Serendipitous In Situ Conservation of Faba Bean Landraces in Tunisia: A Case Study

 , , ,
, , ,  ,
,  ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Seed Phenotypic Traits
2.3. DNA Extraction and SSR Assays
2.4. Data Analysis
3. Results
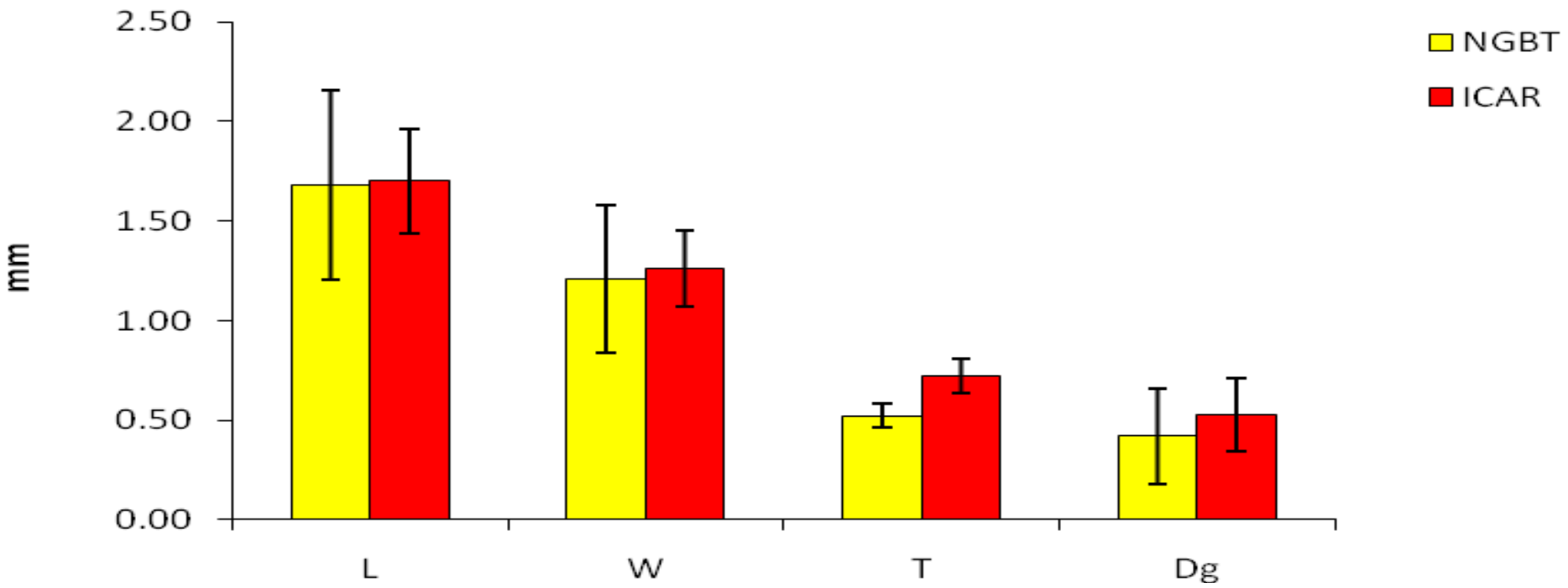
3.1. Variation in Seeds Phenotypic Traits
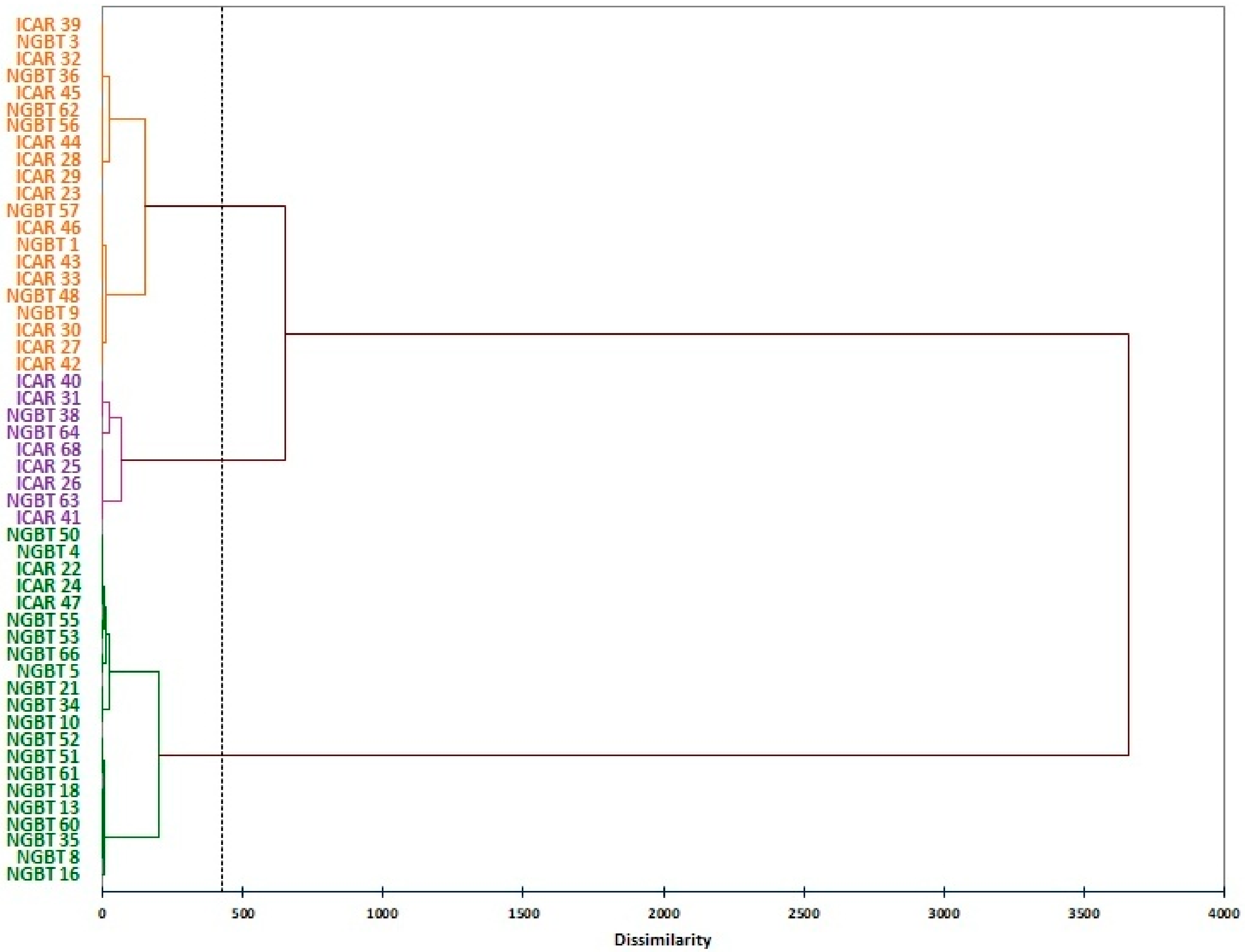
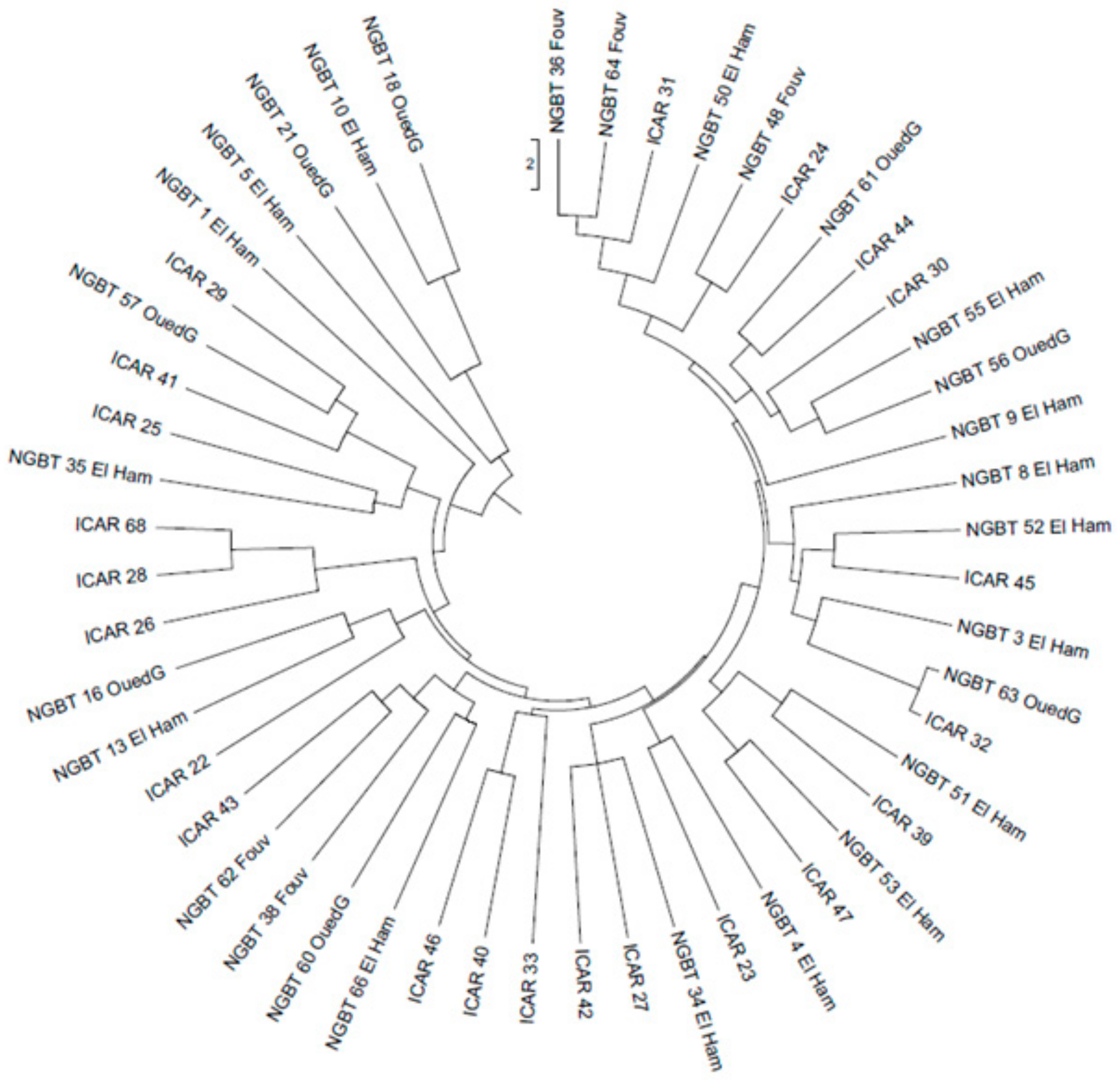
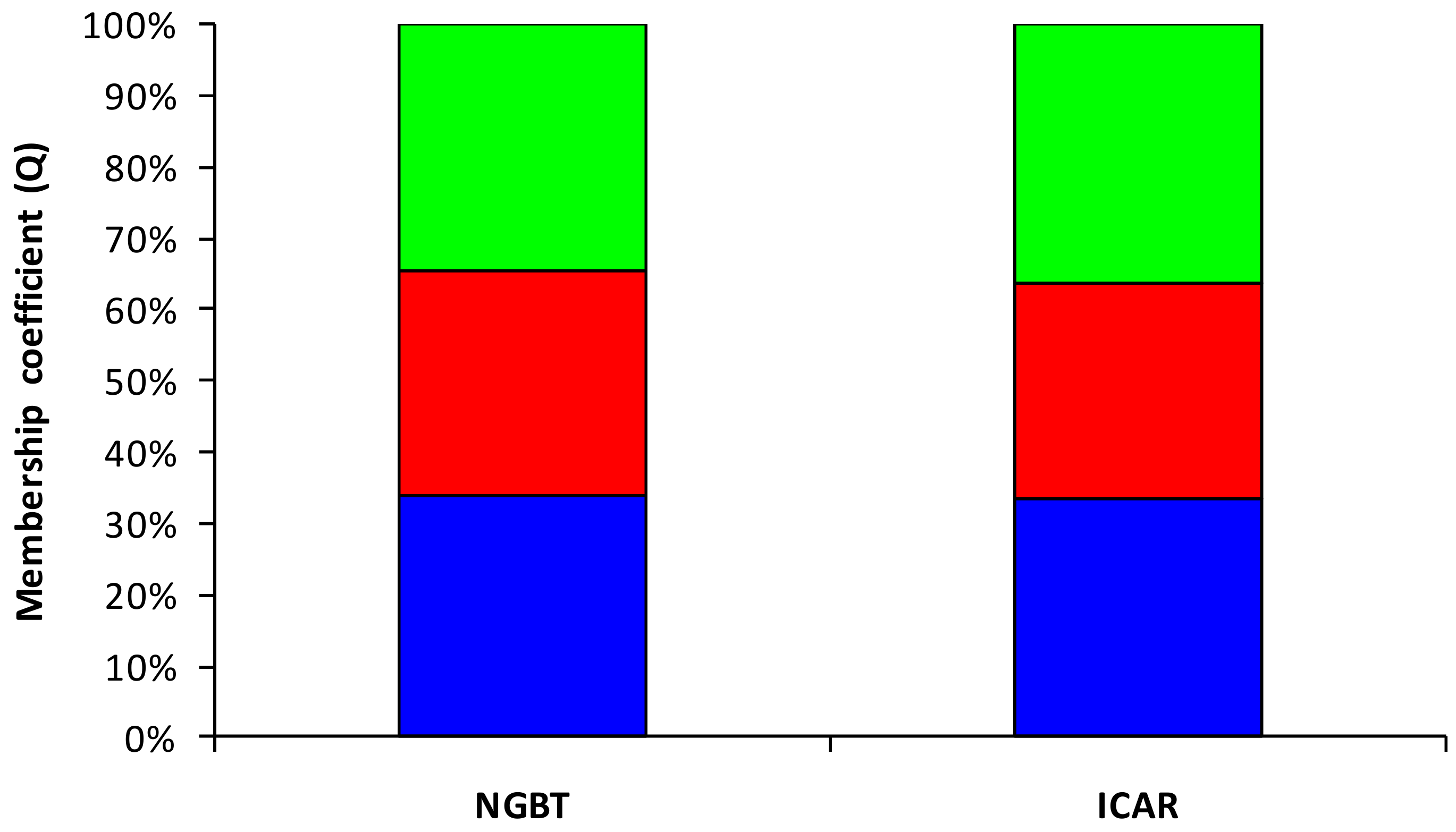
3.2. Molecular Variation of Faba Bean Collection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Leht, M.; Jaaska, V. Cladistic and phenetic analysis of relationships in Vicia subgenus Vicia (Fabaceae) by morphology and isozymes. Plant Syst. Evol. 2002, 232, 237–260. [Google Scholar] [CrossRef]
- Caracuta, V.; Barzilai, O.; Khalaily, H.; Milevski, I.; Paz, Y.; Vardi, J.; Regev, L.; Boaretto, E. The onset of faba bean farming in the Southern Levant. Sci. Rep. 2015, 5, 14370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanno, K.; Willcox, G. The origins of Cicer arietinum L. and Vicia faba L.: Early finds from Tell el Kerkh, north-west Syria, late 10th millennium B.P. Veg. Hist. Archaeobot. 2006, 15, 197–204. [Google Scholar]
- Caracuta, V.; Weinstein-Evron, M.; Kaufman, D.; Yeshurun, R.; Silvent, J.; Boaretto, E. 14,000-year-old seeds indicate the Levantine origin of the lost progenitor of faba bean. Sci. Rep. 2016, 6, 37399. [Google Scholar] [CrossRef] [Green Version]
- Kosterin, O.E. The lost ancestor of the broad bean (Vicia faba L.) and the origin of plant cultivation in the Near East. Vavilov J Genet. PL. BR. 2014, 18, 831–840. [Google Scholar] [CrossRef] [Green Version]
- Hole, F. A Reassessment of the Neolithic Revolution. Paléorien 1984, 10, 49–60. [Google Scholar] [CrossRef]
- Cubero, J.I. On the evolution of Vicia faba L. Theor. Appl. Genet. 1974, 45, 47–51. [Google Scholar] [CrossRef]
