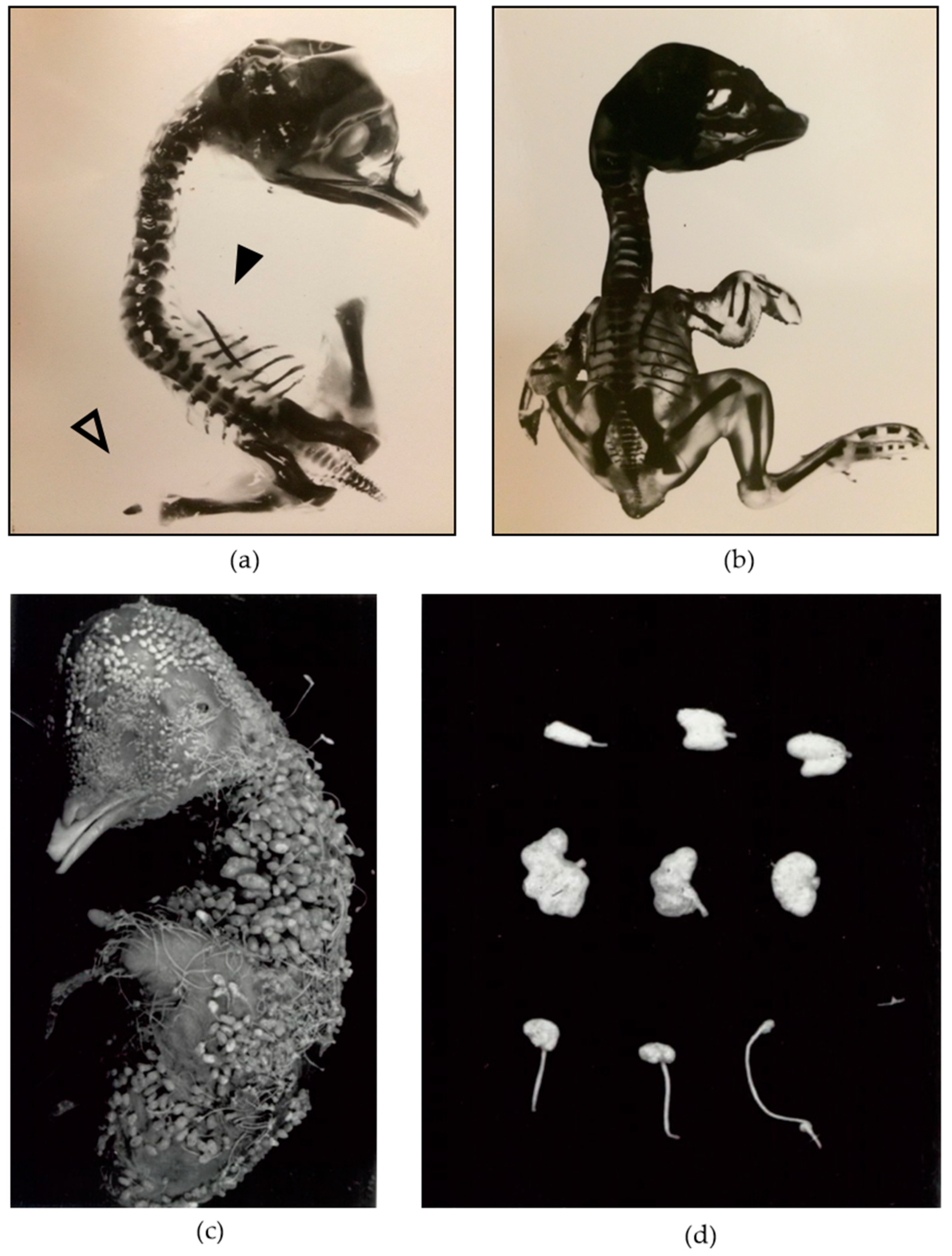
:1. Introduction
2. Materials and Methods
2.1. Genetic Lines
2.2. Capture Array and Next-Generation Sequencing
2.2.1. Sample Collection
2.2.2. Capture Array and Sequencing
2.2.3. Bioinformatic Analysis
2.3. RT-qPCR
2.3.1. Sample Collection
2.3.2. RNA Isolation
2.3.3. cDNA Reverse Transcription
2.3.4. TaqMan Gene Expression Assays
2.3.5. Quantitative Analysis
2.4. Western blotting
2.4.1. Sample Collection
2.4.2. Protein Extraction
2.4.3. SDS-PAGE and Protein Transfer
2.4.4. Western Blotting
3. Results
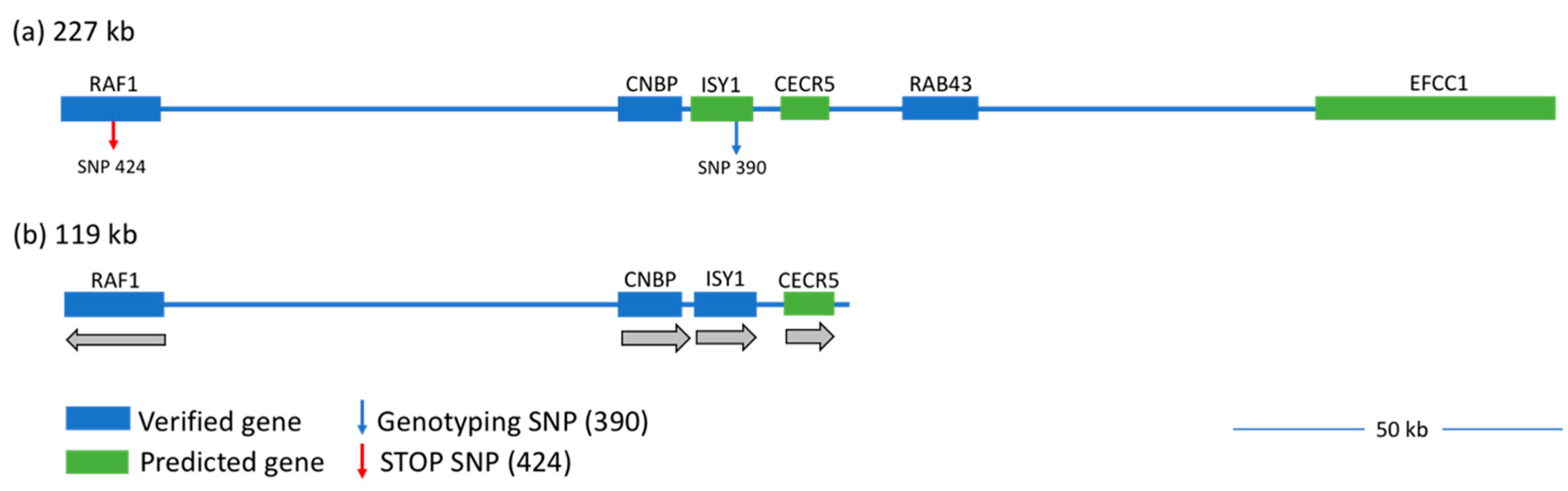
