Inferring Effective Population Size and Divergence Time in the Lithuanian Population According to High-Density Genotyping Data

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Genotyping
2.3. Ne and Divergence Time Analysis
3. Results
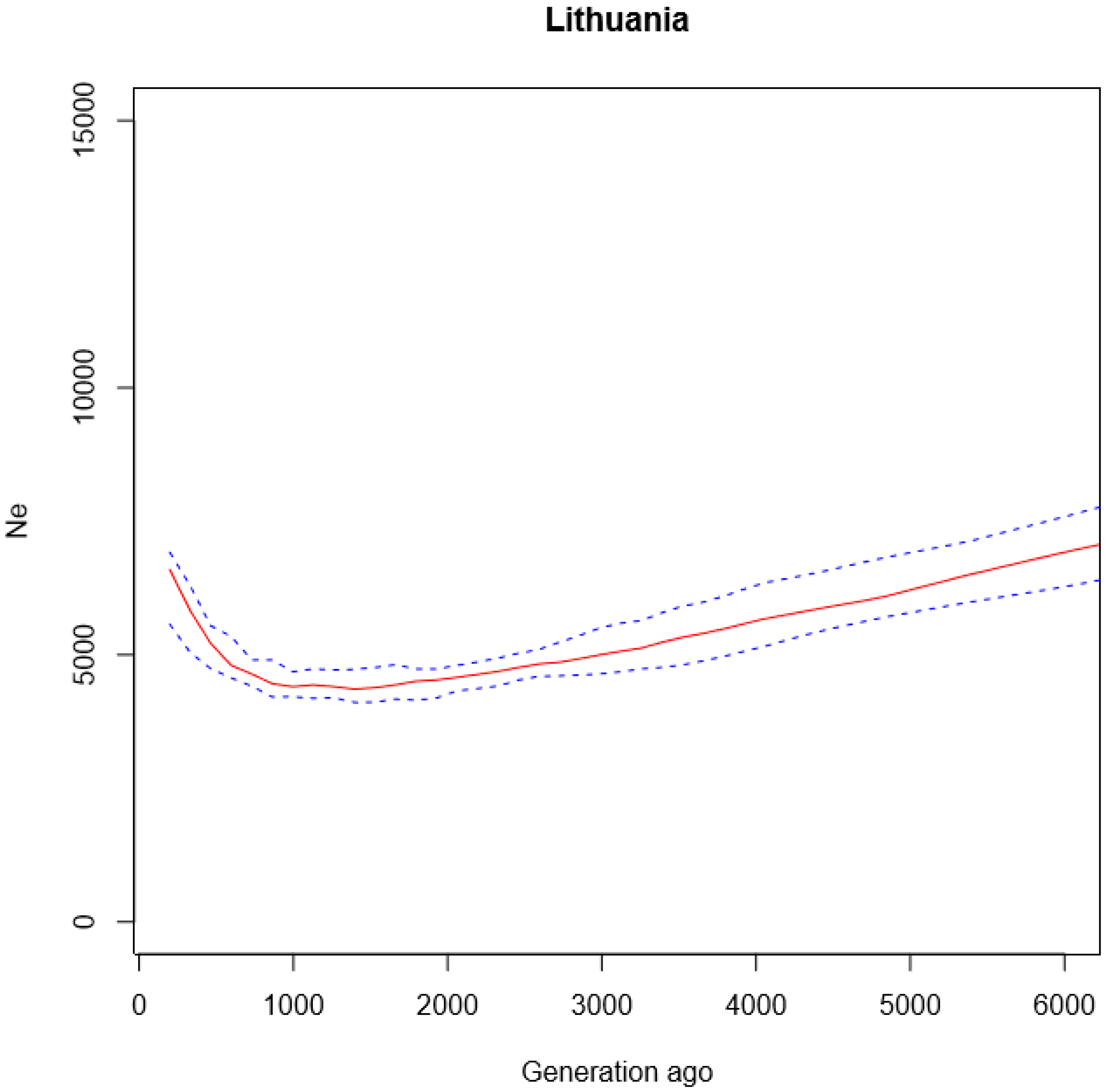
3.1. Historical Demography of Lithuania and Relationship to other Populations
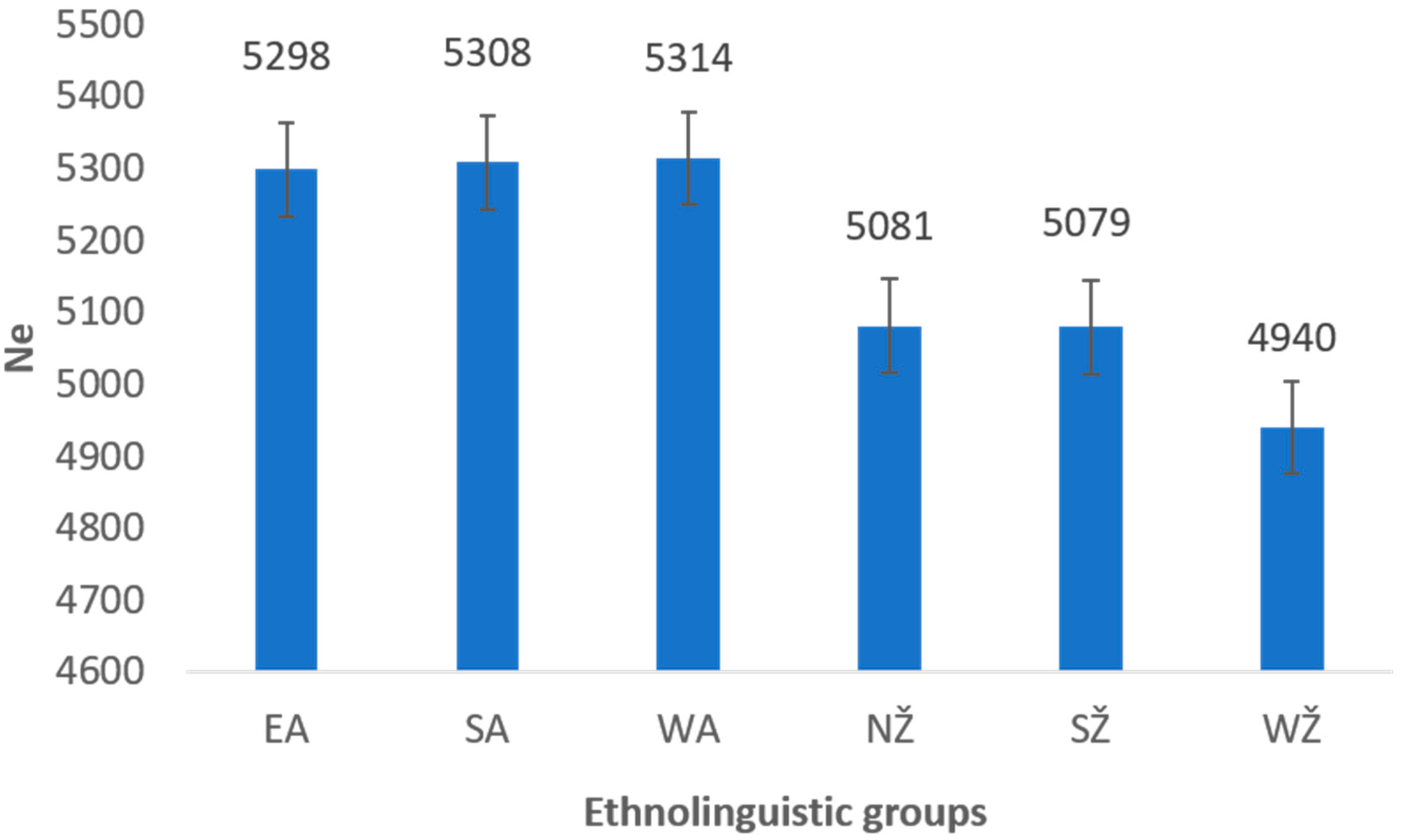
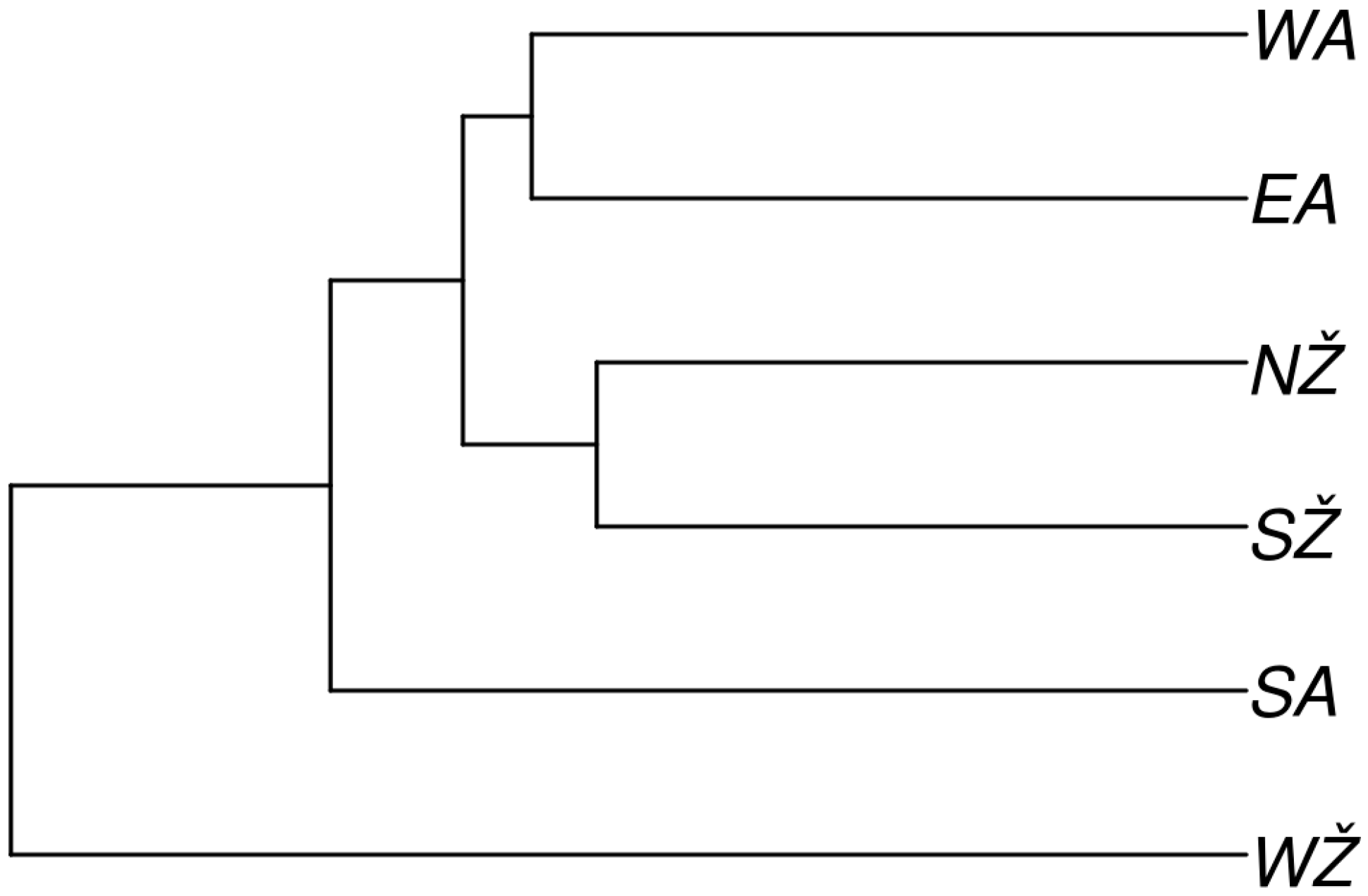
3.2. Reconstructing Past Events between Ethnolinguistic Regions of Lithuania
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wright, S. Evolution in Mendelian Populations. Genetics 1931, 16, 97–159. [Google Scholar] [PubMed]
- Frankham, R. Effective population size/adult population size ratios in wildlife: A review. Genet. Res. 2007, 89, 491–503. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, B. Fundamental concepts in genetics: Effective population size and patterns of molecular evolution and variation. Nat. Rev. Genet. 2009, 10, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Wang, J. Estimation of effective population sizes from data on genetic markers. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 1395–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voight, B.F.; Adams, A.M.; Frisse, L.A.; Qian, Y.; Hudson, R.R.; Di Rienzo, A. Interrogating multiple aspects of variation in a full resequencing data set to infer human population size changes. Proc. Natl. Acad. Sci. USA 2005, 102, 18508–18513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tassi, F.; Ghirotto, S.; Mezzavilla, M.; Vilaça, S.T.; De Santi, L.; Barbujani, G. Early modern human dispersal from Africa: Genomic evidence for multiple waves of migration. Investig. Genet. 2015, 6, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimas, A. Some unique futures of Lithuanian. Lituanus 1984, 30, 51–64. [Google Scholar]