- Valderrábano, M.; Gil, T.; Heywood, V.; de Montmollin, B. Conserving Wild Plants in the South and East Mediterranean Region; IUCN: Gland, Switzerland; Málaga, Spain, 2018; p. 146. [Google Scholar]
- Anonymous. Direction Générale de la Production Agricole (DGPA); Ministry of Agriculture Fisheries and Hydraulic Resources of Tunisia: El Ain, Tunisia, 2018. [Google Scholar]
- Rebaa, F.; Abid, G.; Aouida, M.; Abdelkarim, S.; Aroua, I.; Muhovski, Y.; Baudoin, J.P.; M’hamdi, M.; Sassi, K.; Jebara, M. Genetic variability in Tunisian populations of faba bean (Vicia faba L. var. major) assessed by morphological and SSR markers. Physiol. Mol. Biol. Plants 2017, 23, 397–409. [Google Scholar] [CrossRef] [Green Version]
- Kharrat, M.; Ouchari, H. Faba bean status and prospects in Tunisia. Grain Legumes 2011, 56, 11–12. [Google Scholar]
- Hamza, N. La valorization et l’utilisation des ressources génétiques Locals, moyens de leur conservation durable en Tunisie. Ann. de l’INRAT 2ème Numéro Spécial Centen. 2015, 88, 26–32. [Google Scholar]
- Singh, R.K.; Bohra, N.; Sharm, L. Valorizing faba bean for animal feed supplements via biotechnological approach: Opinion. Biocatal. Agric. Biotechnol. 2019, 17, 366–368. [Google Scholar] [CrossRef]
- De Giovanni, C.; Pavan, S.; Taranto, F.; Di Rienzo, V.; Miazzi, M.M.; Marcotrigiano, A.R.; Mangini, G.; Montemurro, C.; Ricciardi, L.; Lotti, C. Genetic variation of a global germplasm collection of chickpea (Cicer arietinum L.) including Italian accessions at risk of genetic erosion. Physiol. Mol. Biol. Plants 2016. [Google Scholar] [CrossRef]
- Oliveira, H.R.; Tomàs, D.; Silva, M.; Viegas, W.; Veloso, M.M. Genetic diversity and population structure in Vicia faba L. landraces and wild related species assessed by nuclear SSRr. PLoS ONE 2016, 11, e015480. [Google Scholar] [CrossRef]
- Liu, Z.; Li, H.; Wen, Z.; Fan, X.; Li, Y.; Guan, R.; Guo, Y.; Wang, S.; Wang, D.; Qiu, L. Comparison of genetic diversity between Chinese and American soybean (Glycine max (L.)) accessions revealed by high-density SNPs. Front. Plant Sci. 2017, 8, 2014. [Google Scholar] [CrossRef] [Green Version]
- Pavan, S.; Bardaro, N.; Fanelli, V.; Marcotrigiano, A.R.; Mangini, G.; Taranto, F.; Catalano, D.; Montemurro, C.; De Giovanni, C.; Lotti, C.; et al. Genotyping by sequencing of cultivated lentil (Lens culinaris Medik.) highlights population structure in the Mediterranean gene pool associated with geographic patterns and phenotypic variables. Front. Genet. 2019, 10, 872. [Google Scholar] [CrossRef]
- Ouji, A.; Suso, M.J.; Rouaissi, M.; Abdellaoui, R.; El Gazzah, M. Genetic diversity of nine faba bean (Vicia faba L.) populations revealed by isozyme markers. Genes Genom. 2011, 33, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Ouji, A.; El Bok, S.; Syed, N.H.; Abdellaoui, R.; Rouaissi, M.; Flavell, A.J.; El Gazzah, M. Genetic diversity of faba bean (Vicia faba L.) populations revealed by sequence specific amplified polymorphism (SSAP) markers. Afr. J. Biotechnol. 2012, 11, 2162–2168. [Google Scholar]
- Yahia, Y.; Hannachi, H.; Monforte, A.J.; Cockram, J.; Loumerem, M.; Zarouri, B.; Ferchichi, A. Genetic diversity in Vicia faba L. populations cultivated in Tunisia revealed by simple sequence repeat analysis. Plant Genet. Res. 2014, 278–285. [Google Scholar] [CrossRef]
- Yahia, Y.; Guetat, A.; Walid, E.; Ferchichi, A.; Yahia, H.; Loumerem, M. Analysis of agromorphological diversity of southern Tunisia faba bean (Vicia faba L.) germplasm. Afr. J. Biotechnol. 2012, 11, 11913–11924. [Google Scholar]
- Dempsey, G.J. In Situ Conservation of Crops and Their Relatives: A Review of Current Status and Prospects for Wheat and Maize; Paper 96-08; CIMMYT NRG: Veracruz, Mexico, 1996. [Google Scholar]
- Damania, A.B. History, achievements, and status of genetic resources conservation. Agron. J. 2008, 100, S-27–S-39. [Google Scholar] [CrossRef]
