Genome-Wide Analysis Reveals Human-Mediated Introgression from Western Pigs to Indigenous Chinese Breeds

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
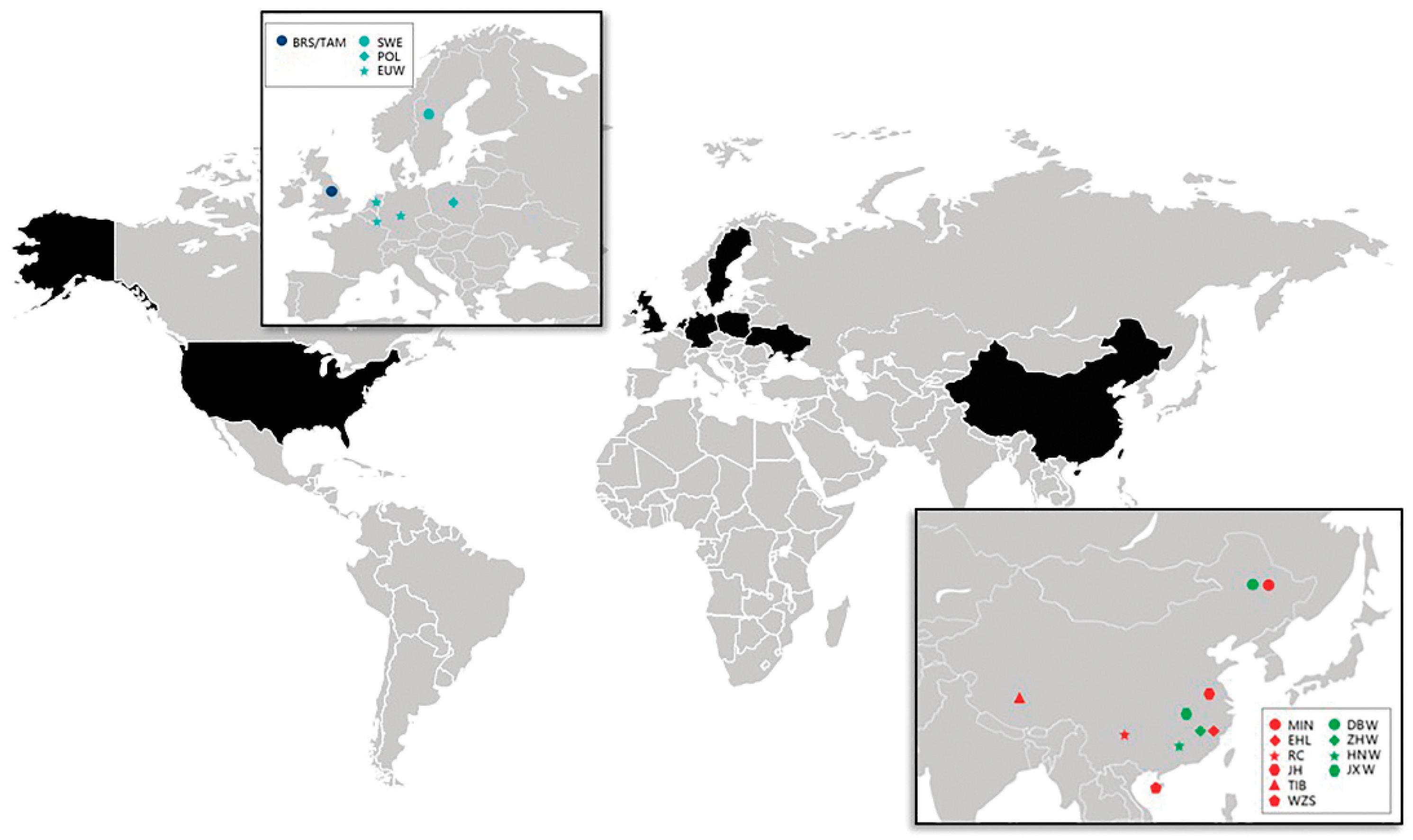
2.1. Sample Collection and Quality Control
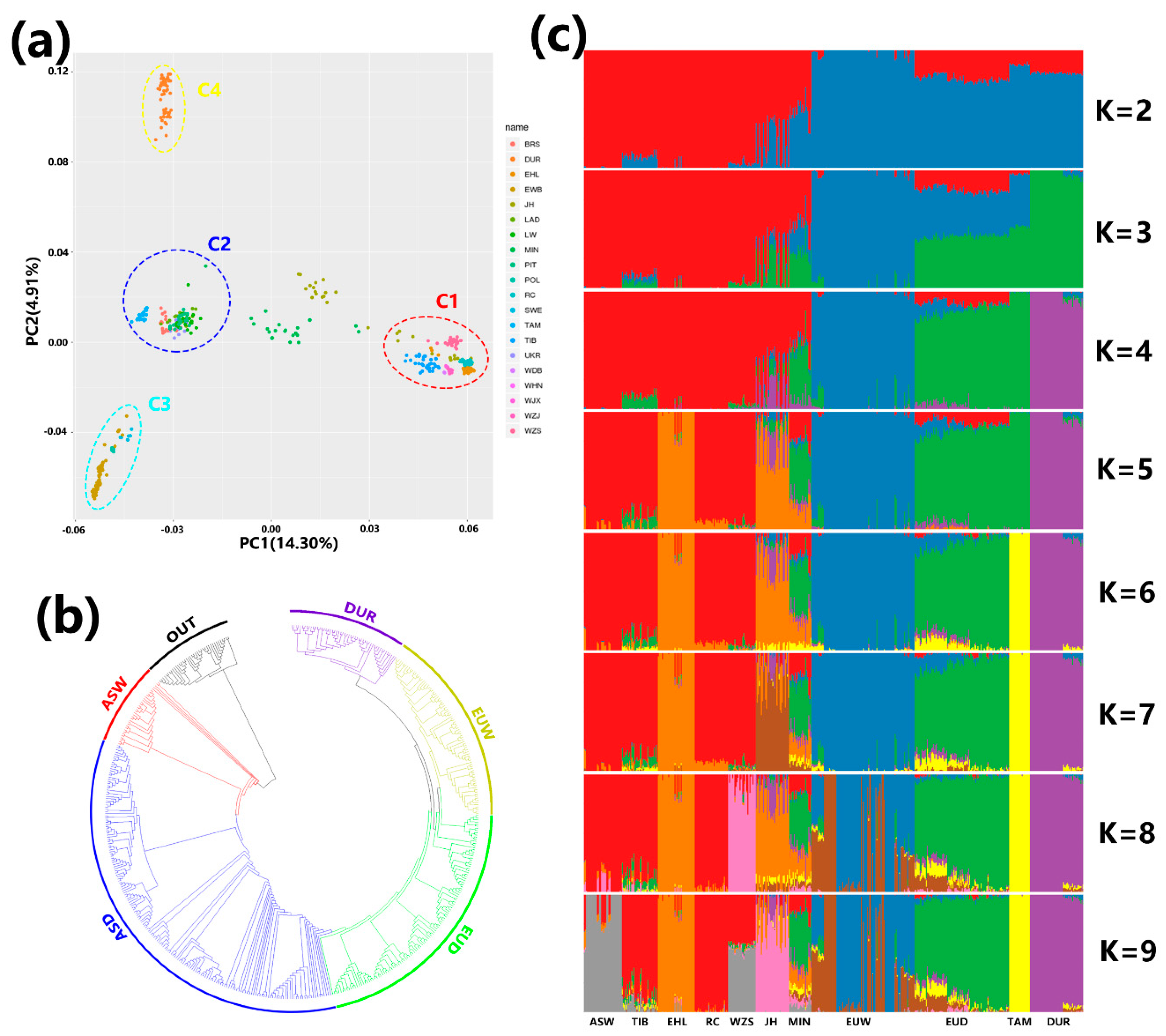
2.2. Population Structure
2.3. Whole-Genome Analysis of Genomic Introgression
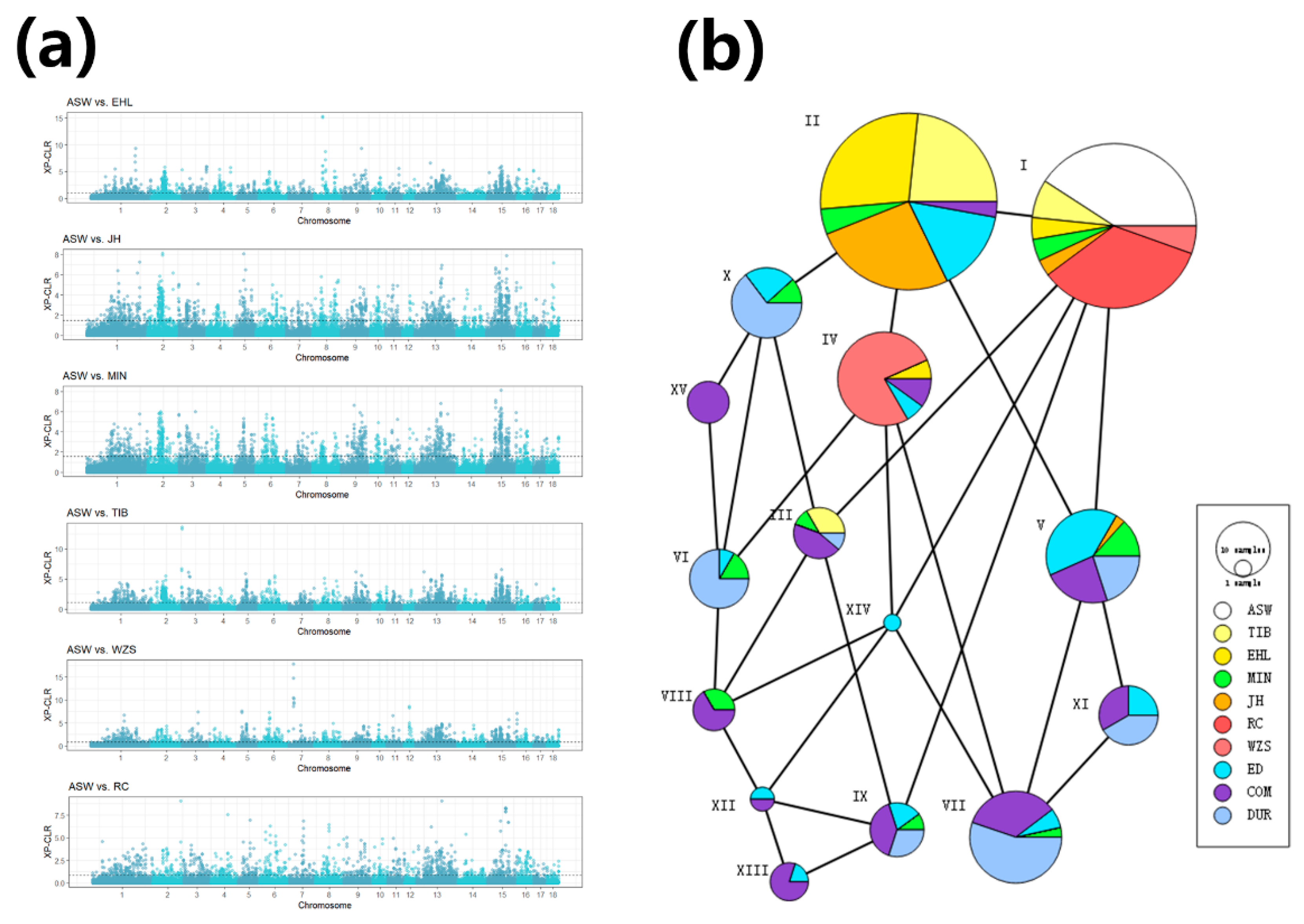
2.4. Selection Scan
3. Results
3.1. Inference of Population Structure and Degree of Breed Admixture
3.2. Introgression from European Pigs to Chinese Indigenous Breeds
3.3. Investigation of Introgressed Regions Affected by Artificial Selection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- United States Department of Agriculture Foreign Agricultural Service (USDA). PSD Online. 2006. Available online: https://apps.fas.usda.gov/psdonline/app/index.html#/app/home (accessed on 1 March 2020).
- United States Department of Agriculture Foreign Agricultural Service (USDA). Swine Summary Selected Countries. Available online: https://www.fsis.usda.gov/wps/wcm/connect/843d74f4-1549-42bf-9af5-3ec3d1c2b213/croatia-2019-far.pdf (accessed on 1 March 2020).