- Rimantienė, R. Akmens Amžius Lietuvoje; Žiburys: Vilnius, Lithuania, 1966. [Google Scholar]
- Gimbutas, M. The Balts; Fredrick, A., Ed.; Praeger: London, UK; New York, NY, USA, 1963. [Google Scholar]
- Kučinskas, V. Population genetics of Lithuanians. Ann. Hum. Biol. 2001, 28, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Urnikyte, A.; Flores-Bello, A.; Mondal, M.; Molyte, A.; Comas, D.; Calafell, F.; Bosch, E.; Kučinskas, V. Patterns of genetic structure and adaptive positive selection in the Lithuanian population from high-density SNP data. Sci. Rep. 2019, 9, 9163. [Google Scholar] [CrossRef] [PubMed]
- Cann, H.M.; de Toma, C.; Cazes, L.; Legrand, M.F.; Morel, V.; Piouffre, L.; Bodmer, J.; Bodmer, W.F.; Bonne-Tamir, B.; Cambon-Thomsen, A.; et al. A human genome diversity cell line panel. Science 2002, 296, 261–262. [Google Scholar] [CrossRef] [PubMed]
- Urnikytė, A.; Molytė, A.; Kučinskas, V. Recent effective population size estimated from segments of identity by descent in the Lithuanian population. In Anthropological Science; The Anthropological Society of Nippon: Kyoto, Japan, 2017; Volume 125, pp. 53–58. [Google Scholar]
- Mezzavilla, M.; Ghirotto, S. Neon: An R Package to Estimate Human Effective Population Size and Divergence Time from Patterns of Linkage Disequilibrium between SNPS. J. Comput. Sci. Syst. Biol. 2015, 8, 37–44. [Google Scholar] [CrossRef]
