Crosstalk between Dendritic Cells and Immune Modulatory Agents against Sepsis


Abstract
:1. Introduction
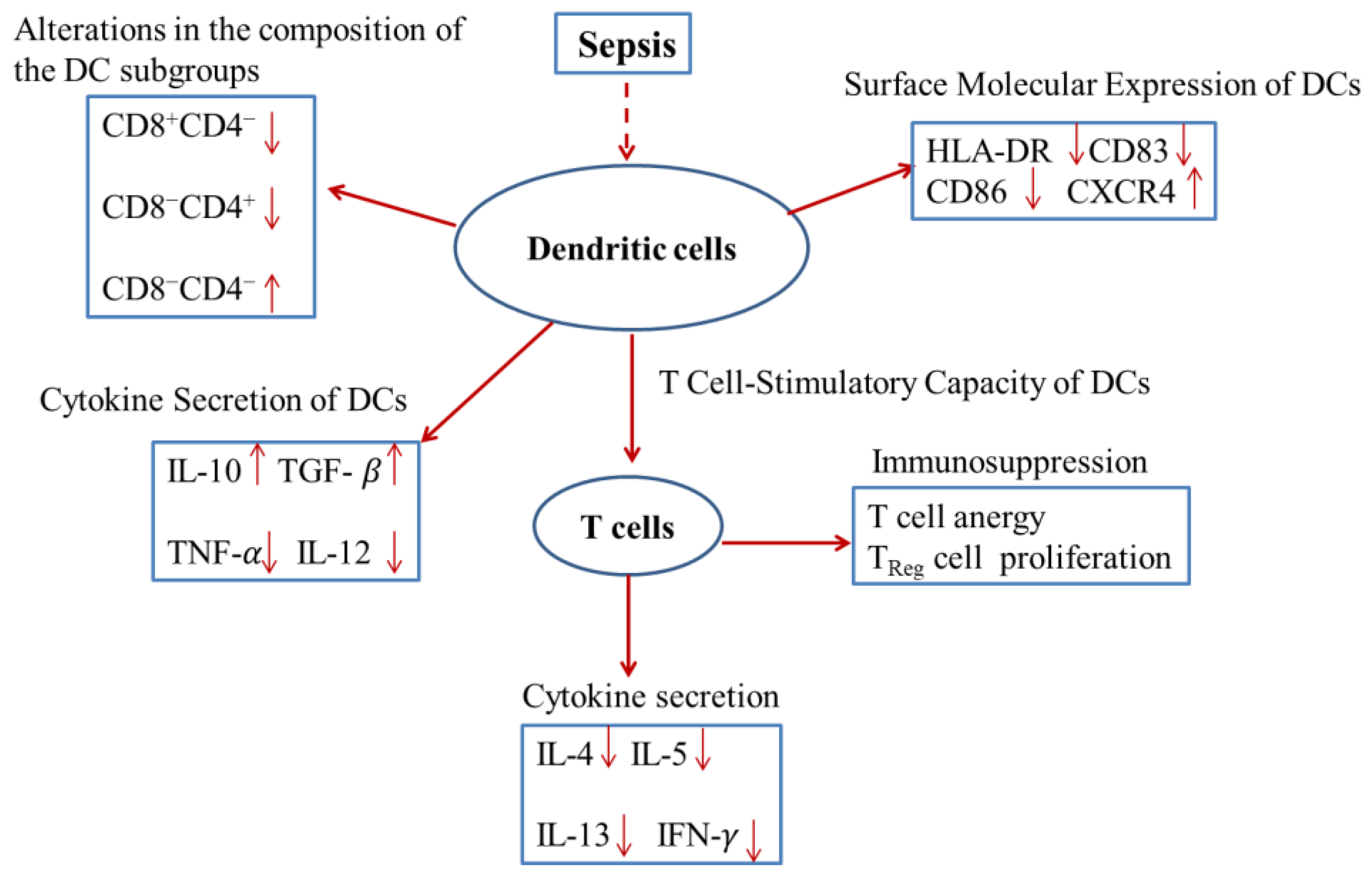
2. DC Numbers, Subgroups and Immune Functions in Sepsis
2.1. Sepsis can Lead to Reduction in DC Number
2.2. Alterations in the Composition of the DC Subgroups during Sepsis
2.3. Sepsis can Lead to Functional Impairments in DCs
3. Immunization of Sepsis with DC as the Target
3.1. Increasing the Number of DCs in Vivo
3.2. Anti-DC Apoptosis
3.3. Function Modification of DCs
4. New Approaches: Immunotherapies in Sepsis
4.1. Recombinant Human IL-7
4.2. PD1/ PDL1-Specific Antibodies
4.3. IFN- γ
4.4. G-CSF and GM-CSF
4.5. IL-15
4.6. IL-1ra
4.7. IL-6
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cavaillon, J.M.; Singer, M.; Skirecki, T. Sepsis therapies: Learning from 30 years of failure of translational research to propose new leads. EMBO Mol. Med. 2020, e10128. [Google Scholar] [CrossRef] [PubMed]
- Strother, R.K.; Danahy, D.B.; Kotov, D.I.; Kucaba, T.A.; Zacharias, Z.R.; Griffith, T.S.; Legge, K.L.; Badovinac, V.P. Polymicrobial Sepsis Diminishes Dendritic Cell Numbers and Function Directly Contributing to Impaired Primary CD8 T-Cell Responses in vivo. J. Immunol. 2016, 197, 4301–4311. [Google Scholar] [CrossRef] [PubMed]
- Weber, G.F.; Maier, S.L.; Tina, Z.N.; Michael, B.; Therese, S.; Isabella, K.; Jürgen, W. Analysis of circulating plasmacytoid dendritic cells during the course of sepsis. Surgery 2015, 158, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Gnoni, A.; De Nitto, E.; Scacco, S.; Santacroce, L.; Palese, L.L. A New Look at the Structures of Old Sepsis Actors by Exploratory Data Analysis Tools. Antibiotics 2019, 8, 225. [Google Scholar] [CrossRef] [Green Version]
- Rubio, I.; Osuchowski, M.F.; Shankar-Hari, M.; Skirecki, T.; Winkler, M.S.; Lachmann, G.; La Rosée, P.; Monneret, G.; Venet, F.; Bauer, M.; et al. Current gaps in sepsis immunology: New opportunities for translational research. Lancet Infect. Dis. 2019, 19, e422–e436. [Google Scholar] [CrossRef]
- Darkwah, S.; Nago, N.; Appiah, M.G.; Myint, P.K.; Kawamoto, E.; Shimaoka, M.; Park, E.J. Differential Roles of Dendritic Cells in Expanding CD4 T Cells in Sepsis. Biomedicines 2019, 7, 52. [Google Scholar] [CrossRef] [Green Version]
