Expression Profile of Porcine TRIM26 and Its Inhibitory Effect on Interferon-β Production and Antiviral Response

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Sample Collection
2.2. Cells, Viruses and Infections
2.3. Cloning and Sequence Analysis of porTRIM26
2.4. Generation of Polyclonal Antibody Against porTRIM26
2.5. Plasmid Transfection, Small Interfering RNA (siRNA), and Polyinosinic:polycytidylic Acid (poly (I:C)) Stimulation
2.6. Western Blottting Analysis
2.7. Enzyme-linked Immunosorbent Assay (ELISA)
2.8. Reverse transcription (RT)-quantitative PCR (qPCR) Analysis
2.9. Statistical Analysis
3. Results
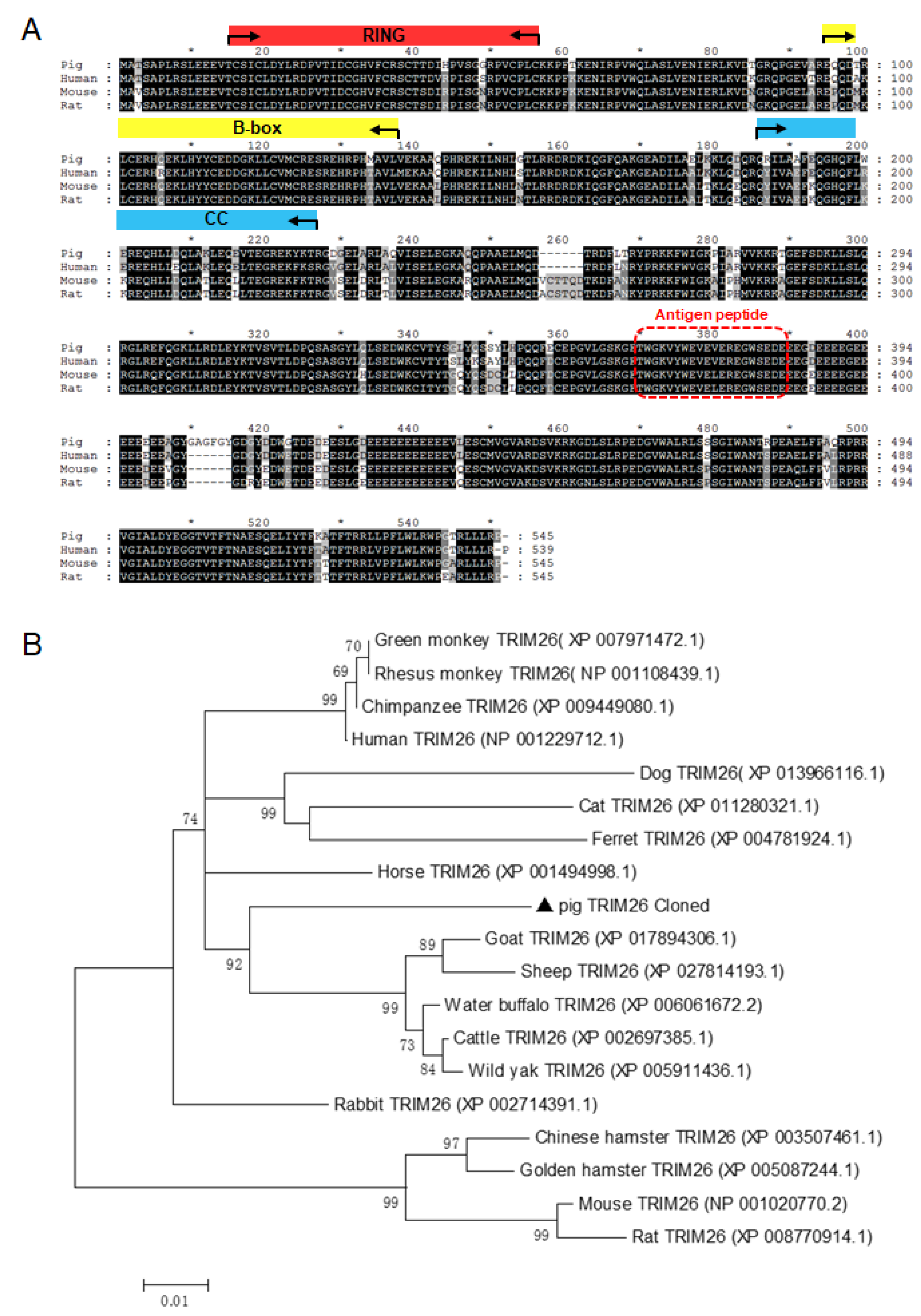
3.1. Sequence Analysis and Generation of Polyclonal Antibody Against porTRIM26
3.2. Expression Profiles of porTRIM26 in Different Cell Lines and Tissues
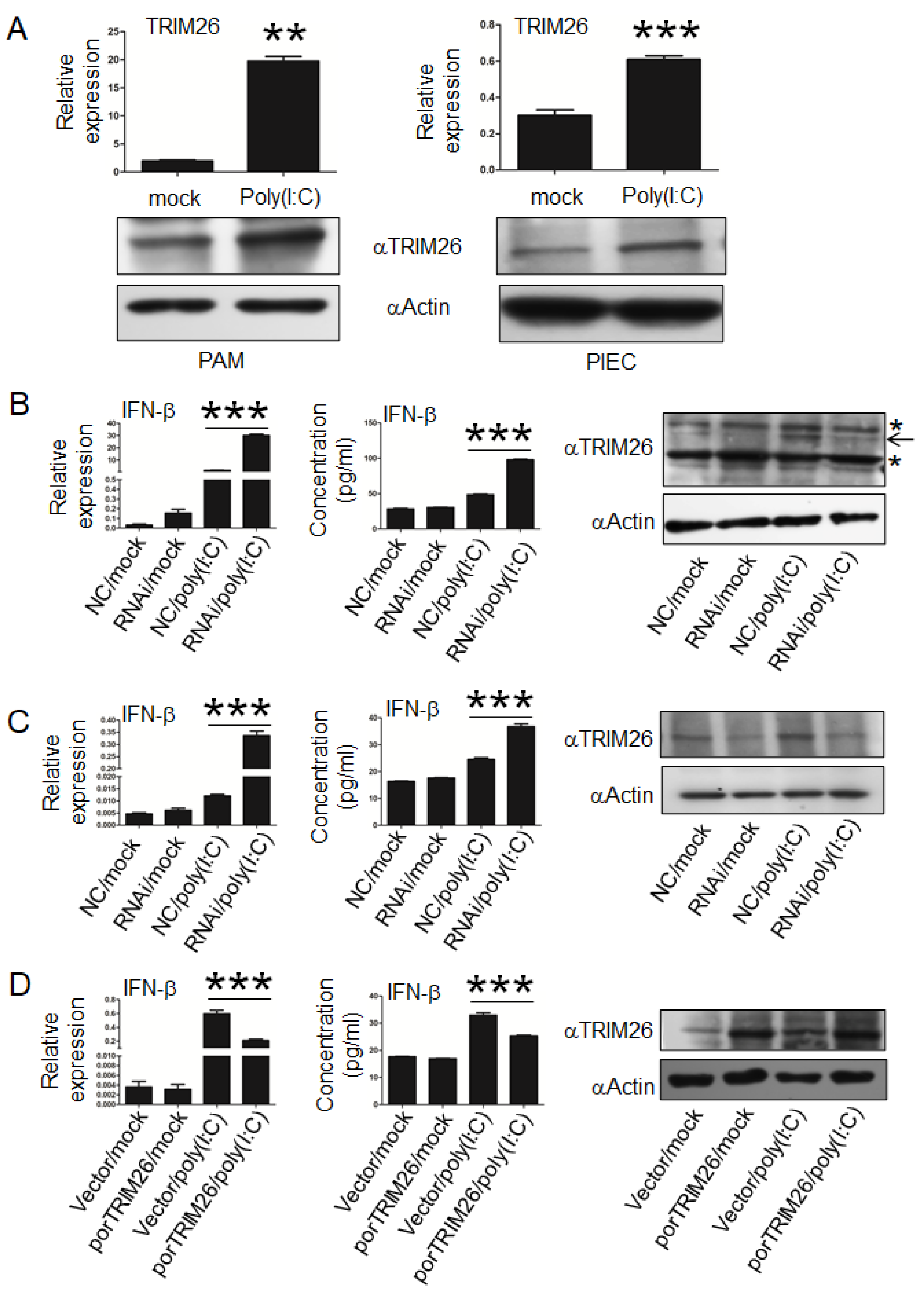
3.3. porTRIM26 Negatively Regulates Expression of IFN-β
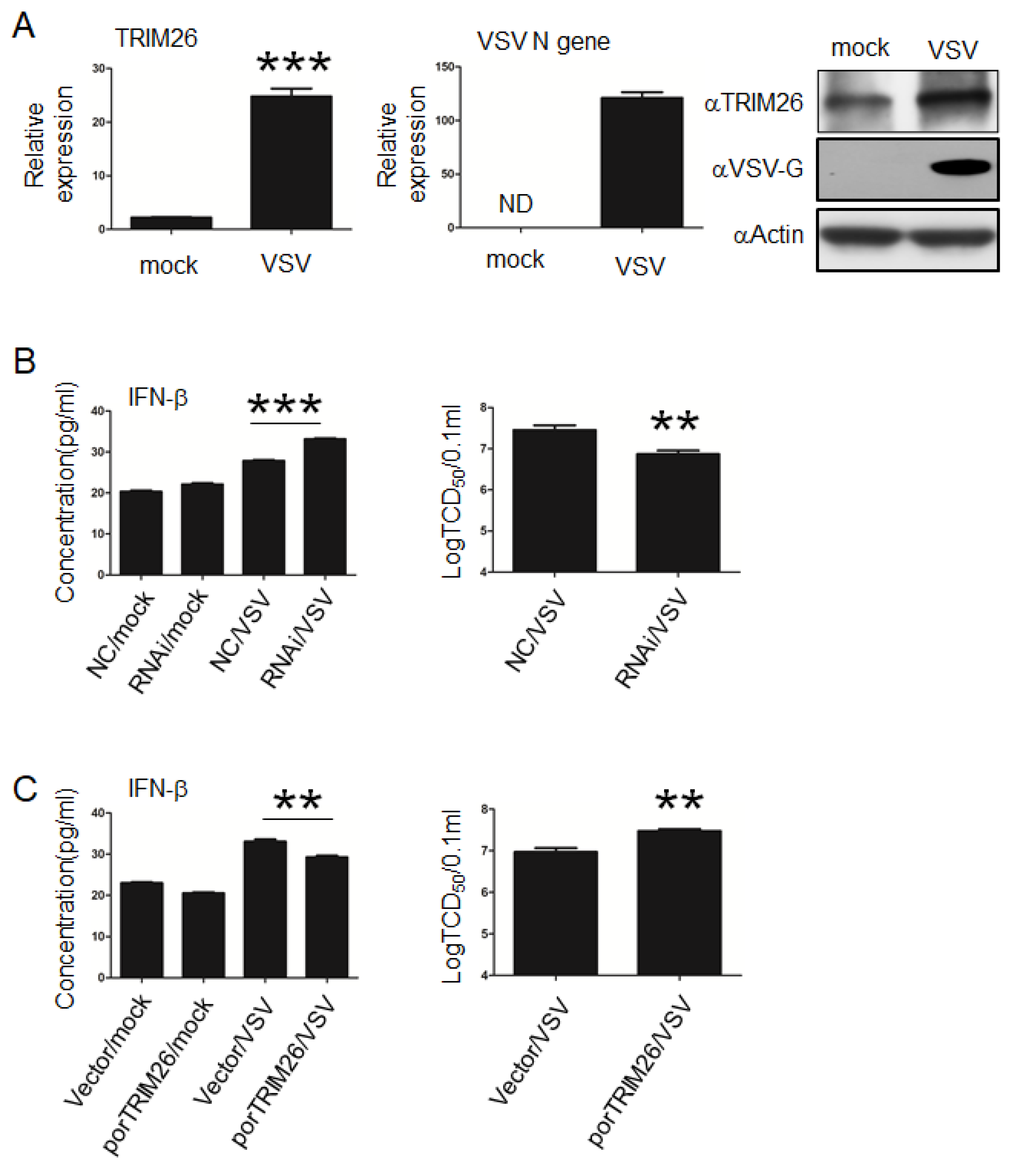
3.4. porTRIM26 Negatively Regulates Antiviral Response to VSV Infection
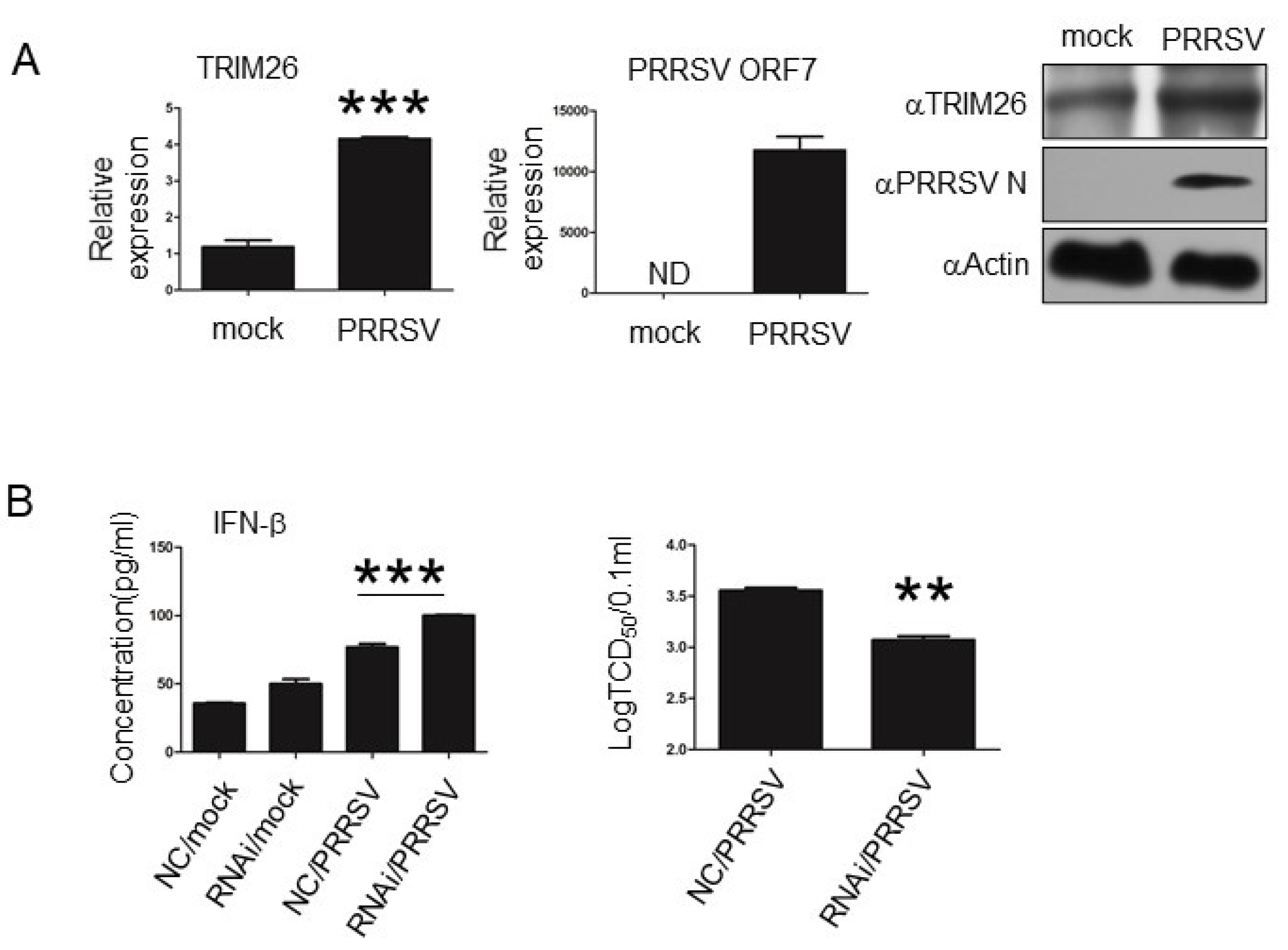
3.5. porTRIM26 Negatively Regulates Antiviral Response to PRRSV Infection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Rajsbaum, R.; Garcia-Sastre, A.; Versteeg, G.A. TRIMmunity: The roles of the TRIM E3-ubiquitin ligase family in innate antiviral