Latitudinal Cline in Chromosome Numbers of Ice Cod A. glacialis (Gadidae) from Northeast Greenland

 , and
, and
Abstract
:1. Introduction
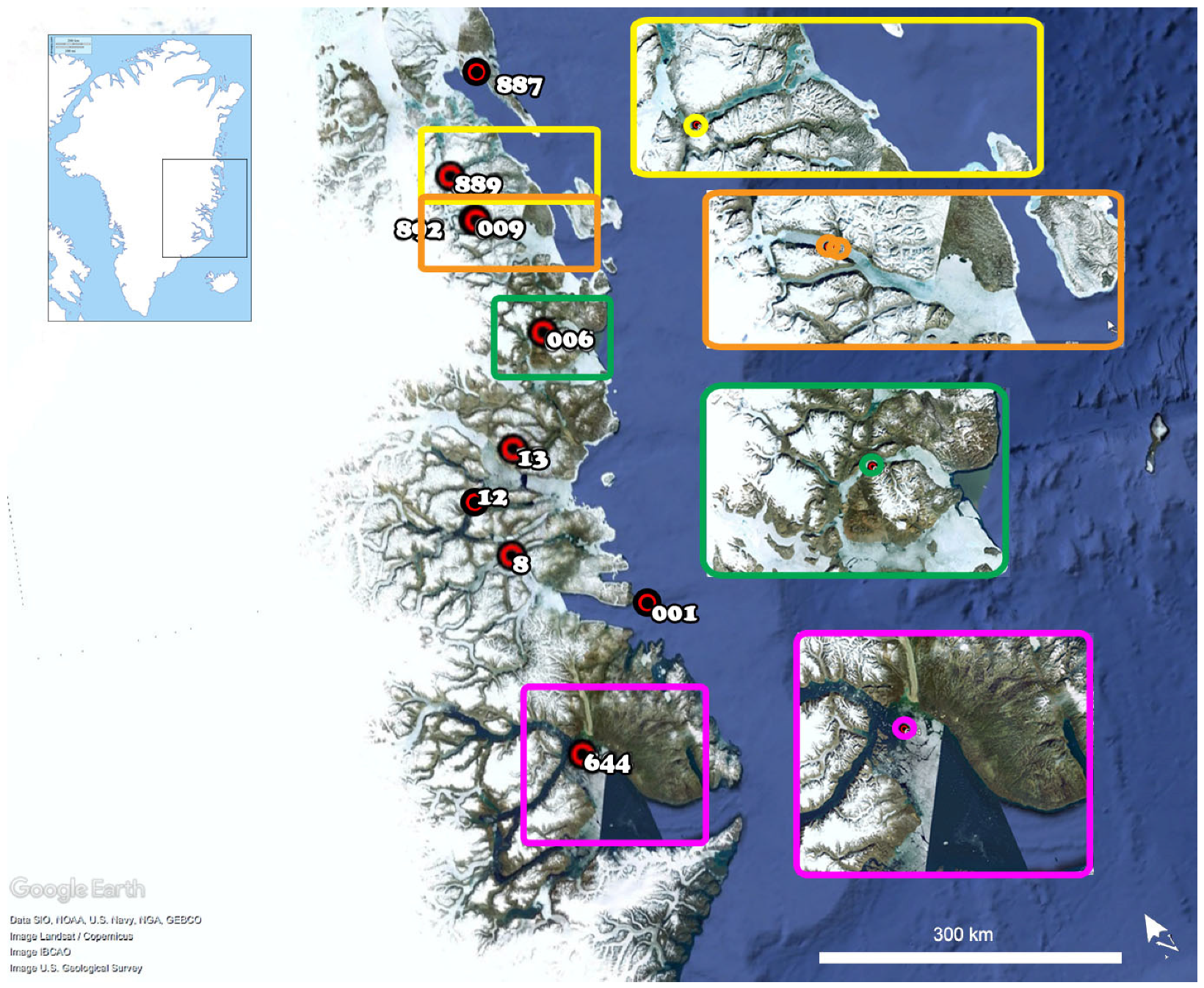
2. Materials and Methods
2.1. Sampling and Chromosome Preparation
2.2. Chromosome Analysis and Karyotyping
2.3. Statistical Analysis
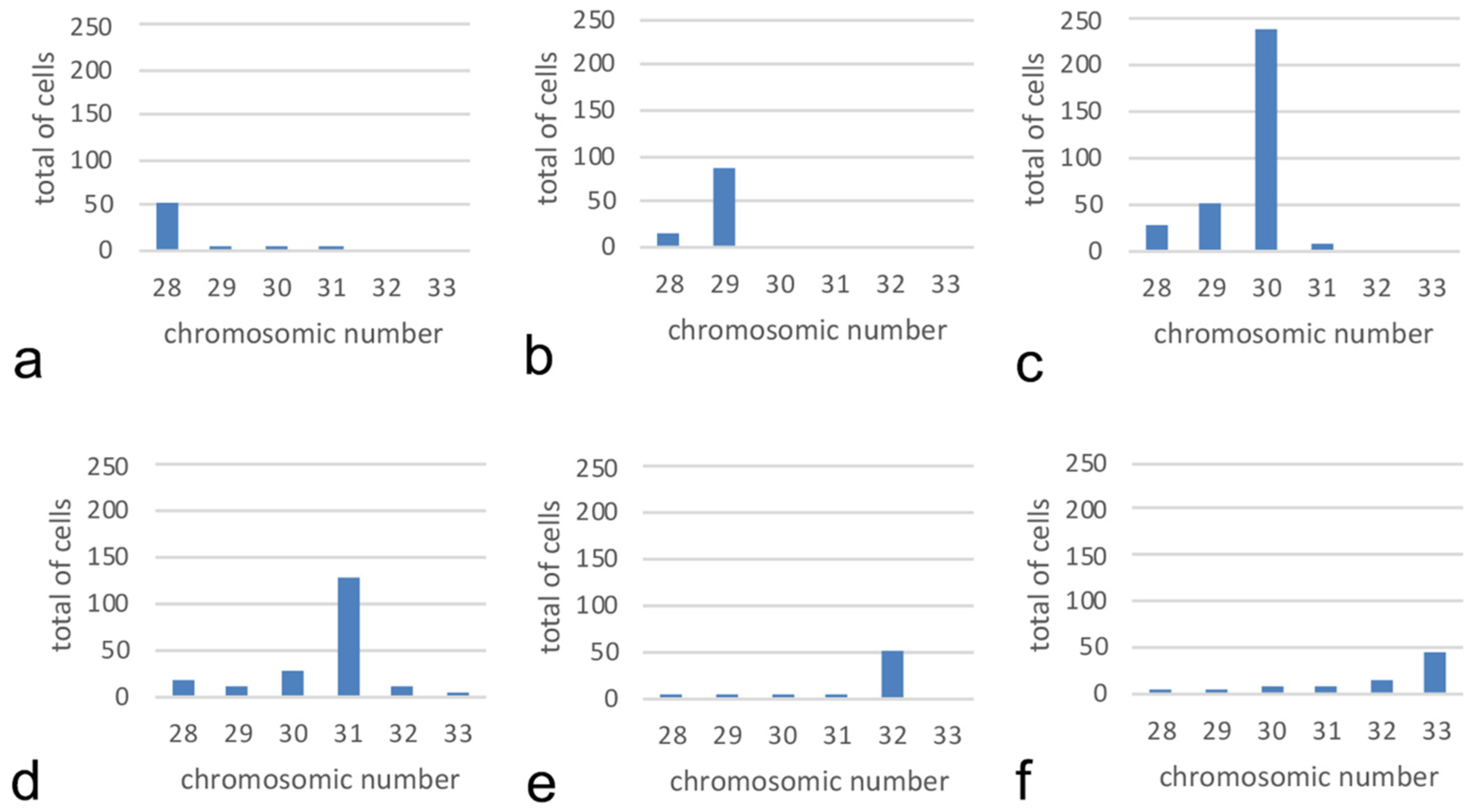
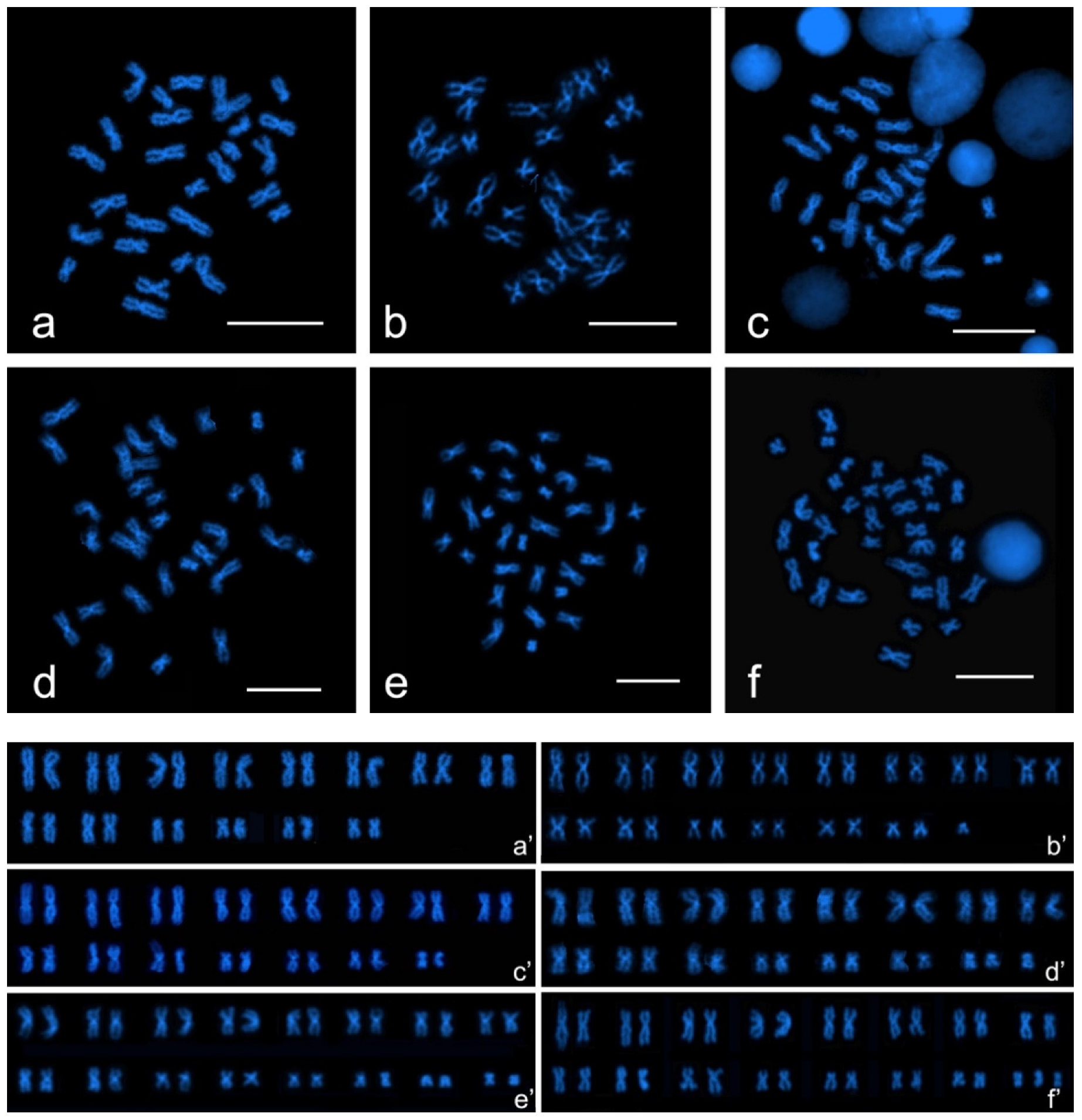
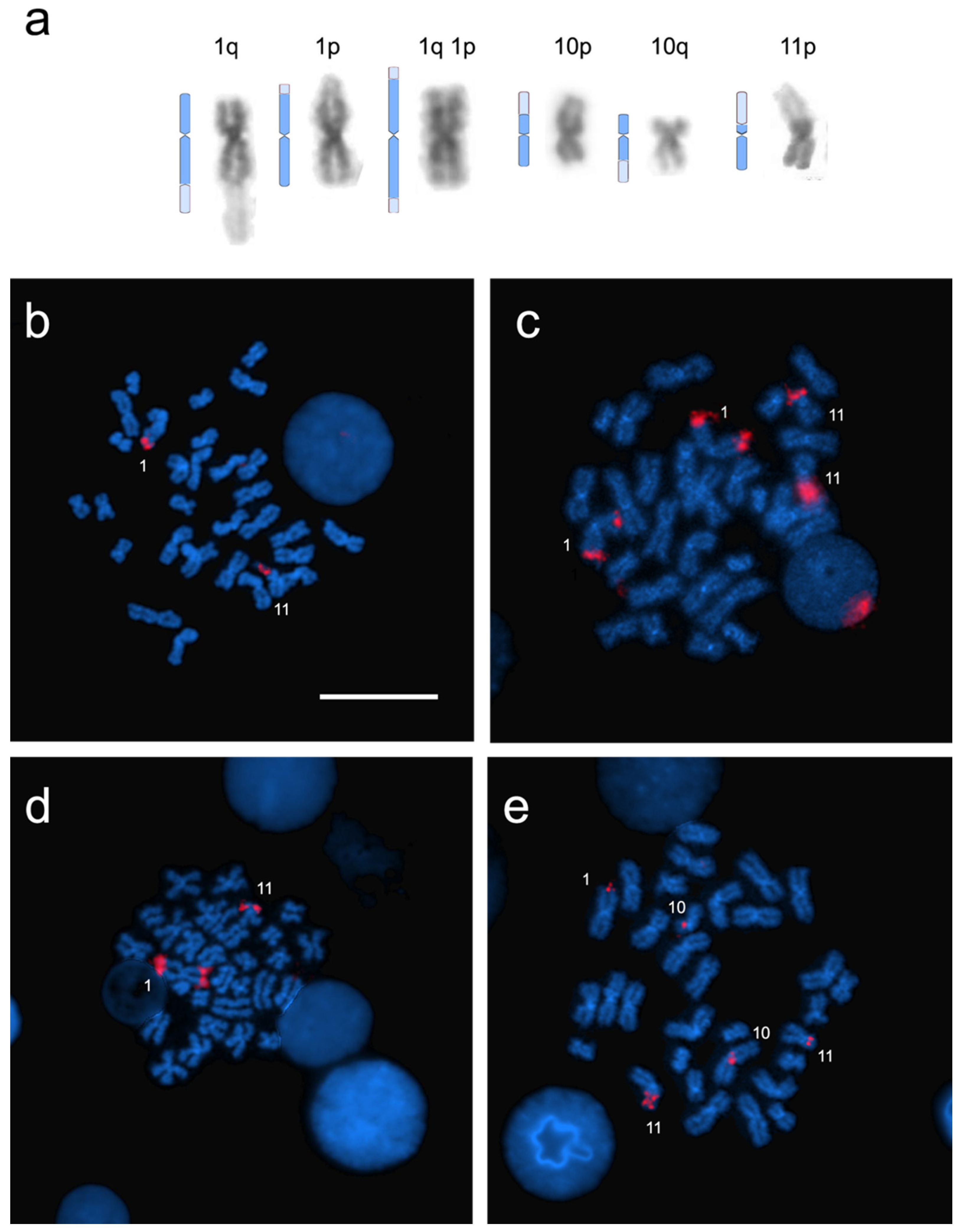
3. Results
3.1. Chromosome Number and Karyotype
3.2. Heterochromatin Distribution and Physical Mapping of Major Ribosomal Genes
3.3. Mitotic Instability, Chromosome Number, and Geographical Distribution
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Aschan, M.; Karamushko, O.V.; Byrkjedal, I.; Wienerroither, R.; Borkin, I.V.; Christiansen, J.S. Records of the gadoid fish Arctogadus glacialis (Peters, 1874) in the European Arctic. Polar Biol. 2009, 32, 963–970. [Google Scholar] [CrossRef]
- Nielsen, J.G.; Jensen, J.M. Revision of the Arctic cod genus, Arctogadus (Pisces: Gadidae). Med. Grønl. 1967, 184, 1–26. [Google Scholar]
- Jordan, A.D.; Møller, P.R.; Nielsen, J.G. Revision of the Arctic cod genus Arctogadus. J. Fish. Biol. 2003, 62, 1339–1352. [Google Scholar] [CrossRef]
- Mecklenburg, C.; Steinke, D. Ichthyofaunal Baselines in the Pacific Arctic Region and RUSALCA Study Area. Oceanography 2015, 28, 158–189. [Google Scholar] [CrossRef]
- Bouchard, C.; Mollard, S.; Suzuki, K.; Robert, D.; Fortier, L. Contrasting the early life histories of sympatric Arctic gadids Boreogadus saida and Arctogadus glacialis in the Canadian Beaufort Sea. Polar Biol. 2016, 39, 1005–1022. [Google Scholar] [CrossRef]
- Mueter, F.J.; Nahrgang, J.; Nelson, R.J.; Berge, J. The ecology of gadid fishes in the circumpolar Arctic with a special emphasis on the polar cod (Boreogadus saida). Polar Biol. 2016, 39, 961–967. [Google Scholar] [CrossRef]
- Süfke, L.; Piepenburg, D.; von Dorrien, C.F. Body size, sex ratio and diet composition of Arctogadus glacialis (Peters, 1874) (Pisces: Gadidae) in the Northeast Water Polynya (Greenland). Polar Biol. 1998, 20, 357–363. [Google Scholar] [CrossRef]