- Elsayh, K.I.; Zahran, A.M.; Mohamad, I.L.; Aly, S.S. Dendritic cells in childhood sepsis. J. Crit. Care 2013, 28, 881.e7–881.e13. [Google Scholar] [CrossRef]
- Schraml, B.U.; Reis e Sousa, C. Defining dendritic cells. Curr. Opin. Immunol. 2015, 32, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Collin, M.; Mcgovern, N.; Haniffa, M. Human dendritic cell subsets. Immunology 2013, 140, 22–30. [Google Scholar] [CrossRef]
- Vincent, J.L.; Mongkolpun, W. Non-antibiotic therapies for sepsis: An update. Expert Rev. Anti-Infe. 2019, 17, 169–175. [Google Scholar] [CrossRef]
- Venet, F.; Monneret, G. Advances in the understanding and treatment of sepsis-induced immunosuppression. Nat. Rev. Nephro. 2018, 14, 121–137. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhao, R.; Fan, K.; Iwanowycz, S.; Fan, H.; Li, Z.; Liu, B. Regulation of dendritic cell function improves survival in experimental sepsis through immune chaperone. Innate. Immun. 2019, 25, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Bouras, M.; Asehnoune, K.; Roquilly, A. Contribution of Dendritic Cell Responses to Sepsis-Induced Immunosuppression and to Susceptibility to Secondary Pneumonia. Front. Immunol. 2018, 9, 2590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guisset, O.; Dilhuydy, M.S.; Thiebaut, R.; Lefevre, J.; Camou, F.; Sarrat, A.; Gabinski, C.; Moreau, J.F.; Blanco, P. Decrease in circulating dendritic cells predicts fatal outcome in septic shock. Intensive Care Med. 2007, 33, 148–152. [Google Scholar] [CrossRef]
- Kumar, V. Dendritic cells in sepsis: Potential immunoregulatory cells with therapeutic potential. Mol. Immunol. 2018, 101, 615–626. [Google Scholar] [CrossRef]
- Macri, C.; Pang, E.S.; Patton, T.; O’Keeffe, M. Dendritic cell subsets. Semin. Cell Dev. Biol. 2018, 84, 11–21. [Google Scholar] [CrossRef]
- Sato, K.; Fujita, S. Dendritic Cells-Nature and Classification. Allergol. Int. 2007, 56, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Poehlmann, H.; Schefold, J.C.; Zuckermann-Becker, H.; Volk, H.-D.; Meisel, C. Phenotype changes and impaired function of dendritic cell subsets in patients with sepsis: A prospective observational analysis. