Diversity of Antimicrobial Peptides in Three Partially Sympatric Frog Species in Northeast Asia and Implications for Evolution

Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Skin Tissues
2.2. Cloning of cDNA of Antimicrobial Peptide (AMP) Genes
2.3. Prediction of Physicochemical Properties and Secondary Structure of Antimicrobial Peptides (AMPs)
3. Results
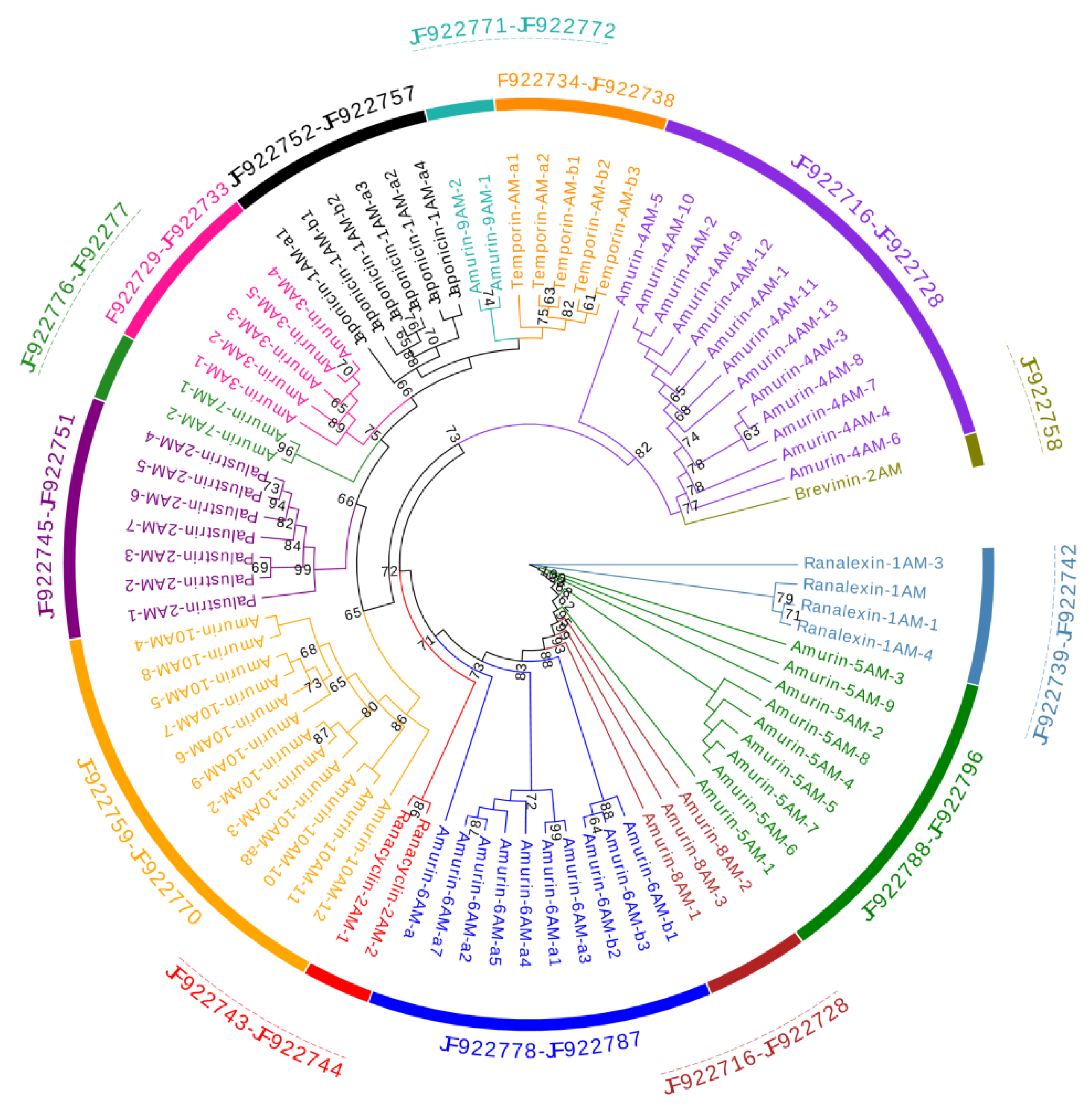
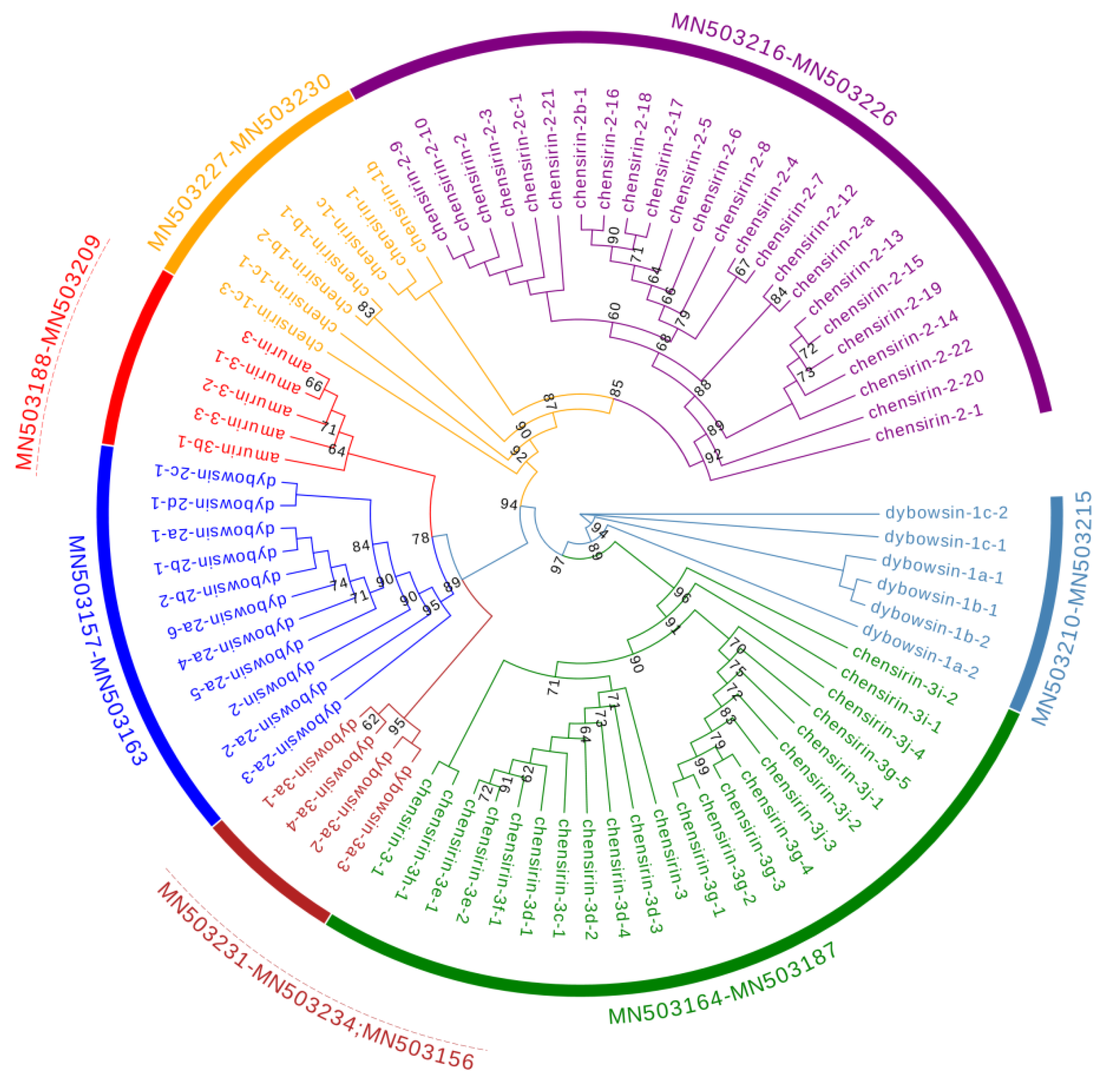
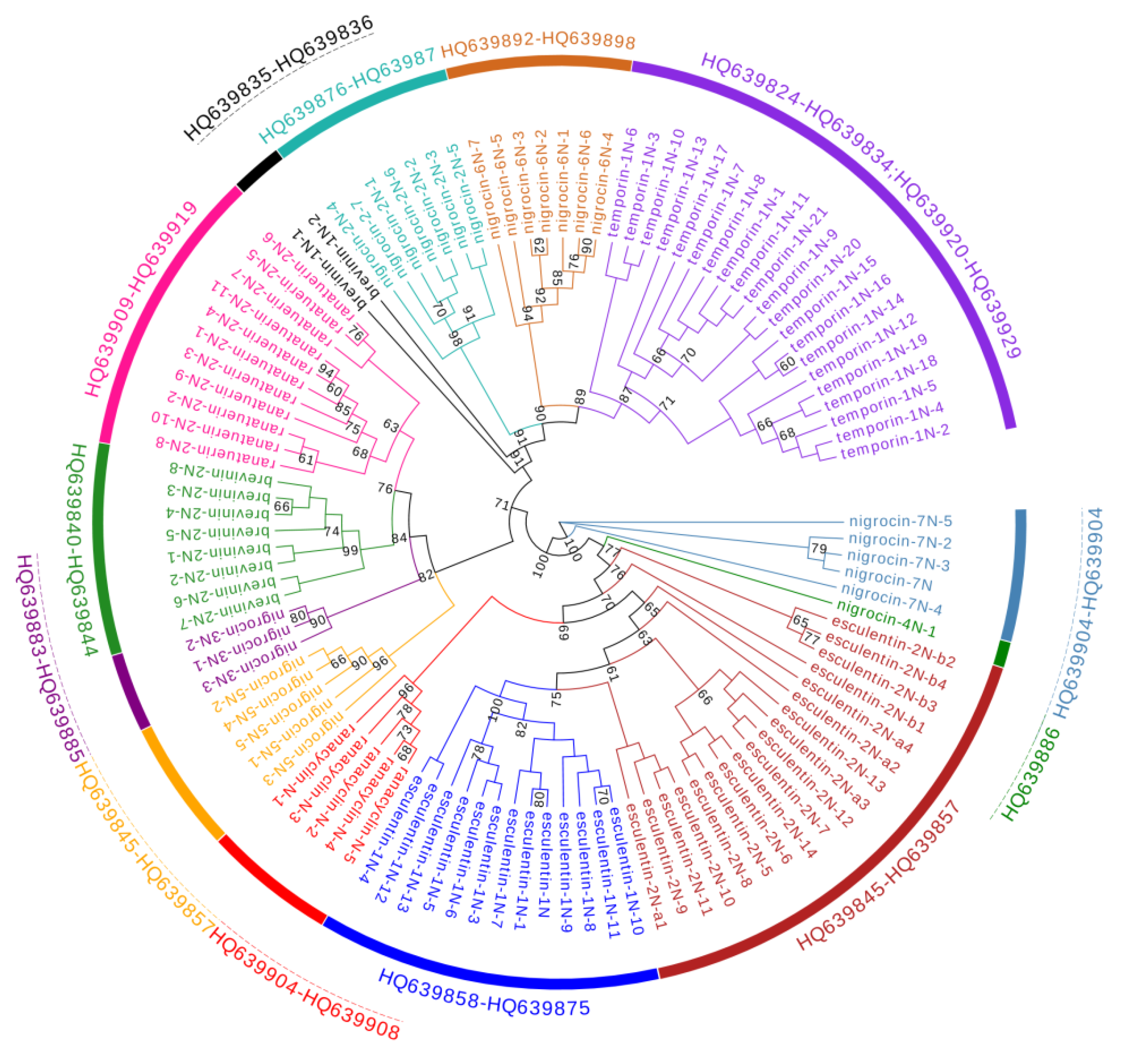
3.1. Cloning and Sequence Analysis of Antimicrobial Peptide (AMP) cDNAs
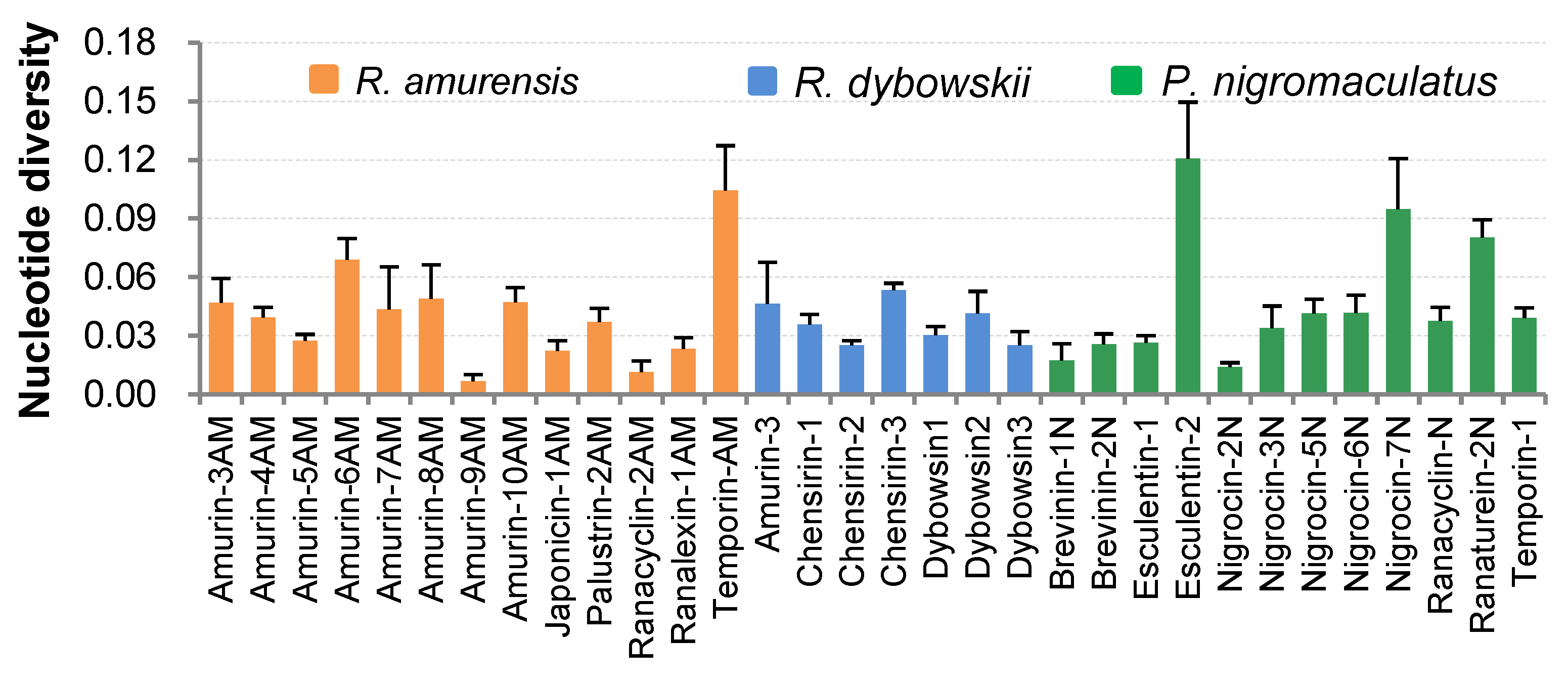
3.2. Nucleotide Diversity of Antimicrobial Peptide (AMP) Families
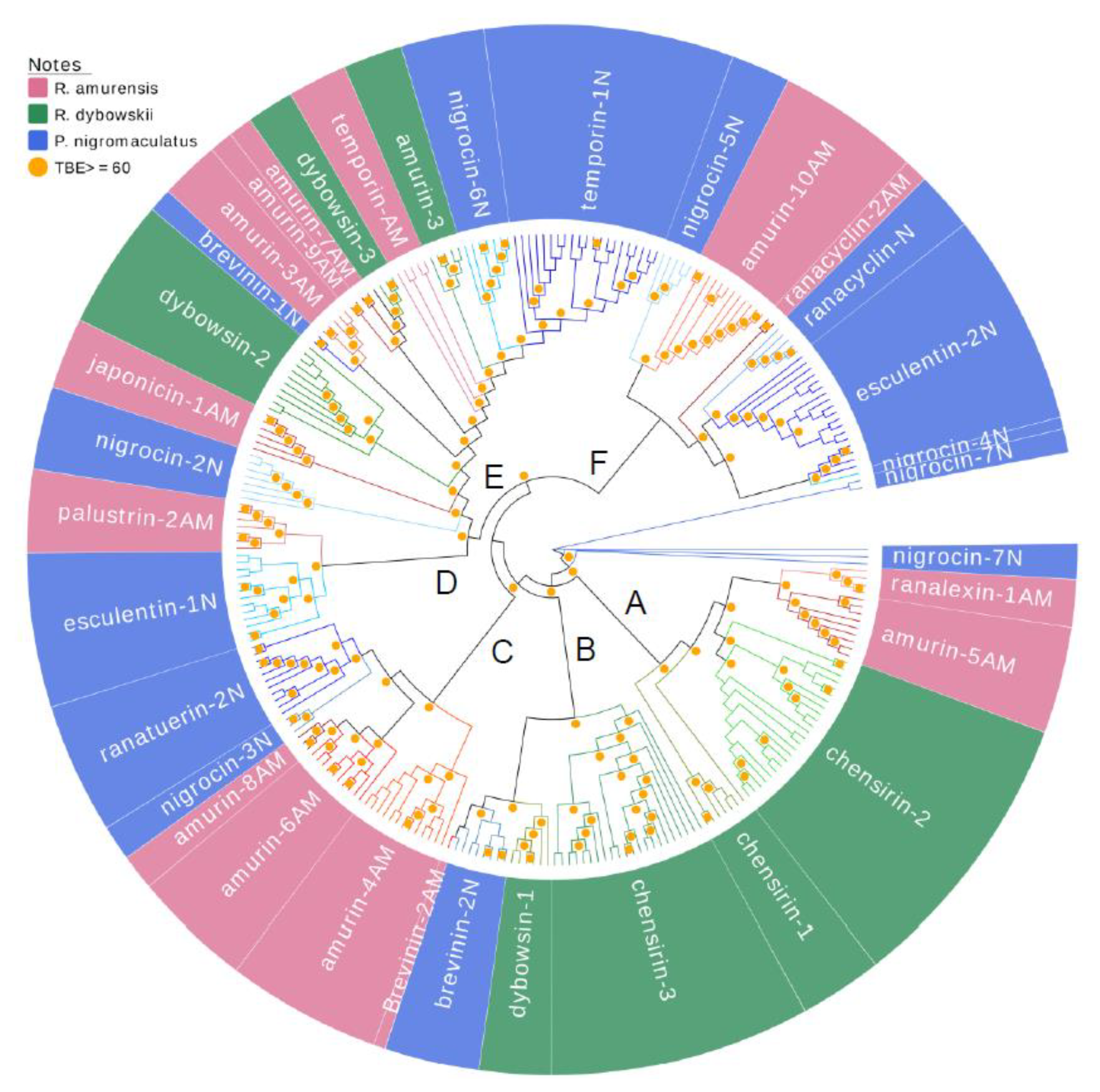
3.3. Divergence Pattern of Antimicrobial Peptides (AMPs) in Three Species
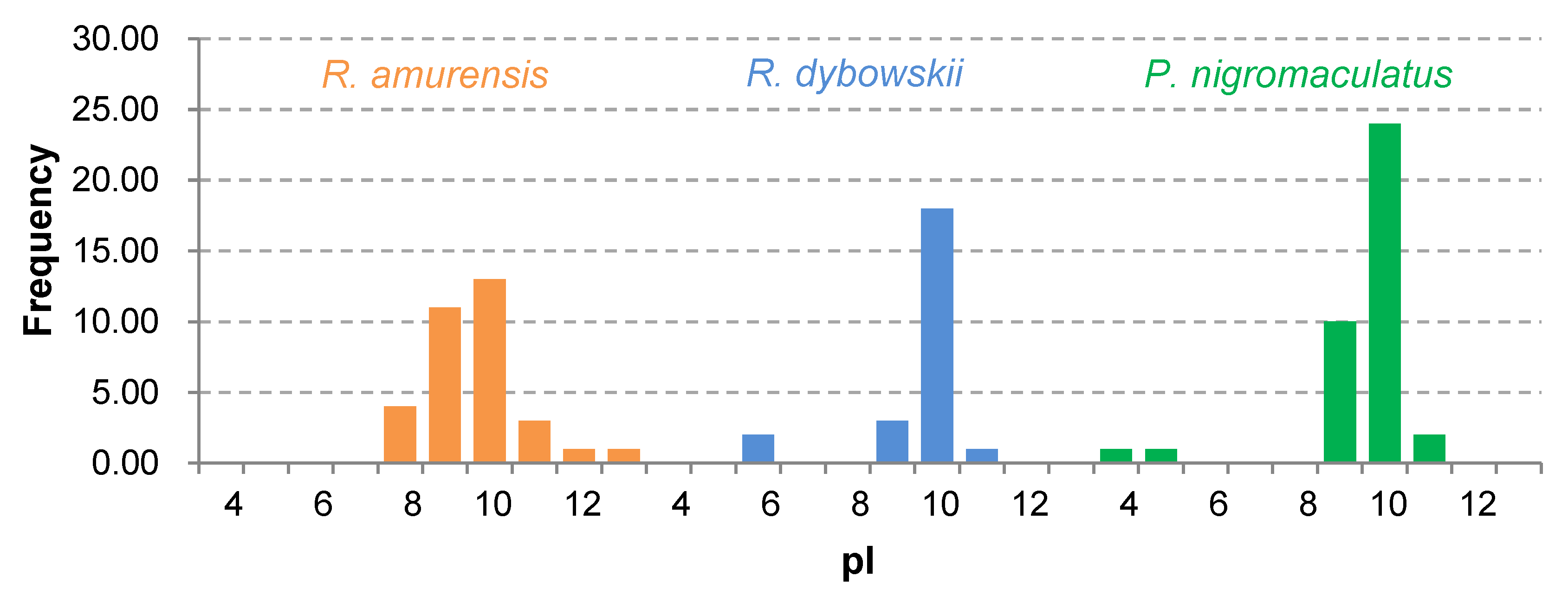
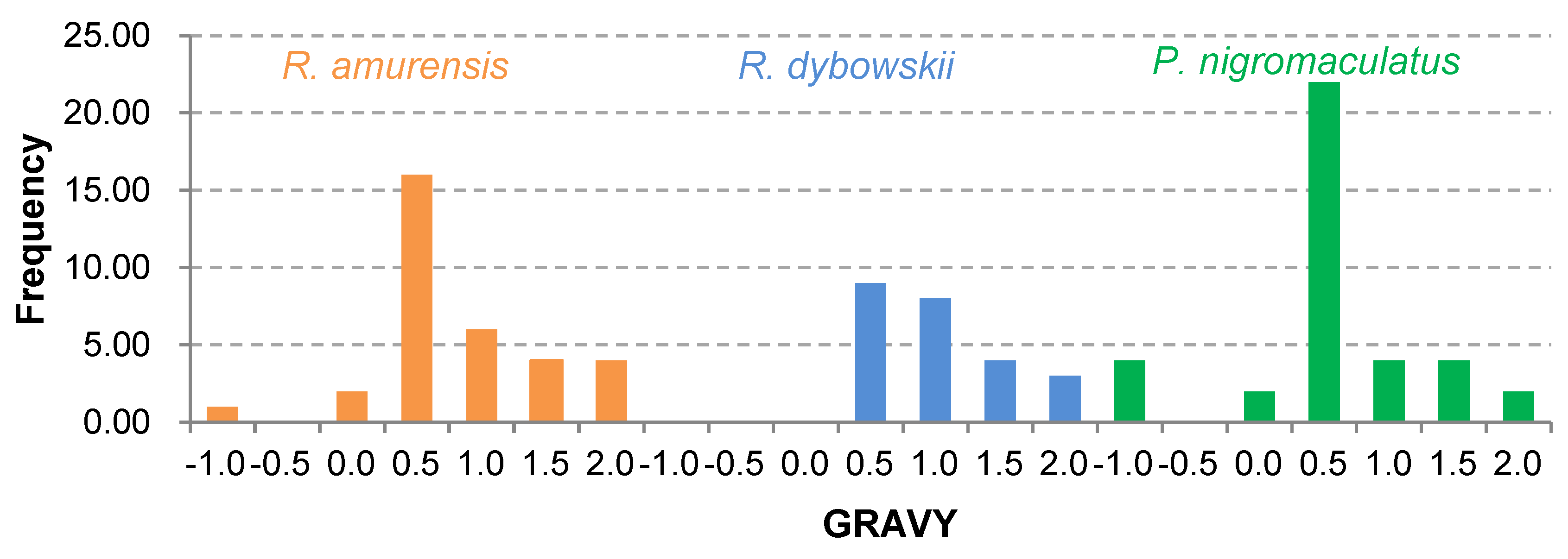
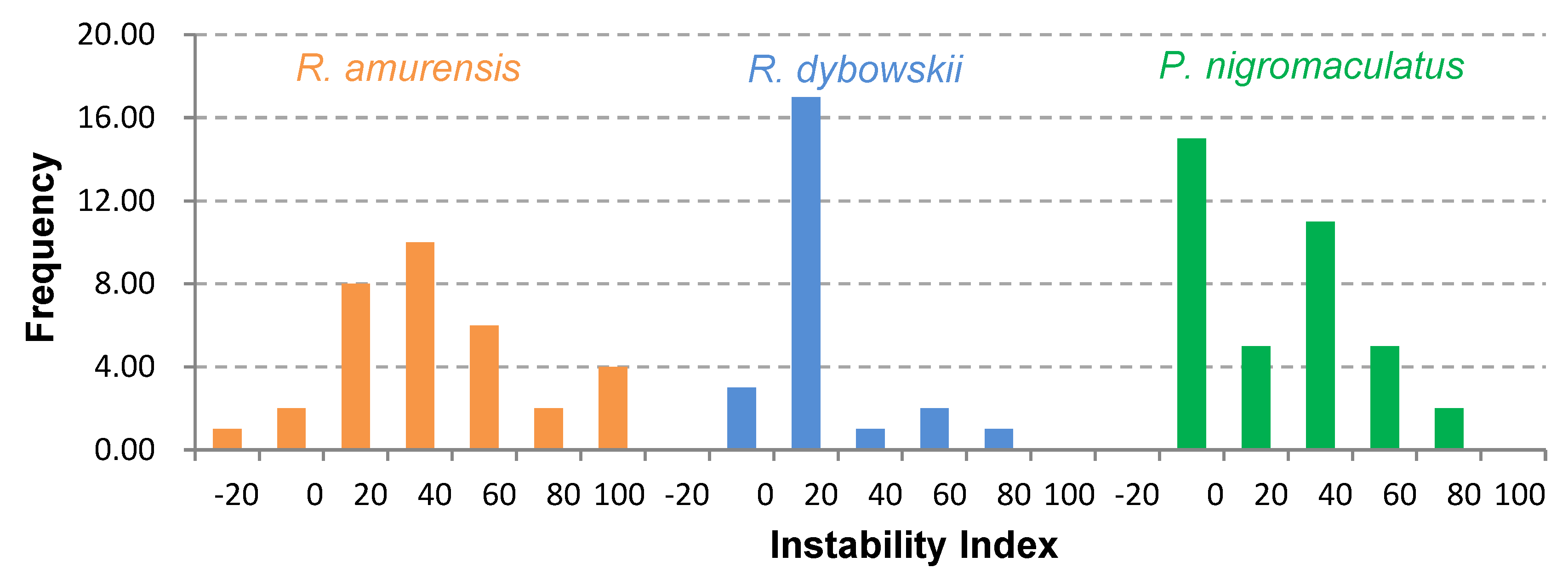
3.4. Prediction of Physicochemical Properties and Secondary Structure of Antimicrobial Peptides (AMPs)
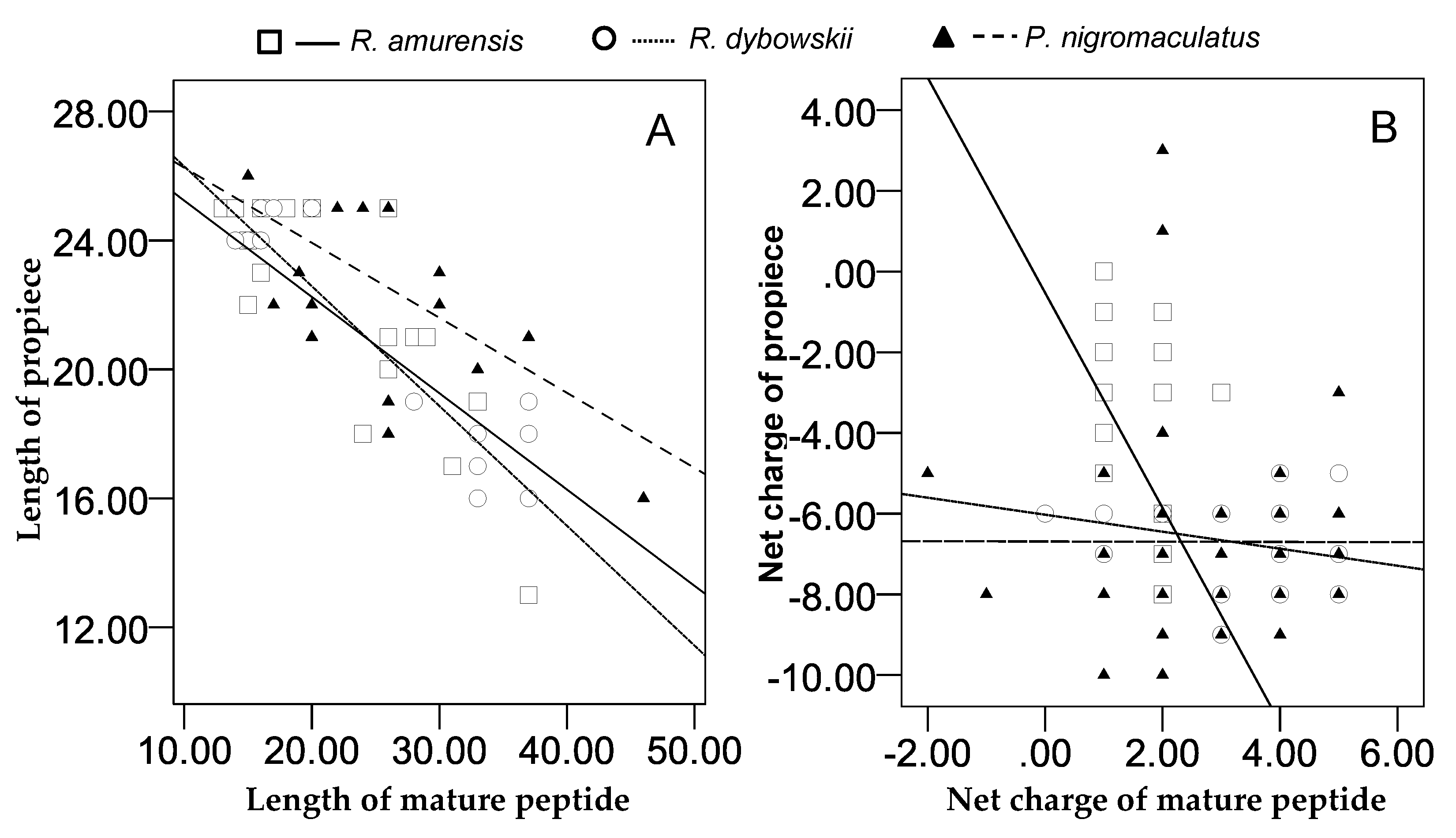
3.5. Relationship between Propieces and Mature Peptides
3.6. Pattern of Nucleotide Substitution
3.6.1. ts/tv Ratio of Propieces to Mature Peptides
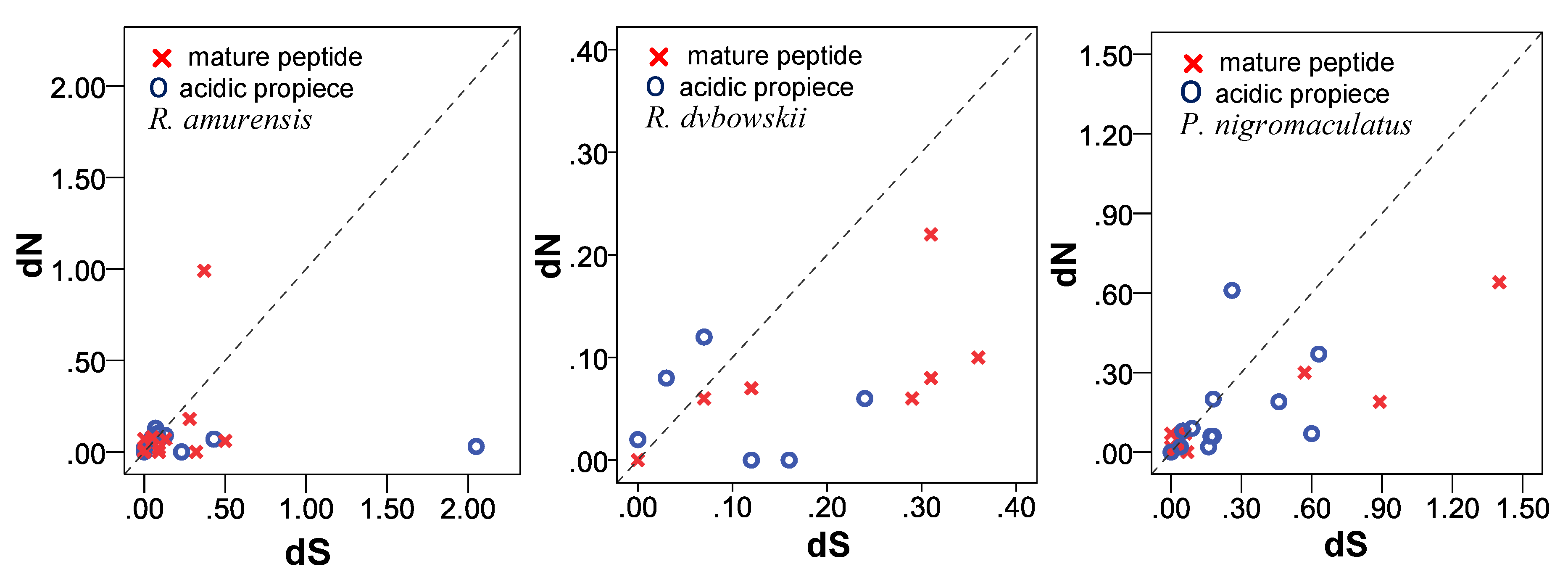
3.6.2. dN/dS Ratio of Propieces and Mature Peptides Region
4. Discussion
4.1. Interspecific Variation of antimicrobial peptide (AMP) Composition
4.2. Correlation between Propieces and Mature Peptides
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO. | AF | MW | pI | Mature Peptides | II | GRAVY | SS |
|---|---|---|---|---|---|---|---|
| 1 | Ranacyclin-2AM | 3823.50 | 10.31 | LAKALGANKVLGQIVRLANSAARDTTQNVLSAITCKI | 0.85 | 0.38 | all-α |
| CHHHHCCHHHHHHHHHHHHHHHHHHHHHHHHHHHCCC | |||||||
| 3857.50 | 10.31 | LAKALGANKVLGQIVRFANSAARDTTQNVLSAITCKI | 0.85 | 0.35 | all-α | ||
| CHHHHCCHHHHHHHHHHHHHHHHHHHHHHHHHHHCCC | |||||||
| 2 | Brevinin-2AM | 3351.00 | 9.24 | GLMDSLKGLAATAGKTVLQGLLKTASCKLEKTC | 22.43 | 0.27 | all-α |
| CCHHHHHHHHHHHHHHHHHHHHHHHHHHCCCCC | |||||||
| 3 | Palustrin-2AM | 3367.00 | 9.51 | GIWDSIKTFGKKFALNIMDKIKCKIGGGCPP | 35.26 | −0.03 | all-α |
| CHHHHHHHHCHHHHHHHHHHHHCCCCCCCCC | |||||||
| 4 | Amurin-6AM | 2872.50 | 9.90 | SLLGLLKGAGKTLLSAGLNKIACKLTGKC | 9.55 | 0.60 | all-α |
| CHHHHHHHHHHHHHHHCCHHHHHHHCCCC | |||||||
| 3007.70 | 9.51 | GLMDFLKGAGKKLLAAGLDKLNCKLTGKC | −23.21 | 0.18 | all-α | ||
| CCCHHHHHHCHHHHHHCCCCCCEEECCCC | |||||||
| 5 | Amurin-5AM | 3114.60 | 8.71 | ALPFLPSLDADIVQQVCCLPRRRCTNGK | 85.13 | 0.01 | mixed |
| CCCCCCCCCCHHHHHHHHCCCCCCCCCC | |||||||
| 3015.50 | 8.00 | ALPFLPSLDADIVQQVCCLPRGRCTNGK | 41.19 | 0.16 | mixed | ||
| CCCCCCCCCCHHHHHHHHCCCCCCCCCC | |||||||
| 3015.50 | 8.00 | ALPFLPSLDADIVQQVCCLPRRGCTNGK | 61.64 | 0.16 | mixed | ||
| CCCCCCCCCCHHHHHHHHCCCCCCCCCC | |||||||