- Darwin, C. The Variation of Animals and Plants under Domestication; Cambridge University Press: Cambridge, UK, 1868; Volume 2. [Google Scholar]
- Ervynck, A.; Dobney, K.; Hongo, H.; Meadow, R. Born Free? New Evidence for the Status of “Sus scrofa” at Neolithic Çayönü Tepesi (Southeastern Anatolia, Turkey). Paléorient 2001, 1, 47–73. [Google Scholar] [CrossRef]
- Epstein, H.; Bichard, M. Pig. In Evolution of Domesticated Animals; Longman: London, UK, 1984. [Google Scholar]
- Cucchi, T.; Hulme-Beaman, A.; Yuan, J.; Dobney, K. Early Neolithic pig domestication at Jiahu, Henan Province, China: Clues from molar shape analyses using geometric morphometric approaches. J. Archaeol. Sci. 2011, 38, 11–22. [Google Scholar] [CrossRef]
- Ek, W.; Marklund, S.; Ragavendran, A.; Siegel, P.B.; Muir, W.M.; Carlborg, O. Generation of a multi-locus chicken introgression line to study the effects of genetic interactions on metabolic phenotypes in chickens. Front. Genet. 2012, 3, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frantz, L.A.; Schraiber, J.G.; Madsen, O.; Megens, H.-J.; Cagan, A.; Bosse, M.; Paudel, Y.; Crooijmans, R.P.; Larson, G.; Groenen, M.A. Evidence of long-term gene flow and selection during domestication from analyses of Eurasian wild and domestic pig genomes. Nat. Genet. 2015, 47, 1141–1148. [Google Scholar] [CrossRef]
- Hu, X.; Yang, J.; Xie, X.; Lv, F.; Cao, Y.; Li, W.; Liu, M.; Wang, Y.; Li, J.; Liu, Y.; et al. The genome landscape of tibetan sheep reveals adaptive introgression from argali and the history of early human settlements on the qinghai-tibetan plateau. Mol. Biol. Evol. 2019, 36, 283–303. [Google Scholar] [CrossRef] [Green Version]
- Leroy, G.; Kayang, B.B.; Youssao, I.; Yapignaore, C.V.; Oseiamponsah, R.; Loukou, N.E.; Fotsa, J.C.; Benabdeljelil, K.; Bedhom, B.; Tixierboichard, M.; et al. Gene diversity, agroecological structure and introgression patterns among village chicken populations across North, West and Central Africa. BMC Genet. 2012, 13, 34. [Google Scholar] [CrossRef] [Green Version]
- Randi, E.; Hulva, P.; Fabbri, E.; Galaverni, M.; Galov, A.; Kusak, J.; Bigi, D.; Bolfikova, B.C.; Smetanova, M.; Caniglia, R. Multilocus detection of wolf x dog hybridization in Italy, and guidelines for marker selection. PLoS ONE 2014, 9, e86409. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Lin, D.; Wang, Y.; Peng, D.; Li, H.; Fei, J.; Chen, K.; Yang, N.; Hu, X.; Zhao, Y.; et al. Widespread introgression in Chinese indigenous chicken breeds from commercial broiler. Evol. Appl. 2019, 12, 610–621. [Google Scholar] [CrossRef] [Green Version]
- Frantz, A.C.; Zachos, F.E.; Kirschning, J.; Cellina, S.; Bertouille, S.; Mamuris, Z.; Koutsogiannouli, E.A.; Burke, T. Genetic evidence for introgression between domestic pigs and wild boars (Sus scrofa) in Belgium and Luxembourg: A comparative approach with multiple marker systems. Biol. J. Linn. Soc. 2013, 110, 104–115. [Google Scholar] [CrossRef] [Green Version]
- Burgos-Paz, W.; Souza, C.; Megens, H.; Ramayo-Caldas, Y.; Melo, M.; Lemús-Flores, C.; Caal, E.; Soto, H.; Martínez, R.; Alvarez, L.; et al. Porcine colonization of the Americas: A 60k SNP story. Heredity 2013, 110, 321. [Google Scholar] [CrossRef] [Green Version]