- Maxted, N.; Bennett, S.J. Conservation, diversity and use of Mediterranean legumes. In Plant Genetic Resources of Legumes in the Mediterranean; Current Plant Science and Biotechnology in Agriculture; Maxted, N., Bennett, S.J., Eds.; Springer: Dordrecht, The Netherlands, 2001; Volume 39. [Google Scholar]
- Altieri, M.A.; Merrick, L. In situ conservation of crop genetic resources through maintenance of traditional farming systems. Econ. Bot. 1987, 41, 41–86. [Google Scholar] [CrossRef]
- Khoury, C.; Laliberté, B.; Guarino, L. Trends in ex situ conservation of plant genetic resources: A review of global crop and regional conservation strategies. Genet. Resour. Crop Evol. 2010, 5, 57–625. [Google Scholar] [CrossRef]
- Enjalbert, J.; Dawson, J.C.; Paillard, S.; Rhoné, B.; Rousselle, Y.; Thomas, M.; Goldringer, I. Dynamic management of crop diversity: From an experimental approach to on-farm conservation. C. R. Biol. 2011, 334, 458–468. [Google Scholar] [CrossRef]
- Mohsenin, N.N. Physical Properties of Plant and Animal Materials; Gordon and Breach Science Publishers: New York, NY, USA, 1970. [Google Scholar]
- Fulton, T.; Chunwongse, J.; Tanksley, S. Microprep protocol for extraction of DNA from tomato and other herbaceous plants. Plant Mol. Biol. Rep. 1995, 13, 207–209. [Google Scholar] [CrossRef]
- Ma, Y.; Yang, T.; Guan, J.; Wang, S.; Wang, H.; Sun, X.; Zong, X. Development and characterization of 21 EST-derived microsatellite markers in Vicia faba (fava bean). Am. J. Bot. 2011, 98, e22–e24. [Google Scholar] [CrossRef]
- Gong, Y.M.; Xu, S.C.; Mao, W.H.; Hu, Q.Z.; Zhang, G.W.; Ding, J.; Li, Z.Y. Generation and characterization of 11 novel est derived microsatellites from Vicia faba (Fabaceae). Am. J. Bot. 2010, 97, 69–71. [Google Scholar] [CrossRef]
- Zeid, M.M.; Mitchell, S.; Link, W.; Carter, W.; Nawar, A.; Fulton, T.; Kresovich, S. Simple sequence repeats (SSRs) in faba bean: New loci from Orobanche-resistant cultivar ‘Giza 402’. Plant Breed. 2009, 128, 149–155. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earl, D.A.; von Holdt, B.M. Structure harvester: A website and program for visualizing structure output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Alghamdi, S.S.; Migdadi, H.M.; Ammar, M.H.; Paull, J.G.; Siddique, K.H.M. Faba bean genomics: Current status and future prospects. Euphytica 2012, 186, 609–624. [Google Scholar] [CrossRef]