| 3104.60 | 8.71 | ALPFLPSLDADIVQQVCCLSRRRCTNGK | 87.83 | 0.04 | mixed | ||
| CCCCCCCCCCHHHHHHHHCCCCCCCCCC | |||||||
| 3130.70 | 8.71 | ALPFLPSLDADIVQQVCCLLRRRCTNGK | 87.83 | 0.21 | all-α | ||
| CCCCCCCCCHHHHHHHHHHHHCCCCCCC | |||||||
| 3056.60 | 9.22 | ALPFLPSLGADIVQQVCCLPRRRCTNGK | 82.10 | 0.13 | mixed | ||
| CCCCCCCCCCHHHHHHHHCCCCCCCCCC | |||||||
| 6 | Amurin-3AM | 2947.50 | 8.70 | FLPGLKRCCFEVMSSVFCAITRRPDA | 77.14 | 0.45 | mixed |
| CCCCCHHHHHHHHHHHHHEECCCCCC | |||||||
| 2919.50 | 8.68 | FLPGLKRCCFEVMSSVFCAITRKPDA | 44.80 | 0.47 | mixed | ||
| CCCCHHHHHHHHHHHHHHEECCCCCC | |||||||
| 2816.40 | 7.95 | FLPGLKGCCFEVMSSVFCAIPRKPDA | 24.99 | 0.59 | mixed | ||
| CCCCCCHHHHHHHHHHHHEECCCCCC | |||||||
| 2788.30 | 7.94 | FLPGLKGCCFEVMSSVFCAIPKKPDA | 27.89 | 0.62 | mixed | ||
| CCCCCCHHHHHHHHHHHHECCCCCCC | |||||||
| 7 | Amurin-8AM | 2828.50 | 9.51 | GFVGKYMGKISADHLKMIICKVTGKC | 6.40 | 0.33 | mixed |
| CCCHHHCCCCCHHHHHHHHHEECCCC | |||||||
| 8 | Amurin-4AM | 2621.10 | 8.59 | EAKINFAEMFAAVKKILQQGVPMG | 29.56 | 0.26 | all-α |
| CCCCCHHHHHHHHHHHHHHCCCCC | |||||||
| 2646.10 | 9.70 | EAKINFAEMFAAVKKILQQGVPRG | 26.03 | -0.01 | all-α | ||
| CCCCCHHHHHHHHHHHHHHCCCCC | |||||||
| 2603.10 | 8.59 | EAKINFAEIFAAVKKILQQGVPMG | 37.59 | 0.37 | all-α | ||
| CCCCCHHHHHHHHHHHHHHCCCCC | |||||||
| 2611.10 | 10.00 | EAKINFAKIFAAVKKILHQGVPMG | 4.43 | 0.36 | all-α | ||
| CCCCCHHHHHHHHHHHHHHCCCCC | |||||||
| 9 | Ranalexin-1AM | 2286.90 | 8.90 | FFPLLLSALPSFLCLVFKKC | 40.51 | 1.62 | all-α |
| CCHHHHHHHHHHHHHHHHCC | |||||||
| 2252.80 | 8.90 | FLPLLLSALPSFLCLVFKKC | 40.51 | 1.67 | all-α | ||
| CCHHHHHHHHHHHHHHHHCC | |||||||
| 10 | Amurin-7AM | 2456.80 | 12.52 | SLGRFQGRFGRRTHRKHFVN | 32.95 | -1.31 | mixed |
| CCCCCCCCCCCCCHHHHHCC | |||||||
| 11 | Japonicin-1AMa | 2025.50 | 9.39 | FLPPTLVGLAFCKLFKKC | 13.06 | 1.03 | all-α |
| CCCCHHHHHHHHHHHHHC | |||||||
| 1517.90 | 9.39 | FLGLAFCKLFKKC | −8.86 | 1.12 | all-α | ||
| CCCCHHHHHHHHC | |||||||
| 12 | Amurin-9AM | 1745.10 | 9.39 | FLPFFAACAITKKCGK | 55.61 | 0.79 | all-α |
| CCCHHHHHHHHHHHCC | |||||||
| 13 | Temporin –Ama | 1705.10 | 8.75 | ILPLLGNLLNDLLRK | 42.19 | 0.68 | all-α |
| CCCHHHHHHHHHHHC | |||||||
| 1639.10 | 11.00 | FLPVLISLIGRLLGK | 29.35 | 1.56 | all-α | ||
| CCHHHHHHHHHHHCC | |||||||
| 1487.80 | 8.75 | FVSLLTNLLGLLGK | −7.51 | 1.44 | all-α | ||
| CHHHHHHHHHHHCC | |||||||
| 14 | Amurin-10AM | 1537.80 | 9.75 | FLGSILGSLASLFRG | 16.51 | 1.27 | all-α |
| CCHHHHHHHHHHHCC | |||||||
| 1640.90 | 12.00 | FLGSILGSRAFLFRG | 29.35 | 0.95 | mixed | ||
| CCCHHHCCCEEEECC | |||||||
| 1597.90 | 9.75 | FLGSILGSLAFLFRG | 16.51 | 1.51 | all-α | ||
| CCHHHHHHHHHHHCC |
| NO. | AF | MW | pI | Mature Peptides | II | GRAVY | SS |
|---|---|---|---|---|---|---|---|
| 1 | Chensirin-3 | 3732.46 | 9.51 | GLFSEVKGVLKGVGKNVAKNVAGSLLEQLKCKISGGC | 6.64 | 0.27 | all-α |