3.1. Capture Array and Next Generation Sequencing
3.2. RT-qPCR
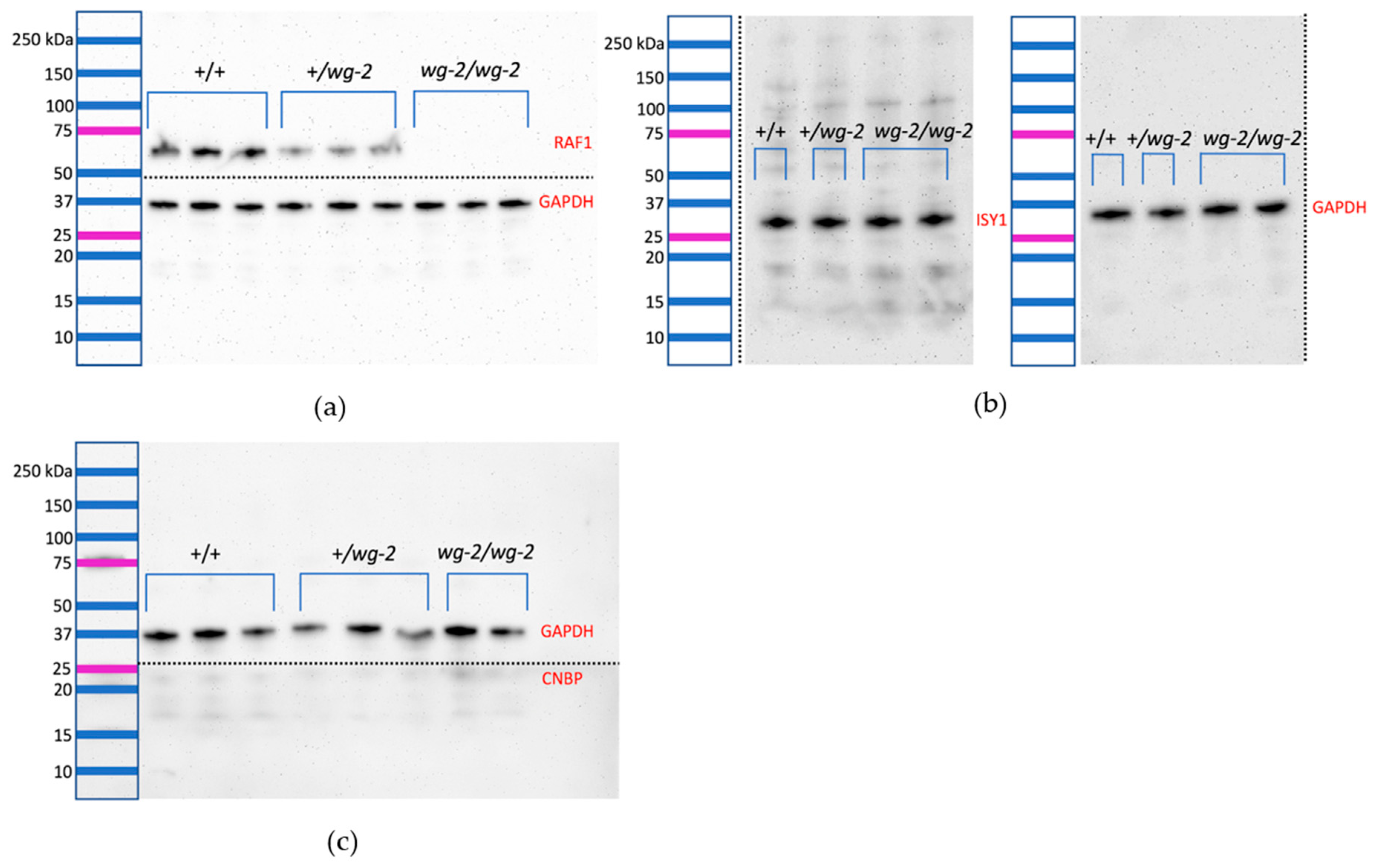
3.3. Western blotting
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Davey, M.G.; Towers, M.; Vargesson, N.; Tickle, C. The chick limb: Embryology, genetics and teratology. Int. J. Dev. Biol. 2018, 62, 85–95. [Google Scholar] [CrossRef]
- Carrington, J.L.; Fallon, J.F. Evidence that the ectoderm is the affected germ layer in the wingless mutant chick embryo. J. Exp. Zool. 1984, 232, 297–308. [Google Scholar] [CrossRef]
- Zwilling, E. Interaction between limb bud ectoderm and mesoderm in the chick embryo. II. Experimental limb duplication. J. Exp. Zool. 1956, 132, 173–187. [Google Scholar] [CrossRef]
- Pisenti, J.M.; Delany, M.E.; Taylor, R.L.; Abbott, U.K.; Abplanalp, H.; Arthur, J.A.; Bakst, M.R.; Baxter-Jones, C.; Bitgood, J.J.; Bradley, F.A.; et al. Avian genetic resources at risk: An assessment and proposal for conservation of genetic stocks in the USA and Canada. Avian Poult. Biol. Rev. 1999, 12, i–xiii, 1–102. Available online: http://grcp.ucdavis.edu/publications/index.htm (accessed on 29 April 2019).