- Satovic, Z.; Avila, C.M.; Cruz-Izquierdo, S.; Díaz-Ruíz, R.; García-Ruíz, G.M.; Palomino, C.; Gutiérrez, N.; Vitale, S.; Ocaña-Moral, S.; Gutiérrez, M.V.; et al. A reference consensus genetic map for molecular markers and economically important traits in faba bean (Vicia faba L.). BMC Genom. 2013, 14, 932. [Google Scholar] [CrossRef] [Green Version]
- Torres, A.M.; Avila, C.M.; Gutierrez, N.; Palomino, C.; Moreno, M.T.; Cubero, J.I. Marker-assisted selection in faba bean (Vicia faba L.). Field crops Res. 2010, 115, 243–252. [Google Scholar] [CrossRef]
- Negri, V. Landraces in central Italy: Where and why they are conserved and perspectives for their on-farm conservation. Genet. Resour. Crop Evol. 2003, 50, 871–885. [Google Scholar] [CrossRef]
- Gresta, F.; Avola, G.; Albertini, E.; Raggi, L.; Abbate, V. A study of variability in the Sicilian faba bean landrace ‘Larga di Leonforte’. Genet. Resour. Crop Evol. 2010, 57, 523–531. [Google Scholar] [CrossRef]
- Sun, Y.N.; Pan, J.B.; Shi, X.L.; Du, X.Y.; Wu, Q.; Qi, Z.M.; Jiang, H.W.; Xin, D.W.; Liu, C.Y.; Hu, G.H.; et al. Multi-environment mapping and meta-analysis of 100-seed weight in soybean. Mol. Biol. Rep. 2012, 39, 9435–9443. [Google Scholar] [CrossRef]
- Verma, S.; Gupta, S.; Bandhiwal, N.; Kumar, T.; Bharadwaj, C.; Bhatia, S. High-density linkage map construction and mapping of seed trait QTLs in chickpea (Cicer arietinum L.) using Genotyping-by-Sequencing (GBS). Sci. Rep. 2015, 5, 17512. [Google Scholar] [CrossRef] [Green Version]
- Verma, P.; Goyal, R.; Chahota, R.K.; Sharma, T.R.; Abdin, M.Z.; Bhatia, S. Construction of a genetic linkage map and identification of QTLs for seed weight and seed size traits in lentil, Lens culinaris Medik. PLoS ONE 2015, 10, e0139666. [Google Scholar] [CrossRef] [Green Version]