| CCHHHHHHHHHHHCHHHHHHHHHHHHHHHHCHCCCCC | |||||||
| 3674.42 | 9.79 | GLFSVVKGVLKAAGKNVAKNVGGSLLEQLKCKISGGC | 7.06 | 0.41 | all-α | ||
| CCHHHHHHHHHHHCHHHHHHHHHHHHHHHHCHCCCCC | |||||||
| 3702.48 | 9.79 | GLFSVVKGVLKGVGKNVAKNVAGSLLEQLKCKISGGC | 1.43 | 0.47 | all-α | ||
| CCHHHHHHHHHHHCHHHHHHHHHHHHHHHHCHCCCCC | |||||||
| 3702.48 | 9.79 | GLFSVVKGVLKAVGKNVAKNVGGSLLEQLKCKISGGC | 4.76 | 0.47 | all-α | ||
| CCHHHHHHHHHHHCHHHHHHHHHHHHHHHHCHCCCCC | |||||||
| 3289.99 | 9.60 | GLFSVVKGVLKGVGKNVAGSLLEQLKCKISGGC | 0.39 | 0.57 | all-α | ||
| CCCHHHHHHHHHHHHHHHHHHHHHHHCHCCCCC | |||||||
| 3319.97 | 9.24 | GLFSVVKGVLKGEGKNVAGSLLEQLKCKISGGC | 0.68 | 0.34 | all-α | ||
| CCCCHHHHHHHHCCHHHHHHHHHHHHCHCCCCC | |||||||
| 3213.89 | 9.60 | GILSAVKGVLKGVGKNVAGSLLDQLKCKLSGGC | −2.18 | 0.53 | all-α | ||
| CCHHHHHHHHHHHHHHHHHHHHHHHHHHHCCCC | |||||||
| 3289.99 | 9.60 | GLLSVFKGVLKGVGKNVAGSLLDQLKCKISGGC | −9.13 | 0.56 | all-α | ||
| CCHHHHHHHHHHHHHHHHHHHHHHHHCHECCCC | |||||||
| 3275.96 | 9.60 | GLFSVVKGVLKGVGKNVAGSLLDQLKCKISGGC | −5.45 | 0.57 | all-α | ||
| CCCHHHHHHHHHHHHHHHHHHHHHHHCHECCCC | |||||||
| 2 | Dybowsin-1 | 2908.64 | 10.07 | GLMGILKVAVNKLLAAGMNKPRCKAAHC | 12.89 | 0.46 | all-α |
| CCCCHHHHHHHHHHHHCCCCHHHCCCCC | |||||||
| 3000.69 | 9.70 | GLMDIFKVAVNKLLAAGMNKPRCKAAHC | 6.71 | 0.31 | all-α | ||
| CCCCHHHHHHHHHHHHCCCCHHHCCCCC | |||||||
| 2966.68 | 9.70 | GLMDILKVAVNKLLAAGMNKPRCKAAHC | 15.92 | 0.35 | all-α | ||
| CCCHHHHHHHHHHHHHCCCCHHHCCCCC | |||||||
| 3 | Dybowsin-2 | 2361.93 | 9.39 | FFSLAFAGFPKFLCFLFKKC | 11.62 | 1.26 | mixed |
| CCCCCCCCCCHHHHHHHHHC | |||||||
| 2312.00 | 9.39 | FLPLFFAALPKFLCLVLKKC | 34.15 | 1.60 | all-α | ||
| CCHHHHHHHHHHHHHHHHCC | |||||||
| 2235.82 | 9.39 | FLSLALAGFSKFLCLVFKKC | 11.62 | 1.47 | all-α | ||
| CCCHHHHHHHHHHHHHHHCC | |||||||
| 2269.83 | 9.39 | FFSLALAGFSKFLCLVFKKC | 11.62 | 1.42 | all-α | ||
| CCCHHHHHCHHHHHHHHHCC | |||||||
| 4 | Dybowsin-3 | 1910.42 | 8.96 | FLIGMTQGLICLITRKC | 14.61 | 1.21 | mixed |
| CCCCCCHHHHHHHECCC | |||||||
| 5 | Chensirin-1 | 1611.94 | 5.81 | VFPLVGNLLNDLLGK | 5.19 | 0.89 | all-α |
| CCCCHHHHHHHHHCC | |||||||
| 1577.93 | 5.81 | VLPLVGNLLNDLLGK | 5.19 | 0.96 | all-α | ||
| CCCCHHHHHHHHHCC | |||||||
| 6 | Amurin-3. | 1487.93 | 8.75 | ILPILAPLIGGLLGK | 63.26 | 1.73 | all-α |
| CCCHHHHHHHHHCCC | |||||||
| 1406.82 | 8.75 | ILPILSLIGGLLGK | 53.31 | 1.79 | all-α | ||
| CCHHHHHHHHHCCC | |||||||
| 7 | Chensirin-2 | 1450.78 | 9.70 | IIPLSLGYFAKKT | 0.48 | 0.66 | mixed |
| CCCCCHHHCECCC | |||||||
| 1476.82 | 9.53 | IIPLPLGYYAKKT | 43.01 | 0.29 | mixed | ||
| CCCCCCCCCCCCC | |||||||
| 1460.82 | 9.70 | IIPLPLGYFAKKT | 15.30 | 0.60 | mixed | ||
| CCCCCCCCCCCCC |
| NO. | AF | MW | pI | Mature Peptides | II | GRAVY | SS |
|---|---|---|---|---|---|---|---|