- Webb, A.E.; Youngworth, I.A.; Kaya, M.; Gitter, C.L.; O’Hare, E.A.; May, B.; Cheng, H.H.; Delany, M.E. Narrowing the wingless-2 mutation to a 227 kb candidate region on chicken chromosome 12. Poult. Sci. 2018, 97, 1872–1880. [Google Scholar] [CrossRef] [PubMed]
- Robb, E.A.; Gitter, C.L.; Cheng, H.H.; Delany, M.E. Chromosomal mapping and candidate gene discovery of chicken developmental mutants and genome-wide variation analysis of MHC congenics. J. Hered. 2011, 102, 141–156. [Google Scholar] [CrossRef]
- Pisenti, J.M. Genetic and Environmental Influences on the Expression of the Mutation wingless-2 in the Chick Embryo. Ph.D. Thesis, University California, Davis, CA, USA, 1995. [Google Scholar]
- Robb, E.A.; Delany, M.E. Case study of sequence capture enrichment technology: Identification of variation underpinning developmental syndromes in an amniote model. Genes 2012, 3, 233–247. [Google Scholar] [CrossRef]
- Kent, W.J.; Sugnet, C.W.; Furey, T.S.; Roskin, K.M.; Pringle, T.H.; Zahler, A.M.; Haussler, D. The human genome browser at UCSC. Genome Res. 2002, 12, 996–1006. [Google Scholar] [CrossRef]
- Baccarini, M. Second nature: Biological functions of the Raf-1 “kinase”. FEBS Lett. 2005, 579, 3271–3277. [Google Scholar] [CrossRef] [Green Version]
- Galabova-Kovacs, G.; Matzen, D.; Piazzolla, D.; Meissl, K.; Plyushch, T.; Chen, A.P.; Silva, A.; Baccarini, M. Essential role of B-Raf in ERK activation during extraembryonic development. Proc. Natl. Acad. Sci. USA 2006, 103, 1325–1330. [Google Scholar] [CrossRef] [Green Version]
- Matallanas, D.; Birtwistle, M.; Romano, D.; Zebisch, A.; Rauch, J.; von Kriegsheim, A.; Kolch, W. Raf family kinases: Old dogs have learned new tricks. Genes Cancer. 2011, 2, 232–260. [Google Scholar] [CrossRef]
- Molina, J.R.; Adjei, A.A. The Ras/Raf/MAPK pathway. J. Thorac. Oncol. 2006, 1, 7–9. [Google Scholar] [CrossRef] [PubMed]
- Abe, Y.; Chen, W.; Huang, W.; Nishino, M.; Li, Y.P. CNBP regulates forebrain formation at organogenesis stage in chick embryos. Dev. Biol. 2006, 295, 116–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Liang, Y.; Deng, W.; Shimizu, K.; Ashique, A.M.; Li, E.; Li, Y.P. The zinc-finger protein CNBP is required for forebrain formation in the mouse. Development 2003, 130, 1367–1379. [Google Scholar] [CrossRef] [Green Version]
- Flink, I.L.; Morkin, E. Organization of the gene encoding cellular nucleic acid-binding protein. Gene 1995, 163, 279–282. [Google Scholar] [CrossRef]
- Michelotti, E.F.; Tomonaga, T.; Krutzsch, H.; Levens, D. Cellular nucleic acid binding protein regulates the CT element of the human c-myc protooncogene. J. Biol. Chem. 1995, 270, 9494–9499. [Google Scholar] [CrossRef]