- Webb, A.; Cottage, A.; Wood, T. A SNP-based consensus genetic map for synteny based trait targeting in faba bean (Vicia faba L.). Plant Biotechnol. J. 2016, 14, 177–185. [Google Scholar]
- Suso, M.J.; Pierre, J.; Moreno, M.T.; Esnault, R.; Le Guen, J. Variation in outcrossing levels in faba bean cultivars: Role of ecological factors. J. Agric. Sci. 2001, 136, 399–405. [Google Scholar] [CrossRef]
- Link, W.; Edered, W.; Metz, P.; Buiel, H.; Melchinger, A.E. Genotypic and environmental variation for degree of cross-fertilization in faba bean. Crop Sci. 1994, 34, 960–964. [Google Scholar] [CrossRef]
- Marzinzig, B.; Brünjes, L.; Biagioni, S.; Behling, H.; Link, W.; Westphal, C. Bee pollinators of faba bean (Vicia faba L.) differ in their foraging behaviour and pollination efficiency. Agric. Ecosyst. Environ. 2018, 264, 24–33. [Google Scholar] [CrossRef]
- Lawson, D.J.; Van Dorp, L.; Falush, D. A tutorial on how not to over-interpret STRUCTURE and ADMIXTURE bar plots. Nat. Commun. 2018, 9, 3258. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Id Name | Local Name | Governorate | Location | Longitude (E) | Latitude (N) | Seed Type |
|---|---|---|---|---|---|---|
| NGBT 1 | Malti | Beja | El Hamra | 9.011641 | 36.52219 | Large |
| NGBT 3 | Malti | ″ | ″ | 9.011641 | 36.52219 | Large |
| NGBT 4 | Malti | ″ | ″ | 9.011641 | 36.52219 | Large |
| NGBT 5 | Chemchali | ″ | ″ | 9.011641 | 36.52219 | Small |
| NGBT 8 | Chemchali | ″ | ″ | 9.011641 | 36.52219 | Small |
| NGBT 9 | ″ | ″ | 9.012502 | 36.52136 | Medium | |
| NGBT 10 | - | ″ | ″ | 9.012502 | 36.52136 | Small |
| NGBT 13 | - | ″ | ″ | 9.012502 | 36.52136 | Small |
| NGBT 34 | - | ″ | ″ | 9.012502 | 36.52136 | Small |
| NGBT 35 | - | ″ | ″ | 9.012502 | 36.52136 | Small |
| NGBT 50 | Chemchali | ″ | ″ | 9.011641 | 36.52219 | Small |
| NGBT 51 | Chemchali | ″ | ″ | 9.011641 | 36.52219 | Small |