| 1 | Esculentin-1 | 4816.84 | 9.63 | GIFSKLAGKKIKNLLISGLKNIGKEVGMDVVRTGIDIAGCKIKGEC | 3.25 | 0.23 | mixed |
| CCCCCCCCCHHHHHHHHHHHHHHHHHHHHHHHHCCCECCEEECCCC | |||||||
| 4876.94 | 9.63 | GIFFKLAGKKIKNLLISGLKNIGKEVGMDVVRTGIDIAGCKIKGEC | −0.01 | 0.30 | mixed | ||
| CCCCCCCCCHHHHHHHHHHHHHHHHHHHHHHHHCCCCCCEEECCCC | |||||||
| 4756.75 | 9.63 | GISSKLAGKKIKNLLISGLKNIGKEVGMDVVRTGIDIAGCKIKGEC | 7.44 | 0.15 | mixed | ||
| CCCCCCCCHHHHHHHHHHHHHHHHHHCHHHHHHCCCECCEEECCCC | |||||||
| 4802.82 | 9.63 | GIFSKLAGKKIKNLLISGLKNVGKEVGMDVVRTGIDIAGCKIKGEC | −8.14 | 0.22 | mixed | ||
| CCCCCCCCCHHHHHHHHHHHHHHHHHHHHHHHHCCCECCEEECCCC | |||||||
| 4869.89 | 10.02 | GIFSKLAGKKIKNLLISGLKNIGKEVGMDVVRTGIDIAGCKIKGER | −6.30 | 0.07 | mixed | ||
| CCCCCCCCCHHHHHHHHHHHHHHHHHCHHHHHHCCCECCEEEECCC | |||||||
| 2 | Esculentin-2 | 3782.47 | 9.60 | GFLSLLSNAAKFLGKTLLKNVGKAGLETAACKATNQC | 9.60 | 0.27 | all-α |
| CHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHCCCC | |||||||
| 3794.68 | 9.93 | GIFSLIKGAAKVVAKGLGKKVGKFGLDLMACKVTNQC | −10.73 | 0.51 | all-α | ||
| CCCHHHHHHHHHHHHHHHHHHHHHHHHHHHEECCCCC | |||||||
| 3795.62 | 9.51 | GIFSLIKGAAKVVAKGLGKEVGKFGLDLMACKVTNQC | −8.44 | 0.52 | all-α | ||
| CCCHHHHHHHHHHHHHHHHHHHHHHHHHHHHHCCCCC | |||||||
| 3735.52 | 9.51 | GISSLIKGAAKVVAKGLGKEVGKFGLDLMACKVTNQC | −3.23 | 0.43 | all-α | ||
| CCCHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHCCCC | |||||||
| 3602.31 | 9.90 | GALSIFSAAAKLLGKTLLKNAGKAGLQVAACKAANQC | 12.83 | 0.54 | all-α | ||
| CHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHCCC | |||||||
| 3 | Brevinin-2N | 3351.98 | 8.02 | GLMDSLEGLAATAGKTVLQGLLKTASCKLEKTC | 27.57 | 0.28 | all-α |
| CCHHHHHHHHHHHHHHHHHHHHHHHHCCCCCCC | |||||||
| 3351.04 | 9.24 | GLMDSLKGLAATAGKTVLQGLLKTASCKLEKTC | 22.43 | 0.27 | all-α | ||
| CCHHHHHHHHHHHHHHHHHHHHHHHHHHCCCCC | |||||||
| Ranaturein-2N | 3001.64 | 9.24 | GILTDTLKGAAKNVAGVLLDKLKCKITGGC | −10.28 | 0.42 | all-α | |
| CCCHHHHHHHHHHHHHHHHHHCCCEECCCC | |||||||
| 3001.64 | 9.24 | GILKDTLKGAATNVAGVLLDKLKCKITGGC | −18.12 | 0.42 | all-α | ||
| CCCHHHHHHHHHHHHHHHHHHHCCEECCCC | |||||||
| 3012.71 | 9.60 | GILLNTLKGAAKNVAGVLLDKLKCKITGGC | −8.10 | 0.57 | all-α | ||
| CCHHHHHHHHHHHHHHHHHHHCCCEECCCC | |||||||
| 3070.74 | 9.24 | GLLLDTVKGAAKNVAGILLNKLKCKITGDC | −1.80 | 0.47 | all-α | ||
| CCHHHHHHHHHHHHHHHHHHHCCCEECCCC | |||||||
| 3086.70 | 8.79 | GLLLDTDKGAAKNVAGILLNKLKCKITGDC | −3.67 | 0.21 | all-α | ||
| CCCCHHHHHHHHHHHHHHHHHCCCEECCCC | |||||||
| 3056.72 | 9.24 | GLLLDTVKGAAKNVAGILLNKLKCKVTGDC | −4.63 | 0.46 | all-α | ||
| CCHHHHHHHHHHHHHHHHHHHCCCEECCCC | |||||||
| 5 | Nigrocin-6N | 2893.45 | 8.86 | VIPFVASVATEMMHHVYCAASKRCKN | 53.88 | 0.35 | all-α |
| CCCHHHHHHHHHHHHHHHHHHHHCCC | |||||||
| 2921.42 | 8.05 | VIPFVASVATEMMHHVYCAASRRCQN | 67.88 | 0.35 | all-α | ||