- Andriyashev, A.P. Fishes of the Northern seas of the U.S.S.R. In Keys to the Fauna of the U.S.S.R.; Pavlovskii, E.N., Ed.; Israel Program for Scienticic Translations: Jerusalem, Israel, 1964; p. 566. [Google Scholar]
- Christiansen, J.S.; Hop, H.; Nilssen, E.M.; Joensen, J. Trophic ecology of sympatric Arctic gadoids, Arctogadus glacialis (Peters, 1872) and Boreogadus saida (Lepechin, 1774), in NE Greenland. Polar Biol. 2012, 35, 1247–1257. [Google Scholar] [CrossRef]
- Mueter, F.; Bouchard, C.; Hop, H.; Laurel, B.; Norcross, B. Arctic gadids in a rapidly changing environment. Polar Biol. 2020, 43, 945–949. [Google Scholar] [CrossRef]
- Pálsson, S.; Källman, T.; Paulsen, J.; Árnason, E. An assessment of mitochondrial variation in Arctic gadoids. Polar Biol. 2009, 32, 471–479. [Google Scholar] [CrossRef]
- Wilson, R.E.; Sonsthagen, S.A.; Smé, N.; Gharrett, A.J.; Majewski, A.R.; Wedemeyer, K.; Talbot, S.L. Mitochondrial genome diversity and population mitogenomics of polar cod (Boreogadus saida) and Arctic dwelling gadoids. Polar Biol. 2020, 43, 979–994. [Google Scholar] [CrossRef]
- Breines, R.; Ursvik, A.; Nymark, M.; Johansen, S.D.; Coucheron, D.H. Complete mitochondrial genome sequences of the Arctic Ocean codfishes Arctogadus glacialis and Boreogadus saida reveal oriL and tRNA gene duplications. Polar Biol. 2008, 31, 1245–1252. [Google Scholar] [CrossRef] [Green Version]
- Madsen, M.L.; Fevolden, S.E.; Christiansen, J.S. A simple molecular approach to distinguish between two Arctic gadoid fishes Arctogadus glacialis (Peters, 1874) and Boreogadus saida (Lepechin, 1774). Polar Biol. 2009, 32, 937–939. [Google Scholar] [CrossRef]
- Nelson, R.J.; Bouchard, C.; Madsen, M.; Praebel, K.; Rondeau, E.; Schalburg, K.; Koop, B.F. Microsatellite loci for genetic analysis of the arctic gadids Boreogadus saida and Arctogadus glacialis. Conserv. Genet. Resour. 2012, 5, 445–448. [Google Scholar] [CrossRef]