| NGBT 52 | Chemchali | ″ | ″ | 9.011641 | 36.52219 | Small |
| NGBT 53 | Chemchali | ″ | ″ | 9.011641 | 36.52219 | Small |
| NGBT 55 | - | ″ | ″ | 9.012502 | 36.52136 | Medium |
| NGBT 66 | - | ″ | ″ | 9.012502 | 36.52136 | Medium |
| NGBT 16 | - | Jendouba | Oued Ghrib | 8.412815 | 36.37286 | Small |
| NGBT 18 | - | ″ | ″ | 8.412815 | 36.37286 | Large |
| NGBT 21 | Bachar | ″ | ″ | 8.414203 | 36.37533 | Small |
| NGBT 56 | - | ″ | ″ | 8.414203 | 36.37533 | Large |
| NGBT 57 | - | ″ | ″ | 8.414203 | 36.37533 | Large |
| NGBT 60 | - | ″ | ″ | 8.412815 | 36.37286 | Small |
| NGBT 61 | - | ″ | ″ | 8.412815 | 36.37286 | Large |
| NGBT 63 | - | ″ | ″ | 8.414203 | 36.37533 | Large |
| NGBT 36 | Malti | ″ | Fouazia | 8.404811 | 36.40103 | Large |
| NGBT 38 | Malti | ″ | ″ | 8.404811 | 36.40103 | Large |
| NGBT 48 | Malti | ″ | ″ | 8.404811 | 36.40103 | Large |
| NGBT 62 | Malti | ″ | ″ | 8.404811 | 36.40103 | Large |
| NGBT 64 | Malti | ″ | ″ | 8.404811 | 36.40103 | Large |
| ICAR 22 | Local | n.a. | n.a. | n.a. | n.a. | Small |
| ICAR 23 | Seville | n.a. | n.a. | n.a. | n.a. | Large |
| ICAR 24 | Misri 32 | Siliana | n.a. | 9.616670 | 36.35000 | Small |
| ICAR 25 | Local | n.a. | n.a. | n.a. | n.a. | Large |
| ICAR 26 | - | Bizerte | n.a. | n.a. | n.a. | Large |
| ICAR 27 | - | Beja | n.a. | n.a. | n.a. | Medium |
| ICAR 28 | - | ″ | n.a. | n.a. | n.a. | Large |
| ICAR 29 | - | n.a. | n.a. | n.a. | n.a. | Large |
| ICAR 30 | Local | Bizerte | n.a. | n.a. | n.a. | Small |
| ICAR 31 | Local | n.a. | n.a. | n.a. | n.a. | Large |
| ICAR 32 | Local | n.a. | n.a. | n.a. | n.a. | Small |
| ICAR 33 | - | n.a. | n.a. | n.a. | n.a. | Small |
| ICAR 39 | Local | Bizerte | n.a. | n.a. | n.a. | Medium |
| ICAR 40 | Malti 24 | ″ | n.a. | 9.666670 | 37.05000 | Medium |
| ICAR 41 | Malti 25 | ″ | n.a. | 9.666680 | 37.06000 | Small |
| ICAR 42 | Misri 39 | Beja | n.a. | 9.583330 | 36.66670 | Small |
| ICAR 43 | Misri 41 | ″ | n.a. | 9.216670 | 36.71670 | Small |