| CCCHHHHHHHHHHHHHHHHHHHHHCC | |||||||
| 2882.47 | 9.39 | VIPFVASVAAEMMRHVYCAASKRCKN | 28.95 | 0.40 | all-α | ||
| CCCHHHHHHHHHHHHHHHHHHHHCCC | |||||||
| 2863.42 | 8.86 | VIPFVASVAAEMMHHVYCAASKRCKN | 46.48 | 0.45 | all-α | ||
| CCCHHHHHHHHHHHHHHHHHHHHCCC | |||||||
| 6 | Nigrocin-7N | 2987.60 | 8.50 | ILKCKNELYQLCQVCSLVCWGSSQKL | 24.63 | 0.30 | mixed |
| CCCCHHHHHHHHHHHHEEEECCCCCC | |||||||
| 2977.58 | 8.40 | ILKCKNELYQLCQVCSLVCWGSSQKC | 27.89 | 0.25 | mixed | ||
| CCCCHHHHHHHHHHHHEEEECCCCCC | |||||||
| 3021.62 | 8.50 | ILKCKNELYQLCQVCSLVCWGSSQKF | 27.89 | 0.26 | mixed | ||
| CCCCHHHHHHHHHHHHEEEECCCCCC | |||||||
| 7 | Brevinin-1N | 2513.22 | 9.70 | FLPALIGIAAKALPSLLCKITKKC | 34.61 | 1.12 | all-α |
| CCCHHHHHHHHHHHHHHHHHHHCC | |||||||
| 2513.22 | 9.70 | FLPALIGLAAKALPSLLCKITKKC | 38.15 | 1.09 | all-α | ||
| CCCHHHHHHHHHHHHHHHHHHHCC | |||||||
| 8 | Nigrocin-2N | 2030.56 | 9.39 | GLLSKVLGVGKKVLCGVSGLC | −6.65 | 1.21 | all-α |
| CCCHHHHCCCCEEEECCCCCC | |||||||
| 2000.53 | 9.39 | GLLGKVLGVGKKVLCGVSGLC | −10.69 | 1.23 | all-α | ||
| CCCCHHHCCCCEEEECCCCCC | |||||||
| 9 | Nigrocin-4N | 2244.50 | 5.26 | SLFSISMLKHESSDVENHLA | 24.52 | −0.09 | all-α |
| CCCCEEECCCCCCHHHHHHC | |||||||
| 10 | Ranacyclin-N | 2148.49 | 9.70 | GAPKGCWTKSYPPQPCSGKR | 58.31 | −1.25 | mixed |
| CCCCCCCCCCCCCCCCCCCC | |||||||
| 2148.53 | 9.90 | GAPKGCWTKSYPPKPCSGKR | 35.28 | −1.27 | mixed | ||
| CCCCCCCCCCCCCCCCCCCC | |||||||
| 2247.62 | 10.06 | GAPKGCWTKSYPPQPCSRKR | 72.19 | −1.46 | mixed | ||
| CCCCCCCCCCCCCCCCCCCC | |||||||
| 2176.50 | 9.85 | GAPRGCWTKSYPPQPCSGKR | 54.54 | −1.28 | mixed | ||
| CCCCCCCCCCCCCCCCCCCC | |||||||
| 11 | Nigrocin-5N | 2194.56 | 9.53 | FLPSSPWNEGTYVLKKLKS | 50.12 | −0.48 | mixed |
| CCCCCCCCCCHHHHHHCCC | |||||||
| 12 | Nigrocin-3N | 1798.06 | 4.00 | SPLYTFEGAPGIWPGCC | 38.86 | 0.28 | mixed |
| CCCCEECCCCCCCCCCC | |||||||
| 13 | Temporin-1 | 1555.96 | 8.72 | VIPIVSGLLFSLLGK | −3.91 | 1.83 | all-α |
| CCCHHHHHHHHHHCC | |||||||
| 1495.87 | 8.72 | VIPIVSGLLSSLLGK | 8.93 | 1.59 | all-α | ||
| CCCHHHHHHHHHHCC |
References
- Haslam, I.S.; Roubos, E.W.; Mangoni, M.L.; Yoshizato, K.; Vaudry, H.; Kloepper, J.E.; Pattwell, D.M.; Maderson, P.F.; Paus, R. From frog integument to human skin: dermatological perspectives from frog skin biology. Biol. Rev. Camb. Philos. Soc. 2014, 89, 618–655. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.F.A.; Bui-Marinos, M.P.; Katzenback, B.A. Frog skin innate immune defences: sensing and surviving pathogens. Front. Immunol. 2018, 9, 3128. [Google Scholar] [CrossRef] [PubMed]
- Gill, D.; Nicholas, B.; Aaron, W.; Kisich, K.O. The roles of antimicrobial peptides in innate host defense. Curr. Pharm. Des. 2009, 15, 2377–2392. [Google Scholar] [CrossRef] [Green Version]