- Dejgaard, S.Y.; Murshid, A.; Erman, A.; Kizilay, O.; Verbich, D.; Lodge, R.; Dejgaard, K.; Ly-Hartig, T.B.N.; Pepperkok, R.; Simpson, J.C.; et al. Rab18 and Rab43 have key roles in ER-Golgi trafficking. J. Cell Sci. 2008, 121, 2768–2781. [Google Scholar] [CrossRef] [Green Version]
- Haas, A.K.; Yoshimura, S.-I.; Stephens, D.J.; Preisinger, C.; Fuchs, E.; Barr, F.A. Analysis of GTPase-activating proteins: Rab1 and Rab43 are key Rabs required to maintain a functional Golgi complex in human cells. J. Cell Sci. 2007, 120, 2997–3010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dix, I.; Russell, C.; Yehuda, S.B.; Kupiec, M.; Beggs, J.D. The identification and characterization of a novel splicing protein, Isy1p, of Saccharomyces cerevisiae. RNA 1999, 5, 360–368. [Google Scholar] [CrossRef] [Green Version]
- Villa, T. The Isy1p component of the NineTeen Complex interacts with the ATPase Prp16p to regulate the fidelity of pre-mRNA splicing. Genes Dev. 2005, 19, 1894–1904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- UniProtKB – Q9BXW7 (HDHD5_HUMAN). Available online: https://www.uniprot.org/uniprot/Q9BXW7 (accessed on 17 January 2019).
- UniProtKB – Q9HA90 (EFCC1_HUMAN). Available online: https://www.uniprot.org/uniprot/Q9HA90 (accessed on 17 January 2019).
- Rauen, K.A. The RASopathies. Annu. Rev. Genom. Hum. Genet. 2013, 14, 355–369. [Google Scholar] [CrossRef] [Green Version]
- Corson, L.B.; Yamanaka, Y.; Lai, K.-M.V.; Rossant, J. Spatial and temporal patterns of ERK signaling during mouse embryogenesis. Development 2003, 130, 4527–4537. [Google Scholar] [CrossRef] [Green Version]
- Hamburger, V.; Hamilton, H.L. A series of normal stages in the development of the chick embryo. Dev. Dyn. 1992, 195, 231–272. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Picard. Broad Institute, Version 2.8.0. Available online: http://broadinstitute.github.io/picard/ (accessed on 7 July 2017).
- Koboldt, D.C.; Zhang, Q.; Larson, D.E.; Shen, D.; McLellan, M.D.; Lin, L.; Miller, C.A.; Mardis, E.R.; Ding, L.; Wilson, R.K. VarScan 2: Somatic mutation and copy number alteration discovery in cancer by exome sequencing. Genome Res. 2012, 22, 568–576. [Google Scholar] [CrossRef] [Green Version]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w 1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bland, J.M.; Altman, D.G. Statistics notes: Multiple significance tests: The Bonferroni method. BMJ 1995, 310, 170. [Google Scholar] [CrossRef]
- Aken, B.L.; Ayling, S.; Barrell, D.; Clarke, L.; Curwen, V.; Fairley, S.; Fernandez Banet, J.; Billis, K.; García Girón, C.; Hourlier, T.; et al. The Ensembl gene annotation system. Database 2016, 2016, baw093. [Google Scholar] [CrossRef]
- UniProtKB – P05625 (RAF1_CHICK). Available online: https://www.uniprot.org/uniprot/P05625 (accessed on 17 January 2019).