- Nelson, R.J.; Bouchard, C.; Fortier, A.L.; Majewski, A.R.; Reist, J.D.; Praebel, K.; Divoky, G.J. Circumpolar genetic population structure of polar cod, Boreogadus saida. Polar Biol. 2020, 43, 951–961. [Google Scholar] [CrossRef]
- Silva, D.M.Z.d.A.; Pansonato-Alves, J.C.; Utsunomia, R.; Araya-Jaime, C.; Ruiz-Ruano, F.J.; Daniel, S.N.; Hashimoto, D.T.; Oliveira, C.; Camacho, J.P.M.; Porto-Foresti, F.; et al. Delimiting the Origin of a B Chromosome by FISH Mapping, Chromosome Painting and DNA Sequence Analysis in Astyanax paranae (Teleostei, Characiformes). PLoS ONE 2014, 9, e94896. [Google Scholar] [CrossRef] [Green Version]
- Artoni, R.F.; Castro, J.P.; Jacobina, U.P.; Lima-Filho, P.A.; Félix da Costa, G.W.W.; Molina, W.F. Inferring diversity and evolution in fish by means of integrative molecular cytogenetics. Sci. World J. 2015, 365787. [Google Scholar] [CrossRef] [Green Version]
- Symonová, R.; Howell, W.M. Vertebrate genome evolution in the light of fish cytogenomics and rDNAomics. Genes 2018, 9, 96. [Google Scholar] [CrossRef] [Green Version]
- Ghigliotti, L.; Mazzei, F.; Fevolden, S.E.; Christiansen, J.S.; Pisano, E. First cytogenetic analysis of the Arctic fishes Boreogadus saida (polar cod) and Arctogadus glacialis (ice cod), family Gadidae. In Polarnet Technical Report; Luporini, P., Morbidoni, M., Eds.; Polarnet Coordinating Unit: Roma, Italy, 2005; pp. 69–73. [Google Scholar]
- Christiansen, J.S. The TUNU-Programme: Euro-Arctic marine fishes—diversity and adaptation. In Adaptation and Evolution in Marine Environments; di Prisco, G., Verde, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; Volume 1, pp. 35–50. [Google Scholar] [CrossRef]
- Rey, O.; d’Hont, A.; Coutanceau, J.; Pisano, E.; Chilmonczyk, S.; Ozouf-Costaz, C. Cephalic kidney and spleen cell culture in Antarctic teleosts. In Fish Cytogenetic Techniques: Ray-Fin Fishes Chondrichthyans; Ozouf-Costaz, C., Pisano, E., Foresti, F., Almeida Toledo, L.F., Eds.; Taylor Francis Group: Abingdon, UK; CRC Press: London, UK, 2015; pp. 74–81. [Google Scholar]