- Pukala, T.L.; Bowie, J.H.; Maselli, V.M.; Musgrave, I.F.; Tyler, M.J. Host-defence peptides from the glandular secretions of amphibians: structure and activity. Nat. Prod. Rep. 2006, 23, 368–393. [Google Scholar] [CrossRef]
- Toledo, R.C.; Jared, C. Cutaneous granular glands and amphibian venoms. Comp. Biochem. Physiol. Part A Physiol. 1995, 111, 1–29. [Google Scholar] [CrossRef]
- Woodhams, D.C. Antimicrobial peptide defenses in Amphibian skin. Integr. Comp. Biol. 2005, 45, 137–142. [Google Scholar] [CrossRef] [Green Version]
- Dennison, S.; Wallace, J.; Harris, F.; Phoenix, D. Amphiphilic α-helical antimicrobial peptides and their structure/function relationships. Protein Pept. Lett. 2005, 12, 31–39. [Google Scholar] [CrossRef] [Green Version]
- WimLey, W.C.; Hristova, K. Antimicrobial peptides: successes, challenges and unanswered questions. J. Membr. Biol. 2011, 239, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Chan, D.I.; Prenner, E.J.; Vogel, H.J. Tryptophan- and arginine-rich antimicrobial peptides: structures and mechanisms of action. Biochim. Et Biophys. Acta 2006, 1758, 1184–1202. [Google Scholar] [CrossRef] [Green Version]
- Brogden, K.A. Antimicrobial peptides: pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Kamysz, W. Are antimicrobial peptides an alternative for conventional antibiotics? Nucl. Med. Rev. Cent. East. Eur. 2005, 8, 78–86. [Google Scholar] [PubMed]
- Ladram, A.; Nicolas, P. Antimicrobial peptides from frog skin: biodiversity and therapeutic promises. Front. Biosci. 2016, 21, 1341–1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patocka, J.; Nepovimova, E.; Klimova, B.; Wu, Q.; Kuca, K. Antimicrobial peptides: Amphibian host defense peptides. Curr. Med. Chem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, P.; El Amri, C. The dermaseptin superfamily: A gene-based combinatorial library of antimicrobial peptides. Biochim. Et Biophys. Acta Biomembr. 2009, 1788, 1537–1550. [Google Scholar] [CrossRef] [Green Version]
- Tennessen, J.A. Enhanced synonymous site divergence in positively selected vertebrate antimicrobial peptide genes. J. Mol. Evol. 2005, 61, 445–455. [Google Scholar] [CrossRef]
- Tennessen, J.A. Molecular evolution of animal antimicrobial peptides: widespread moderate positive selection. J. Evol. Biol. 2005, 18, 1387–1394. [Google Scholar] [CrossRef]
- Duda, T.F.; Damien, V.; Pierre, N. Roles of diversifying selection and coordinated evolution in the evolution of amphibian antimicrobial peptides. Mol. Biol. Evol. 2002, 19, 858–864. [Google Scholar] [CrossRef] [Green Version]
- Conlon, J.M.; Iwamuro, S.; King, J.D. Dermal cytolytic peptides and the system of innate immunity in Anurans. Ann. N. Y. Acad. Sci. 2009, 1163, 75–82. [Google Scholar] [CrossRef]