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef]
- Karousis, E.D.; Nasif, S.; Mühlemann, O. Nonsense-mediated mRNA decay: Novel mechanistic insights and biological impact: Nonsense-mediated mRNA decay. Wiley Interdiscip. Rev. RNA 2016, 7, 661–682. [Google Scholar] [CrossRef]
- Nickless, A.; Bailis, J.M.; You, Z. Control of gene expression through the nonsense-mediated RNA decay pathway. Cell Biosci. 2017, 7. [Google Scholar] [CrossRef]
- Peltz, S.W.; Brown, A.H.; Jacobson, A. mRNA destabilization triggered by premature translational termination depends on at least three cis-acting sequence elements and one trans-acting factor. Genes Dev. 1993, 7, 1737–1754. [Google Scholar] [CrossRef]
- Kuo, R.I.; Tseng, E.; Eory, L.; Paton, I.R.; Archibald, A.L.; Burt, D.W. Normalized long read RNA sequencing in chicken reveals transcriptome complexity similar to human. BMC Genom. 2017, 18. [Google Scholar] [CrossRef]
- Storm, S.M.; Cleveland, J.L.; Rapp, U.R. Expression of raf family proto-oncogenes in normal mouse tissues. Oncogene 1990, 5, 345–351. [Google Scholar]
- Tidyman, W.E.; Rauen, K.A. Pathogenetics of the RASopathies. Hum. Mol. Genet. 2016, 25, R123–R132. [Google Scholar] [CrossRef]
- Tajan, M.; Paccoud, R.; Branka, S.; Edouard, T.; Yart, A. The RASopathy Family: Consequences of Germline Activation of the RAS/MAPK Pathway. Endocr. Rev. 2018, 39, 676–700. [Google Scholar] [CrossRef]
- Lavoie, H.; Therrien, M. Regulation of RAF protein kinases in ERK signalling. Nat. Rev. Mol. Cell Biol. 2015, 16, 281–298. [Google Scholar] [CrossRef]
- Wellbrock, C.; Karasarides, M.; Marais, R. The RAF proteins take centre stage. Nat. Rev. Mol. Cell Biol. 2004, 5, 875–885. [Google Scholar] [CrossRef] [Green Version]
- Brewer, J.R.; Mazot, P.; Soriano, P. Genetic insights into the mechanisms of Fgf signaling. Genes Dev. 2016, 30, 751–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gros, J.; Hu, J.K.-H.; Vinegoni, C.; Feruglio, P.F.; Weissleder, R.; Tabin, C.J. WNT5A/JNK and FGF/MAPK Pathways Regulate the Cellular Events Shaping the Vertebrate Limb Bud. Curr. Biol. 2010, 20, 1993–2002. [Google Scholar] [CrossRef]
- Eblaghie, M.C.; Lunn, J.S.; Dickinson, R.J.; Münsterberg, A.E.; Sanz-Ezquerro, J.-J.; Farrell, E.R.; Mathers, J.; Keyse, S.M.; Storey, K.; Tickle, C. Negative Feedback Regulation of FGF Signaling Levels by Pyst1/MKP3 in Chick Embryos. Curr. Biol. 2003, 13, 1009–1018. [Google Scholar] [CrossRef] [Green Version]
- Abbott, U.K.; Pisenti, J.M. Making the connection: Exploring classical concepts in normal and abnormal limb development using contemporary approaches. Prog. Clin. Biol. Res. 1993, 383A, 99–112. [Google Scholar]
- Wojnowski, L.; Stancato, L.F.; Zimmer, A.M.; Hahn, H.; Beck, T.W.; Larner, A.C.; de Rapp, U.R.; Zimmer, A. Craf-1 protein kinase is essential for mouse development. Mech. Dev. 1998, 76, 141–149. [Google Scholar] [CrossRef]
- Mikula, M. Embryonic lethality and fetal liver apoptosis in mice lacking the c-raf-1 gene. EMBO J. 2001, 20, 1952–1962. [Google Scholar] [CrossRef] [Green Version]
- Huser, M. MEK kinase activity is not necessary for Raf-1 function. EMBO J. 2001, 20, 1940–1951. [Google Scholar] [CrossRef] [Green Version]
- Wojnowski, L.; Stancato, L.F.; Larner, A.C.; Rapp, U.R.; Zimmer, A. Overlapping and specific functions of Braf and Craf-1 proto-oncogenes during mouse embryogenesis. Mech. Dev. 2000, 91, 97–104. [Google Scholar] [CrossRef]
- Pearson, G.; Bumeister, R.; Henry, D.O.; Cobb, M.H.; White, M.A. Uncoupling Raf1 from MEK1/2 Impairs Only a Subset of Cellular Responses to Raf Activation. J. Biol. Chem. 2000, 275, 37303–37306. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Gene Target | Assay ID | Genome Build | Amplicon Size (Bases) | Dye |
|---|---|---|---|---|
| RAF1 | Gg03349021_m1 | galGal2 | 127 | FAM-MGB 1 |
| CNBP | Gg03339297_m1 | galGal2 | 114 | FAM-MGB 1 |
| ISY1 | Gg03328872_m1 | galGal2 | 71 | FAM-MGB 1 |
| RAB43 | Hs03006628_gH | GRCh38 2 | 59 | FAM-MGB 1 |
| CECR5 | Hs00215190_m1 | GRCh38 2 | 68 | FAM-MGB 1 |
| EFCC1 | Hs01088833_m1 | GRCh38 2 | 99 | FAM-MGB 1 |
| Protein Target 1 | Company | ID | Host Species 1 | Clonality | Dilution |
|---|---|---|---|---|---|
| RAF1 | Abcam | ab181115 | Rabbit | monoclonal | 1:1000 |
| CNBP | Abcam | ab83038 | Rabbit | polyclonal | 1:150 |
| ISY1 | Novus Biologicals | NBP1-81865 | Rabbit | polyclonal | 1:250 |
| GAPDH 2 | Novus Biologicals | NB300-322 | Rabbit | polyclonal | 1:2000 |
| Rabbit IgG 3 | Abcam | ab205718 | Goat | polyclonal | 1:50,000 |
| 227-kb Linked Variants 1 | |||
|---|---|---|---|
| Variant | Non-Coding | Splice Region | Exon |
| SNVs | 503 | 4 | 9 |
| Indels | 53 | 0 | 0 |
| Fold Changes in Gene Expression | ||||||
|---|---|---|---|---|---|---|
| Gene | wg-2/wg-2:+/+ | p-Value | wg-2/wg-2:+/wg-2 | p-Value | +/wg-2:+/+ | p-Value |
| RAF1 | 1.7 ** | 0.001 | 1.2 | 0.22 | 1.4 * | 0.03 |
| ISY1 | 1.0 | 1.159 | 1.1 | 0.09 | 0.9 | 0.11 |
| CNBP | 1.1 | 1.383 | 1.1 | 1.80 | 1.1 | 9.64 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Youngworth, I.; Delany, M.E. A Premature Stop Codon in RAF1 Is the Priority Candidate Causative Mutation of the Inherited Chicken Wingless-2 Developmental Syndrome. Genes 2019, 10, 353. https://doi.org/10.3390/genes10050353
Youngworth I, Delany ME. A Premature Stop Codon in RAF1 Is the Priority Candidate Causative Mutation of the Inherited Chicken Wingless-2 Developmental Syndrome. Genes. 2019; 10(5):353. https://doi.org/10.3390/genes10050353
Chicago/Turabian StyleYoungworth, Ingrid, and Mary E. Delany. 2019. "A Premature Stop Codon in RAF1 Is the Priority Candidate Causative Mutation of the Inherited Chicken Wingless-2 Developmental Syndrome" Genes 10, no. 5: 353. https://doi.org/10.3390/genes10050353
APA StyleYoungworth, I., & Delany, M. E. (2019). A Premature Stop Codon in RAF1 Is the Priority Candidate Causative Mutation of the Inherited Chicken Wingless-2 Developmental Syndrome. Genes, 10(5), 353. https://doi.org/10.3390/genes10050353