- Hanotte, O.; Bradley, D.G.; Ochieng, J.W.; Verjee, Y.; Hill, E.W.; Rege, J.E.O. African pastoralism: Genetic imprints of origins and migrations. Science 2002, 296, 336–339. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Calus, M.P.L.; Bosse, M.; Sahana, G.; Lund, M.S.; Guldbrandtsen, B. Human-mediated introgression of haplotypes in a modern dairy cattle breed. Genetics 2018, 209, 1305–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Wang, H.; Zhang, Y.; Tang, Z.; Li, K.; Liu, B. Genome-wide analysis reveals artificial selection on coat colour and reproductive traits in Chinese domestic pigs. Mol. Ecol. Resour. 2015, 15, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Haley, C.; d’Agaro, E.; Ellis, M. Genetic components of growth and ultrasonic fat depth traits in Meishan and Large White pigs and their reciprocal crosses. Anim. Sci. 1992, 54, 105–115. [Google Scholar] [CrossRef]
- Rothschild, M.; Jacobson, C.; Vaske, D.; Tuggle, C.; Wang, L.; Short, T.; Eckardt, G.; Sasaki, S.; Vincent, A.; McLaren, D.; et al. The estrogen receptor locus is associated with a major gene influencing litter size in pigs. Proc. Natl. Acad. Sci. USA 1996, 93, 201–205. [Google Scholar] [CrossRef] [Green Version]
- Bosse, M.; Megens, H.-J.; Frantz, L.A.; Madsen, O.; Larson, G.; Paudel, Y.; Duijvesteijn, N.; Harlizius, B.; Hagemeijer, Y.; Crooijmans, R.P.; et al. Genomic analysis reveals selection for Asian genes in European pigs following human-mediated introgression. Nat. Commun. 2014, 5, 4392. [Google Scholar] [CrossRef] [Green Version]
- Fang, M.; Andersson, L. Mitochondrial diversity in European and Chinese pigs is consistent with population expansions that occurred prior to domestication. Proc. R. Soc. Lond. B Biol. Sci. 2006, 273, 1803–1810. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Zhu, L.; Tang, G.; Li, M.; Jiang, A.; Cen, W.; Xing, S.; Chen, J.; Wen, A.; He, T.; et al. Carcass and meat quality traits of four commercial pig crossbreeds in China. Genet. Mol. Res. 2012, 11, 4447–4455. [Google Scholar] [CrossRef]
- Yang, S.; Li, X.; Li, K.; Fan, B.; Tang, Z. A genome-wide scan for signatures of selection in Chinese indigenous and commercial pig breeds. BMC Genet. 2014, 15, 7. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.-L.; Wang, Z.-G.; Liu, B.; Zhang, G.-X.; Zhao, S.-H.; Yu, M.; Fan, B.; Li, M.-H.; Xiong, T.-A.; Li, K. Genetic variation and relationships of eighteen Chinese indigenous pig breeds. Genet. Sel. Evol. 2003, 35, 657. [Google Scholar] [CrossRef] [Green Version]
- Ramos, A.M.; Crooijmans, R.P.; Affara, N.A.; Amaral, A.J.; Archibald, A.L.; Beever, J.E.; Bendixen, C.; Churcher, C.; Clark, R.; Dehais, P.; et al. Design of a high density SNP genotyping assay in the pig using SNPs identified and characterized by next generation sequencing technology. PLoS ONE 2009, 4, e6524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goedbloed, D.; Megens, H.; Van Hooft, P.; Herrero-Medrano, J.; Lutz, W.; Alexandri, P.; Crooijmans, R.; Groenen, M.; Van Wieren, S.; Ydenberg, R.; et al. Genome-wide single nucleotide polymorphism analysis reveals recent genetic introgression from domestic pigs into Northwest European wild boar populations. Mol. Ecol. 2013, 22, 856–866. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Cui, L.; Perez-Enciso, M.; Traspov, A.; Crooijmans, R.P.; Zinovieva, N.; Schook, L.B.; Archibald, A.; Gatphayak, K.; Knorr, C.; et al. Genome-wide SNP data unveils the globalization of domesticated pigs. Genet. Sel. Evol. 2017, 49, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Lee, S.H.; Goddard, M.E.; Visscher, P.M. GCTA: A tool for genome-wide complex trait analysis. Am. J. Hum. Genet. 2011, 88, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef] [Green Version]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Durand, E.Y.; Patterson, N.; Reich, D.; Slatkin, M. Testing for ancient admixture between closely related populations. Mol. Biol. Evol. 2011, 28, 2239–2252. [Google Scholar] [CrossRef] [Green Version]