- Conlon, J.M. The contribution of skin antimicrobial peptides to the system of innate immunity in anurans. Cell Tissue Res. 2011, 343, 201–212. [Google Scholar] [CrossRef]
- Ponti, D.; Mignogna, G.M.L.; De, B.D.; Simmaco, M.; Barra, D. Expression and activity of cyclic and linear analogues of esculentin-1, an anti-microbial peptide from Amphibian skin. FEBS J. 2010, 263, 921–927. [Google Scholar] [CrossRef] [Green Version]
- Tennessen, J.A.; Blouin, M.S. Balancing selection at a frog antimicrobial peptide locus: fluctuating immune effector alleles? Mol. Evol. 2008, 25, 2669–2680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanhoye, D.; Bruston, F.; Nicolas, P.; Amiche, M. Antimicrobial peptides from hylid and ranin frogs originated from a 150-million-year-old ancestral precursor with a conserved signal peptide but a hypermutable antimicrobial domain. Eur. J. Biochem. 2003, 270, 2068–2081. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.H.; Li, Y.; Lai, R.; Li, S.; Zhang, Y.; Wang, W. Variety of antimicrobial peptides in the Bombina maxima toad and evidence of their rapid diversification. Eur. J. Immunol. 2005, 35, 1220–1229. [Google Scholar] [CrossRef] [PubMed]
- AmphibiaWeb Provides Information on Amphibian Declines, Natural History, Conservation, and Taxonomy. Available online: http://amphibiaweb.org (accessed on 1 December 2019).
- Zhang, X.L.; Bi, J.-H.; Zhang, X.D.; Ma, Y.-W. New distribution of Rana dybowskii from Inner Mongolia. J. Inn. Mong. Norm. Univ. 2017, 46, 715–716, 721. [Google Scholar] [CrossRef]
- Lee, J.E.; Dong, Y.Y.R.K. Genetic relationships of Rana amurensis based on mitochondrial cytochrome b gene sequences. Korean J. Biol. Sci. 1999, 3, 303–309. [Google Scholar] [CrossRef]
- Ma, D.; Liu, H.-T.; Ji, Y.-J.; Shi, C.-M.; Yang, Y.-H.; Zhang, D.-X. Black-spotted pond frog (Pelophylax nigromaculatus) on the Chinese Loess Plateau represents a cryptic species: evidence from molecular phylogeny and ecological niche modeling. J. Syst. Evol. 2015, 53, 339–350. [Google Scholar] [CrossRef]
- WebLogo is a Web-based Application Designed to Make the Generation of Sequence Logos Easy and Painless. Available online: http://weblogo.threeplusone.com/ (accessed on 4 March 2019).
- Lefort, V.; Longueville, J.E.; Gascuel, O. SMS: smart model selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef] [Green Version]
- Lemoine, F.; Domelevo Entfellner, J.B.; Wilkinson, E.; Correia, D.; Davila Felipe, M.; De Oliveira, T.; Gascuel, O. Renewing Felsenstein’s phylogenetic bootstrap in the era of big data. Nature 2018, 556, 452–456. [Google Scholar] [CrossRef]
- EvolView is an Easy-to-use Online Tool for Displaying, Managing and Customizing Phylogenetic Trees. Available online: https://www.evolgenius.info/evolview/ (accessed on 2 May 2018).