immunity. J. Mol. Biol. 2014, 426, 1265–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renard, C.; Vaiman, M.; Chiannilkulchai, N.; Cattolico, L.; Robert, C.; Chardon, P. Sequence of the pig major histocompatibility region containing the classical class I genes. Immunogenetics 2001, 53, 490–500. [Google Scholar] [CrossRef]
- Kimsa, M.W.; Strzalka-Mrozik, B.; Kimsa, M.C.; Mazurek, U.; Kruszniewska-Rajs, C.; Gola, J.; Adamska, J.; Twardoch, M. Differential expression of tripartite motif-containing family in normal human dermal fibroblasts in response to porcine endogenous retrovirus infection. Folia Biol. 2014, 60, 144–151. [Google Scholar]
- van Tol, S.; Hage, A.; Giraldo, M.I.; Bharaj, P.; Rajsbaum, R. The TRIMendous Role of TRIMs in Virus-Host Interactions. Vaccines 2017, 5, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, H.; Sun, S.C. Ubiquitin signaling in immune responses. Cell Res. 2016, 26, 457–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nisole, S.; Stoye, J.P.; Saib, A. TRIM family proteins: Retroviral restriction and antiviral defence. Nat. Rev. Microbiol. 2005, 3, 799–808. [Google Scholar] [CrossRef] [PubMed]
- McNab, F.W.; Rajsbaum, R.; Stoye, J.P.; O’Garra, A. Tripartite-motif proteins and innate immune regulation. Curr. Opin. Immunol. 2011, 23, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Han, K.; Lou, D.I.; Sawyer, S.L. Identification of a genomic reservoir for new TRIM genes in primate genomes. PLoS Genet. 2011, 7, e1002388. [Google Scholar] [CrossRef] [Green Version]
- Short, K.M.; Cox, T.C. Subclassification of the RBCC/TRIM superfamily reveals a novel motif necessary for microtubule binding. J. Biol. Chem. 2006, 281, 8970–8980. [Google Scholar] [CrossRef] [Green Version]
- Chu, T.W.; Capossela, A.; Coleman, R.; Goei, V.L.; Nallur, G.; Gruen, J.R. Cloning of a new “finger” protein gene (ZNF173) within the class I region of the human MHC. Genomics 1995, 29, 229–239. [Google Scholar] [CrossRef]
- Genome-wide association study implicates HLA-C*01:02 as a risk factor at the major histocompatibility complex locus in schizophrenia. Biol. Psychiatry 2012, 72, 620–628. [CrossRef] [Green Version]
- Ando, A.; Shigenari, A.; Kulski, J.K.; Renard, C.; Chardon, P.; Shiina, T.; Inoko, H. Genomic sequence analysis of the 238-kb swine segment with a cluster of TRIM and olfactory receptor genes located, but with no class I genes, at the distal end of the SLA class I region. Immunogenetics 2005, 57, 864–873. [Google Scholar] [CrossRef] [PubMed]