- Green, R.E.; Krause, J.; Briggs, A.W.; Maricic, T.; Stenzel, U.; Kircher, M.; Patterson, N.; Li, H.; Zhai, W.; Fritz, M.H.-Y.; et al. A draft sequence of the Neandertal genome. Science 2010, 328, 710–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, S.H.; Davey, J.W.; Jiggins, C.D. Evaluating the use of ABBA–BABA statistics to locate introgressed loci. Mol. Biol. Evol. 2014, 32, 244–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malinsky, M. Dsuite-fast D-statistics and related admixture evidence from VCF files. BioRxiv 2019, 634477. [Google Scholar]
- Teng, H.; Zhang, Y.; Shi, C.; Mao, F.; Cai, W.; Lu, L.; Zhao, F.; Sun, Z.; Zhang, J. Population genomics reveals speciation and introgression between brown Norway rats and their sibling species. Mol. Biol. Evol. 2017, 34, 2214–2228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hufford, M.B.; Xu, X.; Van Heerwaarden, J.; Pyhäjärvi, T.; Chia, J.-M.; Cartwright, R.A.; Elshire, R.J.; Glaubitz, J.C.; Guill, K.E.; Kaeppler, S.M.; et al. Comparative population genomics of maize domestication and improvement. Nat. Genet. 2012, 44, 808. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Patterson, N.; Reich, D. Population differentiation as a test for selective sweeps. Genome Res. 2010, 20, 393–402. [Google Scholar] [CrossRef] [Green Version]
- Leigh, J.W.; Bryant, D. popart: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Bandelt, H.-J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Rozas, J.; Sánchez-DelBarrio, J.C.; Messeguer, X.; Rozas, R. DnaSP, DNA polymorphism analyses by the coalescent and other methods. Bioinformatics 2003, 19, 2496–2497. [Google Scholar] [CrossRef]
- The R Development Core Team. R: A Language and Environment for Statistical Computing; The R Foundation: Vienna, Austria, 2013; p. 409. [Google Scholar]
- Wangsheng, X. Introduction and domestication of European breeds of pig in modern China. Anc. Mod. Agric. 2004, 1, 8. [Google Scholar]
- Zhongge, Z.; Bingtan, L.; Xiaohua, C. Chinese pig breeds. Shanghai Sci. Technol. Agency 1986, 48, 14219–14227. [Google Scholar]
- Dmitriev, N.G.; Ernest, L. Animal Genetic Resources of the USSR; FAO: Roma, Italy, 1989. [Google Scholar]
- Cahill, J.A.; Stirling, I.; Kistler, L.; Salamzade, R.; Ersmark, E.; Fulton, T.L.; Stiller, M.; Green, R.E.; Shapiro, B. Genomic evidence of geographically widespread effect of gene flow from polar bears into brown bears. Mol. Ecol. 2015, 24, 1205–1217. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Lorenzen, E.D.; Fumagalli, M.; Li, B.; Harris, K.; Xiong, Z.; Zhou, L.; Korneliussen, T.S.; Somel, M.; Babbitt, C.; et al. Population genomics reveal recent speciation and rapid evolutionary adaptation in polar bears. Cell 2014, 157, 785–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Z.; Silva, P.; Gronau, I.; Wang, S.; Armero, A.S.; Schweizer, R.M.; Ramirez, O.; Pollinger, J.; Galaverni, M.; Del-Vecchyo, D.O.; et al. Worldwide patterns of genomic variation and admixture in gray wolves. Genome Res. 2016, 26, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Skoglund, P.; Ersmark, E.; Palkopoulou, E.; Dalén, L. Ancient wolf genome reveals an early divergence of domestic dog ancestors and admixture into high-latitude breeds. Curr. Biol. 2015, 25, 1515–1519. [Google Scholar] [CrossRef] [Green Version]