- Benazzo, A.; Panziera, A.; Bertorelle, G. 4P: Fast computing of population genetics statistics from large DNA polymorphism panels. Ecol. Evol. 2015, 5, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Browning, S.R.; Browning, B.L. Accurate Non-parametric Estimation of Recent Effective Population Size from Segments of Identity by Descent. Am. J. Hum. Genet. 2015, 97, 404–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEvoy, B.P.; Powell, J.E.; Goddard, M.E.; Visscher, P.M. Human population dispersal “Out of Africa” estimated from linkage disequilibrium and allele frequencies of SNPs. Genome Res. 2011, 21, 821–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Q.; Posth, C.; Hajdinjak, M.; Petr, M.; Mallick, S.; Fernandes, D.; Furtwängler, A.; Haak, W.; Meyer, M.; Mittnik, A.; et al. The genetic history of Ice Age Europe. Nature 2016, 534, 200–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gronau, I.; Hubisz, M.J.; Gulko, B.; Danko, C.G.; Siepel, A. Bayesian inference of ancient human demography from individual genome sequences. Nat. Genet. 2011, 43, 1031–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellars, P. Why did modern human populations disperse from Africa ca. 60,000 years ago? A new model. PNAS 2006, 103, 9381–9386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gimbutiene, M. Baltai Priešistoriniais Laikais: Etnogenezė, Materialinė Kultūra ir Mitologija; Mokslas: Vilnius, Lithuania, 1985. [Google Scholar]
- Harvey, R.; Tills, D.; Warlow, A.; Kopec, C.; Domaniewska-Sobcz, K.; Suter, D.; Lord, M. Genetic affinities of the Balts: A study of blood groups, serum proteins and enzymes of Lithuanians in the United Kingdom. R. Anthropol. Inst. Great Br. Irel. 1983, 18, 535–552. [Google Scholar] [CrossRef]