| ICAR 44 | - | ″ | n.a. | n.a. | n.a. | Medium |
| ICAR 45 | - | ″ | n.a. | n.a. | n.a. | Small |
| ICAR 46 | Local | Bizerte | n.a. | n.a. | n.a. | Large |
| ICAR 47 | Local | n.a. | n.a. | n.a. | n.a. | Small |
| ICAR 68 | Local | n.a. | n.a. | n.a. | n.a. | Large |
| N° Samples | Na | I | Ho | He | F | Private Alleles | |
|---|---|---|---|---|---|---|---|
| NGBT | 29 | 7 | 1.428 | 0.621 | 0.658 | 0.027 | 23 |
| ICARDA | 22 | 6 | 1.392 | 0.546 | 0.657 | 0.145 | 12 |
| Whole Collection | 51 | 7 | 1.410 | 0.584 | 0.657 | 0.086 |
| Source of Variation | df | SS | MS | Est. Var. | p-Values |
|---|---|---|---|---|---|
| Among groups | 1 | 3.650 | 3.65 | 0.01 | n.s |
| Within groups | 49 | 161.94 | 3.31 | 3.3 | |
| Total | 50 | 165.59 | 3.31 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Babay, E.; Khamassi, K.; Sabetta, W.; Miazzi, M.M.; Montemurro, C.; Pignone, D.; Danzi, D.; Finetti-Sialer, M.M.; Mangini, G. Serendipitous In Situ Conservation of Faba Bean Landraces in Tunisia: A Case Study. Genes 2020, 11, 236. https://doi.org/10.3390/genes11020236
Babay E, Khamassi K, Sabetta W, Miazzi MM, Montemurro C, Pignone D, Danzi D, Finetti-Sialer MM, Mangini G. Serendipitous In Situ Conservation of Faba Bean Landraces in Tunisia: A Case Study. Genes. 2020; 11(2):236. https://doi.org/10.3390/genes11020236
Chicago/Turabian StyleBabay, Elyes, Khalil Khamassi, Wilma Sabetta, Monica Marilena Miazzi, Cinzia Montemurro, Domenico Pignone, Donatella Danzi, Mariella Matilde Finetti-Sialer, and Giacomo Mangini. 2020. "Serendipitous In Situ Conservation of Faba Bean Landraces in Tunisia: A Case Study" Genes 11, no. 2: 236. https://doi.org/10.3390/genes11020236
APA StyleBabay, E., Khamassi, K., Sabetta, W., Miazzi, M. M., Montemurro, C., Pignone, D., Danzi, D., Finetti-Sialer, M. M., & Mangini, G. (2020). Serendipitous In Situ Conservation of Faba Bean Landraces in Tunisia: A Case Study. Genes, 11(2), 236. https://doi.org/10.3390/genes11020236