- Klasan, G.S.; Ivanac, D.; Erzen, D.J.; Picard, A.; Takasawa, S.; Peharec, S.; Arbanas, J.; Girotto, D.; Jerkovic, R. Reg3G gene expression in regenerating skeletal muscle and corresponding nerve. Muscle Nerve 2014, 49, 61–68. [Google Scholar] [CrossRef]
- Li, X.; Liu, P.; Liu, W.; Maye, P.; Zhang, J.; Zhang, Y.; Hurley, M.; Guo, C.; Boskey, A.; Sun, L.; et al. Dkk2 has a role in terminal osteoblast differentiation and mineralized matrix formation. Nat. Genet. 2005, 37, 945. [Google Scholar] [CrossRef]
- Pekkinen, M.; Terhal, P.A.; Botto, L.D.; Henning, P.; Mäkitie, R.E.; Roschger, P.; Jain, A.; Kol, M.; Kjellberg, M.A.; Paschalis, E.P.; et al. Osteoporosis and skeletal dysplasia caused by pathogenic variants in SGMS2. JCI Insight 2019, 4, 7. [Google Scholar] [CrossRef] [Green Version]
- Steensma, D.P.; Neiger, J.D.; Porcher, J.C.; Keats, J.J.; Bergsagel, P.L.; Dennis, T.R.; Knudson, R.A.; Jenkins, R.B.; Santana-Davila, R.; Kumar, R.; et al. Rearrangements and amplification of IER3 (IEX-1) represent a novel and recurrent molecular abnormality in myelodysplastic syndromes. Cancer Res. 2009, 69, 7518–7523. [Google Scholar] [CrossRef] [Green Version]
- Zweers, M.C.; Bristow, J.; Steijlen, P.M.; Dean, W.B.; Hamel, B.C.; Otero, M.; Kucharekova, M.; Boezeman, J.B.; Schalkwijk, J. Haploinsufficiency of TNXB is associated with hypermobility type of Ehlers-Danlos syndrome. Am. J. Hum. Genet. 2003, 73, 214–217. [Google Scholar] [CrossRef] [Green Version]
- Guan, R.J.; Ford, H.L.; Fu, Y.; Li, Y.; Shaw, L.M.; Pardee, A.B. Drg-1 as a differentiation-related, putative metastatic suppressor gene in human colon cancer. Cancer Res. 2000, 60, 749–755. [Google Scholar] [PubMed]
- Shibata, N.; Kawarai, T.; Lee, J.H.; Lee, H.-S.; Shibata, E.; Sato, C.; Liang, Y.; Duara, R.; Mayeux, R.P.; St George-Hyslop, P.H.; et al. Association studies of cholesterol metabolism genes (CH25H, ABCA1 and CH24H) in Alzheimer’s disease. Neurosci. Lett. 2006, 391, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Bosch, D.G.; Boonstra, F.N.; de Leeuw, N.; Pfundt, R.; Nillesen, W.M.; de Ligt, J.; Gilissen, C.; Jhangiani, S.; Lupski, J.R.; Cremers, F.P.; et al. Novel genetic causes for cerebral visual impairment. Eur. J. Hum. Genet. 2016, 24, 660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, T.; Qian, S.; Shen, S.; Zhang, X.; Liu, J.; Jia, W.; Chen, Z.; Ye, J. Reduction of mitochondrial 3-oxoacyl-ACP synthase (OXSM) by hyperglycemia is associated with deficiency of α-lipoic acid synthetic pathway in kidney of diabetic mice. Biochem. Biophys. Res. Commun. 2019, 512, 106–111. [Google Scholar] [CrossRef]
- Boonstra, N.; Limburg, H.; Tijmes, N.; van Genderen, M.; Schuil, J.; van Nispen, R. Changes in causes of low vision between 1988 and 2009 in a Dutch population of children. Acta Ophthalmol. 2012, 90, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Prüfer, K.; Racimo, F.; Patterson, N.; Jay, F.; Sankararaman, S.; Sawyer, S.; Heinze, A.; Renaud, G.; Sudmant, P.H.; De Filippo, C.; et al. The complete genome sequence of a Neanderthal from the Altai Mountains. Nature 2014, 505, 43–49. [Google Scholar] [CrossRef]