- Ghigliotti, L.; Cheng, C.C.H.; Ozouf-Costaz, C.; Vacchi, M.; Pisano, E. Cytogenetic diversity of notothenioid fish from the Ross sea: Historical overview and updates. Hydrobiologia 2015, 761, 373–396. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Sakamoto, Y.; Zacaro, A.A. LEVAN, An ImageJ Plugin for Morphological Cytogenetic Analysis of Mitotic and Meiotic Chromosomes. Initial Version. An open source Java plugin distributed over the Internet. Available online: http://rsbweb.nih.gov/ij/ (accessed on 17 November 2020).
- Levan, A. Nomenclature for centromeric position on chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Ghigliotti, L.; Mazzei, F.; Ozouf-Costaz, C.; Bonillo, C.; Williams, R.; Cheng, C.-H.C.; Pisano, E. The two giant sister species of the Southern Ocean, Dissostichus eleginoides and Dissostichus mawsoni, differ in karyotype and chromosomal pattern of ribosomal RNA genes. Polar Biol. 2007, 30, 625–634. [Google Scholar] [CrossRef]
- Pardo, M.C.; López-León, M.D.; Viseras, E.; Cabrero, J.; Camacho, J.P.M. Mitotic instability of B chromosomes during embryo development in Locusta migratoria. Heredity 1995, 74, 164–169. [Google Scholar] [CrossRef] [Green Version]
- Kahl, G. The Dictionary of Genomics, Transcriptomics and Proteomics; John Wiley & Sons: Weinheim, Germany, 2015. [Google Scholar]
- Molina, W.F. Chromosomal changes and stasis in marine fish groups. In Fish Cytogenetics; Universidade Federal do Rio Grande do Norte: Natal, Brazil, 2007; pp. 69–110. [Google Scholar]
- Paim, F.G.; da Hora Almeida, L.A.; de Mello Affonso, P.R.A.; Sobrinho-Scudeler, P.E.; Oliveira, C.; Diniz, D. Chromosomal stasis in distinct families of marine Percomorpharia from South Atlantic. Comp. Cytogenet. 2017, 11, 299. [Google Scholar] [CrossRef]
- Motta Neto, C.C.; Cioffi, M.D.B.; Costa, G.W.W.F.; Amorim, K.D.J.; Bertollo, L.A.C.; Artoni, R.F.; Molina, W.F. Overview on Karyotype Stasis in Atlantic Grunts (Eupercaria, Haemulidae) and the Evolutionary Extensions for Other Marine Fish Groups. Front. Mar. Sci. 2019, 6, 628. [Google Scholar] [CrossRef] [Green Version]