- Tanaka-Matsuda, M.; Ando, A.; Rogel-Gaillard, C.; Chardon, P.; Uenishi, H. Difference in number of loci of swine leukocyte antigen classical class I genes among haplotypes. Genomics 2009, 93, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Ran, Y.; Zhang, J.; Liu, L.L.; Pan, Z.Y.; Nie, Y.; Zhang, H.Y.; Wang, Y.Y. Autoubiquitination of TRIM26 links TBK1 to NEMO in RLR-mediated innate antiviral immune response. J. Mol. Cell Biol. 2016, 8, 31–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Zhao, W.; Zhao, K.; Zhang, L.; Gao, C. TRIM26 negatively regulates interferon-beta production and antiviral response through polyubiquitination and degradation of nuclear IRF3. PLoS Pathog. 2015, 11, e1004726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, P.; Liu, K.; Wei, J.; Li, Y.; Li, B.; Shao, D.; Wu, Z.; Shi, Y.; Tong, G.; Qiu, Y.; et al. Nonstructural Protein 4 of Porcine Reproductive and Respiratory Syndrome Virus Modulates Cell Surface Swine Leukocyte Antigen Class I Expression by Downregulating beta2-Microglobulin Transcription. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Z.; Wei, J.; Deng, X.; Li, S.; Qiu, Y.; Shao, D.; Li, B.; Zhang, K.; Xue, F.; Wang, X.; et al. Nitazoxanide inhibits the replication of Japanese encephalitis virus in cultured cells and in a mouse model. Virol. J. 2014, 11, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lateef, S.S.; Gupta, S.; Jayathilaka, L.P.; Krishnanchettiar, S.; Huang, J.S.; Lee, B.S. An improved protocol for coupling synthetic peptides to carrier proteins for antibody production using DMF to solubilize peptides. J. Biomol. Tech. JBT 2007, 18, 173–176. [Google Scholar]