- Sankararaman, S.; Mallick, S.; Dannemann, M.; Prüfer, K.; Kelso, J.; Pääbo, S.; Patterson, N.; Reich, D. The genomic landscape of Neanderthal ancestry in present-day humans. Nature 2014, 507, 354–357. [Google Scholar] [CrossRef] [Green Version]
- Wall, J.D.; Lohmueller, K.E.; Plagnol, V. Detecting ancient admixture and estimating demographic parameters in multiple human populations. Mol. Biol. Evol. 2009, 26, 1823–1827. [Google Scholar] [CrossRef] [Green Version]
- Skov, L.; Hui, R.; Shchur, V.; Hobolth, A.; Scally, A.; Schierup, M.H.; Durbin, R. Detecting archaic introgression using an unadmixed outgroup. PLoS Genet. 2018, 14, e1007641. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Breed | Code | Origin | Number * |
|---|---|---|---|
| Dongbei boar | DBW | Northeast China | 3 |
| Zhejiang boar | ZJW | Zhejiang province, China | 15 |
| South China boar | HNW | South of China | 15 |
| Jiangxi boar | JXW | Jiangxi province, China | 5 |
| Tibetan pig | TIB | Tibet A.R., China | 35 |
| Erhualian pig | EHL | Jiangsu province, China | 36 |
| Jinhua pig | JH | Zhejiang province, China | 44 |
| Min pig | MIN | Northeast China | 22 |
| Rongchang pig | RC | Sichuan province, China | 32 |
| Wuzhishan pig | WZS | Hainan province, China | 27 |
| Polish boar | PLW | Poland | 6 |
| Swedish boar | SWE | Sweden | 6 |
| British Saddleback # | BRS | United Kingdom | 20 |
| Tamworth # | TAM | United Kingdom | 20 |
| Landrace # | LAD | Europe | 20 |
| Large White # | LW | United Kingdom | 20 |
| Pietrain # | PIT | Belgium | 20 |
| Other European boar # | EUW | Europe | 88 |
| Duroc # | DUR | United States | 52 |
| Warthogs + | OUT | Africa | 39 |
| Total | 525 |
| COM | p-Value | ED | p-Value | DUR | p-Value | EW | p-Value | |
|---|---|---|---|---|---|---|---|---|
| EHL | 0.03 | 2.2 × 10−03 | 0.03 | 1.1 × 10−03 | 0.03 | 9.0 × 10−04 | 0.03 | 0.02 |
| TIB | 0.12 | 2.2 × 10−16 | 0.13 | 6.1 × 10−15 | 0.11 | 2.2 × 10−16 | 0.61 | 1.0 × 10−26 |
| WZS | 0.03 | 4.9 × 10−04 | 0.02 | 0.01 | 0.03 | 5.9 × 10−04 | 0.04 | 5.4 × 10−04 |
| RC | 1.9 × 10−03 | 0.43 | 5.3 × 10−03 | 0.30 | 1.4 × 10−05 | 0.49 | 3.8 × 10−03 | 0.07 |
| JH | 0.24 | 1.0 × 10−26 | 0.26 | 1.0 × 10−26 | 0.29 | 1.0 × 10−26 | 0.40 | 1.0 × 10−26 |
| MIN | 0.45 | 1.0 × 10−26 | 0.43 | 1.0 × 10−26 | 0.42 | 1.0 × 10−26 | 0.61 | 1.0 × 10−26 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Liu, C.; Chen, J.; Bai, Y.; Wang, K.; Wang, Y.; Fang, M. Genome-Wide Analysis Reveals Human-Mediated Introgression from Western Pigs to Indigenous Chinese Breeds. Genes 2020, 11, 275. https://doi.org/10.3390/genes11030275
Wang J, Liu C, Chen J, Bai Y, Wang K, Wang Y, Fang M. Genome-Wide Analysis Reveals Human-Mediated Introgression from Western Pigs to Indigenous Chinese Breeds. Genes. 2020; 11(3):275. https://doi.org/10.3390/genes11030275
Chicago/Turabian StyleWang, Jue, Chengkun Liu, Jie Chen, Ying Bai, Kejun Wang, Yubei Wang, and Meiying Fang. 2020. "Genome-Wide Analysis Reveals Human-Mediated Introgression from Western Pigs to Indigenous Chinese Breeds" Genes 11, no. 3: 275. https://doi.org/10.3390/genes11030275
APA StyleWang, J., Liu, C., Chen, J., Bai, Y., Wang, K., Wang, Y., & Fang, M. (2020). Genome-Wide Analysis Reveals Human-Mediated Introgression from Western Pigs to Indigenous Chinese Breeds. Genes, 11(3), 275. https://doi.org/10.3390/genes11030275