- Völker, M.; Sonnenberg, R.; Ráb, P.; Kullmann, H. Karyotype differentiation in Chromaphyosemion killifishes (Cyprinodontiformes, Nothobranchiidae). III: Extensive karyotypic variability associated with low mitochondrial haplotype differentiation in C. bivittatum. Cytogenet. Genome. Res. 2007, 116, 116–126. [Google Scholar] [CrossRef]
- Milhomem, S.S.; Pieczarka, J.C.; Crampton, W.G.; Silva, D.S.; De Souza, A.C.; Carvalho, J.R.; Nagamachi, C.Y. Chromosomal evidence for a putative cryptic species in the Gymnotus carapo species-complex (Gymnotiformes, Gymnotidae). BMC Genet. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Ibagón, N.; Maldonado-Ocampo, J.A.; Cioffi, M.D.B.; Dergam, J.A. Chromosomal diversity of Hoplias malabaricus (Characiformes, Erythrinidae) along the Magdalena River (Colombia—northern South America) and its significance for the Neotropical Region. Zebrafish 2020, 17, 211–219. [Google Scholar] [CrossRef]
- Vitturi, R.; Catalano, E.; Conte, M.L.; Alessi, A.M.; Amico, F.P.; Colombera, D. Intra-populational and intra-individual mosaicisms of Uranoscopus scaber L. (Perciformes, Uranoscopidae). Heredity 1991, 67, 325–330. [Google Scholar] [CrossRef] [Green Version]
- Lima-Filho, P.A.; de Bello Cioffi, M.; Bertollo, L.A.C.; Molina, W.F. Chromosomal and morphological divergences in Atlantic populations of the frillfin goby Bathygobius soporator (Gobiidae, Perciformes). J. Exp. Mar. Biol. Ecol. 2012, 434, 63–70. [Google Scholar] [CrossRef]
- Nirchio, M.; Gaviria, J.I.; Siccha-Ramirez, Z.R.; Oliveira, C.; Foresti, F.; Milana, V.; Rossi, A.R. Chromosomal polymorphism and molecular variability in the pearly razorfish Xyrichtys novacula (Labriformes, Labridae): Taxonomic and biogeographic implications. Genetica 2019, 147, 47–56. [Google Scholar] [CrossRef]