- Qiu, Y.; Shen, Y.; Li, X.; Ding, C.; Ma, Z. Molecular cloning and functional characterization of a novel isoform of chicken myeloid differentiation factor 88 (MyD88). Dev. Comp. Immunol. 2008, 32, 1522–1530. [Google Scholar] [CrossRef]
- Badaoui, B.; Rutigliano, T.; Anselmo, A.; Vanhee, M.; Nauwynck, H.; Giuffra, E.; Botti, S. RNA-sequence analysis of primary alveolar macrophages after in vitro infection with porcine reproductive and respiratory syndrome virus strains of differing virulence. PLoS ONE 2014, 9, e91918. [Google Scholar] [CrossRef]
- Fan, W.; Zhang, D.; Qian, P.; Qian, S.; Wu, M.; Chen, H.; Li, X. Swine TRIM21 restricts FMDV infection via an intracellular neutralization mechanism. Antivir. Res. 2016, 127, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Manocha, G.D.; Mishra, R.; Sharma, N.; Kumawat, K.L.; Basu, A.; Singh, S.K. Regulatory role of TRIM21 in the type-I interferon pathway in Japanese encephalitis virus-infected human microglial cells. J. Neuroinflamm. 2014, 11, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, K.; Shi, H.X.; Liu, X.Y.; Shan, Y.F.; Wei, B.; Chen, S.; Wang, C. TRIM21 is essential to sustain IFN regulatory factor 3 activation during antiviral response. J. Immunol. (Baltim. Md. 1950) 2009, 182, 3782–3792. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, H.; Sharma, M.; Zhang, Y.; Li, C.; Liu, K.; Wei, J.; Shao, D.; Li, B.; Ma, Z.; Cao, R.; et al. Expression Profile of Porcine TRIM26 and Its Inhibitory Effect on Interferon-β Production and Antiviral Response. Genes 2020, 11, 1226. https://doi.org/10.3390/genes11101226
Huang H, Sharma M, Zhang Y, Li C, Liu K, Wei J, Shao D, Li B, Ma Z, Cao R, et al. Expression Profile of Porcine TRIM26 and Its Inhibitory Effect on Interferon-β Production and Antiviral Response. Genes. 2020; 11(10):1226. https://doi.org/10.3390/genes11101226
Chicago/Turabian StyleHuang, Hui, Mona Sharma, Yanbing Zhang, Chenxi Li, Ke Liu, Jianchao Wei, Donghua Shao, Beibei Li, Zhiyong Ma, Ruibing Cao, and et al. 2020. "Expression Profile of Porcine TRIM26 and Its Inhibitory Effect on Interferon-β Production and Antiviral Response" Genes 11, no. 10: 1226. https://doi.org/10.3390/genes11101226
APA StyleHuang, H., Sharma, M., Zhang, Y., Li, C., Liu, K., Wei, J., Shao, D., Li, B., Ma, Z., Cao, R., & Qiu, Y. (2020). Expression Profile of Porcine TRIM26 and Its Inhibitory Effect on Interferon-β Production and Antiviral Response. Genes, 11(10), 1226. https://doi.org/10.3390/genes11101226