- Geipel, J. The Europeans: An Ethnohistorical Survey, 1st ed.; Longmans: London, UK, 1969. [Google Scholar]
- Creanza, N.; Ruhlen, M.; Pemberton, T.J.; Rosenberg, N.A.; Feldman, M.W.; Ramachandran, S. A comparison of worldwide phonemic and genetic variation in human populations. Proc. Natl. Acad. Sci. USA 2015, 112, 1265–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urnikytė, A.; Molytė, A.; Pranckevičienė, E.; Kučinskienė, Z.A.; Kučinskas, V. Inferring Effective Population Size and Divergence Time in the Lithuanian Population According to High-Density Genotyping Data. Genes 2020, 11, 293. https://doi.org/10.3390/genes11030293
Urnikytė A, Molytė A, Pranckevičienė E, Kučinskienė ZA, Kučinskas V. Inferring Effective Population Size and Divergence Time in the Lithuanian Population According to High-Density Genotyping Data. Genes. 2020; 11(3):293. https://doi.org/10.3390/genes11030293
Chicago/Turabian StyleUrnikytė, Alina, Alma Molytė, Erinija Pranckevičienė, Zita Aušrelė Kučinskienė, and Vaidutis Kučinskas. 2020. "Inferring Effective Population Size and Divergence Time in the Lithuanian Population According to High-Density Genotyping Data" Genes 11, no. 3: 293. https://doi.org/10.3390/genes11030293
APA StyleUrnikytė, A., Molytė, A., Pranckevičienė, E., Kučinskienė, Z. A., & Kučinskas, V. (2020). Inferring Effective Population Size and Divergence Time in the Lithuanian Population According to High-Density Genotyping Data. Genes, 11(3), 293. https://doi.org/10.3390/genes11030293