- Fan, Z.; Fox, D.P. Robertsonian polymorphism in plaice, Pleuronectes platessa L., and cod, Gadus morhua L., (Pisces Pleuronectiformes and Gadiformes). J. Fish Biol. 1991, 38, 635–640. [Google Scholar] [CrossRef]
- D’Ambrosio, U.; Alonso-Lifante, M.P.; Barros, K.; Kovařík, A.; Mas de Xaxars, G.; Garcia, S. B-chrom: A database on B-chromosomes of plants, animals and fungi. New Phytol. 2017, 216, 635–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venere, P.C.; Miyazawa, C.S.; Galetti, P.M., Jr. New cases of supernumerary chromosomes in Characiform fishes. Genet. Mol. Biol. 1999, 22, 345–349. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, R.A.; Martins-Santos, I.C.; Dias, A.S. B chromosomes: An update about their occurrence in freshwater Neotropical fishes (Teleostei). J. Fish Biol. 2008, 72, 1907–1932. [Google Scholar] [CrossRef]
- Lui, R.L.; Blanco, D.R.; Margarido, V.P.; Moreira Filho, O. First description of B chromosomes in the family Auchenipteridae, Parauchenipterus galeatus (Siluriformes) of the São Francisco River basin (MG, Brazil). Micron 2009, 40, 552–559. [Google Scholar] [CrossRef]
- Molina, W.F.; Martinez, P.A.; Bertollo, L.A.; Bidau, C.J. Preferential accumulation of sex and Bs chromosomes in biarmed karyotypes by meiotic drive and rates of chromosomal changes in fishes. An. Acad. Bras. Ciênc. 2014, 86, 1801–1812. [Google Scholar] [CrossRef] [Green Version]
- Arai, R. Fish Karyotypes: A Check List; Springer: Tokyo, Japan, 2011; pp. 131–132. [Google Scholar]
- Camacho, J.P.M. B chromosomes. In The Evolution of the Genome; Gregory, T.R., Ed.; Elsevier: San Diego, CA, USA, 2005; pp. 223–286. [Google Scholar] [CrossRef]
- Utsunomia, R.; Silva, D.M.Z.A.; Ruiz-Ruano, F.J.; Araya-Jaime, C.; Pansonato-Alves, J.C.; Scacchetti, P.C.; Foresti, F. Uncovering the Ancestry of B Chromosomes in Moenkhausia sanctaefilomenae (Teleostei, Characidae). PLoS ONE 2016, 11, e0150573. [Google Scholar] [CrossRef] [Green Version]
- Jehangir, M.; Ahmad, S.F.; Cardoso, A.L.; Ramos, E.; Valente, G.T.; Martins, C. De novo genome assembly of the cichlid fish Astatotilapia latifasciata reveals a higher level of genomic polymorphism and genes related to B chromosomes. Chromosoma 2019, 128, 81–96. [Google Scholar] [CrossRef]