- Amiche, M.; Ladram, A.; Nicolas, P. A consistent nomenclature of antimicrobial peptides isolated from frogs of the subfamily Phyllomedusinae. Peptides 2008, 29, 2074–2082. [Google Scholar] [CrossRef]
- Rozas, J.; Sanchez-DelBarrio, J.C.; Messeguer, X.; Rozas, R. DnaSP, DNA polymorphism analyses by the coalescent and other methods. Bioinformatics 2003, 19, 2496–2497. [Google Scholar] [CrossRef]
- ProtParam is a Tool which Allows the Computation of Various Physical and Chemical Parameters for a Given Protein Stored in Swiss-Prot or TrEMBL or for a User Entered Protein Sequence. Available online: http://www.expasy.org/ (accessed on 12 May 2017).
- PSSpred (Protein Secondary Structure Prediction) is a Simple Neural Network Training Algorithm for Accurate Protein Secondary Structure Prediction. Available online: https://zhanglab.ccmb.med.umich.edu/PSSpred/ (accessed on 5 April 2018).
- PredictProtein: An Open Resource for Online Prediction of Protein Structural and Functional Features. Available online: https://www.predictprotein.org (accessed on 16 January 2012).
- Yang, Z. PAML: A program package for phylogenetic analysis by maximum likelihood. Comput. Appl. Biosci. 1997, 13, 555–556. [Google Scholar] [CrossRef] [PubMed]
- Mor, A.; Hani, K.; Nicolas, P. The vertebrate peptide antibiotics dermaseptins have overlapping structural features but target specific microorganisms. J. Biol. Chem. 1994, 269, 31635–31641. [Google Scholar] [PubMed]
- Yu, G.; Baeder, D.Y.; Regoes, R.R.; Rolff, J. Combination effects of antimicrobial peptides. Antimicrob. Agents Chemother. 2016, 60, 1717–1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kindrachuk, J.; Paur, N.; Reiman, C.; Scruten, E.; Napper, S. The PhoQ-activating potential of antimicrobial peptides contributes to antimicrobial efficacy and is predictive of the induction of bacterial resistance. Antimicrob. Agents Chemother. 2007, 51, 4374–4381. [Google Scholar] [CrossRef] [Green Version]
- Mishra, B.; Wang, G. The importance of amino acid composition in natural AMPs: An evolutional, structural, and functional perspective. Front. Immunol. 2012, 3, 221. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Kizhakkedathu, J.; Straus, S. Antimicrobial peptides: diversity, mechanism of action and strategies to improve the activity and biocompatibility in vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, P.; Rosa, R.D.; Destoumieux-Garzón, D. An intimate link between antimicrobial peptide sequence diversity and binding to essential components of bacterial membranes. Biochim. Et Biophys. Acta BBA Biomembr. 2016, 1858, 958–970. [Google Scholar] [CrossRef]
- Tennessen, J.A.; Blouin, M.S. Selection for antimicrobial peptide diversity in frogs leads to gene duplication and low allelic variation. J. Mol. Evol. 2007, 65, 605–615. [Google Scholar] [CrossRef]
- Wiens, J.J.; Sukumaran, J.; Pyron, R.A.; Brown, R.M. Evolutionary and biogeographic origins of high tropical diversity in old world frogs. Evol. Int. J. Org. Evol. 2009, 63, 1217–1231. [Google Scholar] [CrossRef]
- Ferrer-Admetlla, A.; Bosch, E.; Sikora, M.; Marques-Bonet, T.; Ramirez-Soriano, A.; Muntasell, A.; Navarro, A.; Lazarus, R.; Calafell, F.; Bertranpetit, J.; et al. Balancing selection is the main force shaping the evolution of innate immunity genes. J. Immunol. 2008, 181, 1315–1322. [Google Scholar] [CrossRef] [Green Version]
- Michaelson, D.; Rayner, J.; Couto, M.; Ganz, T. Cationic defensins arise from charge-neutralized propeptides: A mechanism for avoiding leukocyte autocytotoxicity? J. Leukoc. Biol. 1992, 51, 634–639. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.L.; Yeager, M. Coordinated amino acid changes in the evolution of mammalian defensins. J. Mol. Evol. 1997, 44, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Guruprasad, K.; Reddy, B.V.B.; Pandit, M.W. Correlation between stability of a protein and its dipeptide composition: A novel approach for predicting in vivo stability of a protein from its primary sequence. Protein Eng. 1990, 4, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [Green Version]













| Primer | Sequence (5′–3′) | Ta |
|---|---|---|
| PS1 | GGGAC(C/A) ATCAACTTATCTTGT | 54 °C |
| PS2 | GGGACCATCTCCTT(A/G)TCTCTCTGT | 54 °C |
| PS3 | GGGAT(G/C)ATCTCCTTA(A/T)CTCTCTGT | 56 °C |
| PS4 | GGAACCATCTC(A/G)TCTCTCTGT | 55 °C |
| PS5 | GGGACCATCAACTTCTCTCTCTGT | 57 °C |
| PS6 | GGGACCATCAACTT(A/T)TCT(C/T)T(C/T/A)TGT | 55 °C |
| PS7 | GGGACCATCTCCTTATC(T/G)CT(C/T)TG | 54 °C |
| PS8 | GGGACCATCA(G/C)CTTACT(C/T)TGT | 56 °C |
| PS9 | GGGACCATC(G/T)CCCT(T/A)TCTCTCTG | 55 °C |
| PS10 | GGG(A/G)TC(A/G)TCTC(G/C)TTATCTCTCTGT | 55 °C |
| PC | TTTTTTTTTTTTTTTTTTTTTTT(C/G/A) | 50 °C |
| Species | AMP family | ts/tv | ||
|---|---|---|---|---|
| Propiece | Mature Peptide | Whole AMP * | ||
| R. amurensis | Amurin-3AM | 5.31 | 2.60 | 2.43 |
| Amurin-4AM | 1.91 | 1.57 | 1.43 | |
| Amurin-5AM | - | - | 6.13 | |
| Amurin-6AM | 0.08 | 1.28 | 1.37 | |
| Amurin-7AM | 0.69 | 2.04 | 0.63 | |
| Amurin-8AM | 2.27 | - | 1.62 | |
| Amurin-9AM | 0.00 | 1.99 | 0.00 | |
| Amurin-10AM | - | 1.32 | 2.16 | |
| Japonicin-1AM | - | 2.00 | 2.72 | |
| Palustrin-2AM | 6.33 | 1.01 | 1.76 | |
| Ranacyclin-2AM | 2.00 | 0.00 | 0.00 | |
| Ranalexin-1AM | 1.91 | 0.83 | 4.39 | |
| Temporin-AM | 0.09 | 4.04 | 1.32 | |
| Brevinin-AM | / | / | / | |
| R. dybowskii | Amurin-3 | 1.52 | 0.92 | 1.58 |
| Chensirin-1 | - | 1.88 | 1.83 | |
| Chensirin-2 | - | 7.31 | 0.90 | |
| Chensirin-3 | 1.63 | 0.78 | 1.38 | |
| Dybowsin-1 | 0.00 | 2.71 | 2.78 | |
| Dybowsin-2 | - | 3.29 | 1.66 | |
| Dybowsin-3 | - | 2.00 | 3.20 | |
| P. nigromaculatus | Brevinin-1N | 2.00 | 0.00 | 1.77 |
| Brevinin-2N | 0.23 | - | 3.56 | |
| Esculentin-1 | 1.39 | - | 1.70 | |
| Esculentin-2 | 1.17 | 2.08 | 11.32 | |
| Nigrocin-2N | - | - | 1.72 | |
| Nigrocin-3N | 0.91 | - | 2.36 | |
| Nigrocin-5N | 4.73 | 0.00 | 1.38 | |
| Nigrocin-6N | 0.69 | 5.83 | 1.74 | |
| Nigrocin-7N | 0.98 | 3.56 | 3.57 | |
| Ranacyclin-N | - | 1.18 | 1.63 | |
| Ranaturein-2N | 0.68 | 1.22 | 3.04 | |
| Temporin-1 | 2.36 | 12.92 | 2.43 | |
| Nigrocin-4N | / | / | / | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Xia, R.; Ji, J.J.; Zhu, Q.; Li, X.P.; Ma, Y.; Xu, Y.C. Diversity of Antimicrobial Peptides in Three Partially Sympatric Frog Species in Northeast Asia and Implications for Evolution. Genes 2020, 11, 158. https://doi.org/10.3390/genes11020158
Wang Q, Xia R, Ji JJ, Zhu Q, Li XP, Ma Y, Xu YC. Diversity of Antimicrobial Peptides in Three Partially Sympatric Frog Species in Northeast Asia and Implications for Evolution. Genes. 2020; 11(2):158. https://doi.org/10.3390/genes11020158
Chicago/Turabian StyleWang, Qing, Rui Xia, Jing Jing Ji, Qian Zhu, Xiao Ping Li, Yue Ma, and Yan Chun Xu. 2020. "Diversity of Antimicrobial Peptides in Three Partially Sympatric Frog Species in Northeast Asia and Implications for Evolution" Genes 11, no. 2: 158. https://doi.org/10.3390/genes11020158
APA StyleWang, Q., Xia, R., Ji, J. J., Zhu, Q., Li, X. P., Ma, Y., & Xu, Y. C. (2020). Diversity of Antimicrobial Peptides in Three Partially Sympatric Frog Species in Northeast Asia and Implications for Evolution. Genes, 11(2), 158. https://doi.org/10.3390/genes11020158