- Valente, G.T.; Nakajima, R.T.; Fantinatti, B.E.; Marques, D.F.; Almeida, R.O.; Simões, R.P.; Martins, C. B chromosomes: From cytogenetics to systems biology. Chromosoma 2017, 126, 73–81. [Google Scholar] [CrossRef]
- Ahmad, S.F.; Jehangir, M.; Cardoso, A.L.; Wolf, I.R.; Margarido, V.P.; Cabral-de-Mello, D.C.; Martins, C. B chromosomes of multiple species have intense evolutionary dynamics and accumulated genes related to important biological processes. BMC Genom. 2020, 21, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Pires, L.B.; Sampaio, T.R.; Dias, A.L. Mitotic and meiotic behavior of B chromosomes in Crenicichla lepidota: New report in the family cichlidae. J. Hered. 2015, 106, 289–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macgregor, H.C.; Vlad, M.; Barnett, L. An investigation of some problems concerning nucleolus organizers in Salamanders. Chromosoma 1977, 59, 283–299. [Google Scholar] [CrossRef]
- Silva-Neto, L.C.; Bernardino, A.C.S.; Loreto, V.; Moura, R.C. Physical mapping of 18S and 5S rDNA loci and histone H3 gene in grasshopper species of the subfamily Gomphocerinae (Acrididae). Genet. Mol. Res. 2015, 14, 15008–15015. [Google Scholar] [CrossRef] [PubMed]
- Drouin, G.; De Sa, M.M. The concerted evolution of 5S ribosomal genes linked to the repeat units of other multigene families. Mol. Biol. Evol. 1995, 12, 481–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gornung, E. Twenty years of physical mapping of major ribosomal RNA genes across the teleosts: A review of research. Cytogen. Genome Res. 2013, 141, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Rebordinos, L.; Cross, I.; Merlo, A. High evolutionary dynamism in 5S rDNA of fish: State of the art. Cytogen. Genome Res. 2013, 141, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Symonová, R.; Majtánová, Z.; Sember, A.; Staaks, G.B.; Bohlen, J.; Freyhof, J.; Ráb, P. Genome differentiation in a species pair of coregonine fishes: An extremely rapid speciation driven by stress-activated retrotransposons mediating extensive ribosomal DNA multiplications. BMC Evol. Biol. 2013, 13, 42. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Expedition | Year | Station ID | Latitude N | Longitude W | Depth | Males | Females |
|---|---|---|---|---|---|---|---|
| TUNU I | 2003 | 887 | 76°43′ | 19°17′ | 231 | 1 | 0 |
| TUNU I | 2003 | 889 | 75°58′ | 21°41′ | 234 | 5 | 6 |
| TUNU I | 2003 | 892 | 75°33′ | 21°39′ | 571 | 2 | 3 |
| TUNU II | 2005 | 644 | 71°09′ | 24°57′ | 483 | 11 | 8 |
| TUNU IV | 2010 | 8 | 72°53′ | 24°40′ | 327 | 0 | 1 |
| TUNU IV | 2010 | 12 | 73°25′ | 25°17′ | 440 | 2 | 1 |
| TUNU IV | 2010 | 13 | 73°42′ | 23°28′ | 232 | 0 | 1 |
| TUNU V | 2013 | 001 | 72°00′ | 21°01′ | 466 | 0 | 1 |
| TUNU V | 2013 | 006 | 74°27′ | 21°10′ | 316 | 5 | 4 |
| TUNU V | 2013 | 009 | 75°33′ | 21°38′ | 620 | 2 | 1 |
| Specimen total | 28 | 26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghigliotti, L.; Christiansen, J.S.; Carlig, E.; Di Blasi, D.; Pisano, E. Latitudinal Cline in Chromosome Numbers of Ice Cod A. glacialis (Gadidae) from Northeast Greenland. Genes 2020, 11, 1515. https://doi.org/10.3390/genes11121515
Ghigliotti L, Christiansen JS, Carlig E, Di Blasi D, Pisano E. Latitudinal Cline in Chromosome Numbers of Ice Cod A. glacialis (Gadidae) from Northeast Greenland. Genes. 2020; 11(12):1515. https://doi.org/10.3390/genes11121515
Chicago/Turabian StyleGhigliotti, Laura, Jørgen S. Christiansen, Erica Carlig, Davide Di Blasi, and Eva Pisano. 2020. "Latitudinal Cline in Chromosome Numbers of Ice Cod A. glacialis (Gadidae) from Northeast Greenland" Genes 11, no. 12: 1515. https://doi.org/10.3390/genes11121515
APA StyleGhigliotti, L., Christiansen, J. S., Carlig, E., Di Blasi, D., & Pisano, E. (2020). Latitudinal Cline in Chromosome Numbers of Ice Cod A. glacialis (Gadidae) from Northeast Greenland. Genes, 11(12), 1515. https://doi.org/10.3390/genes11121515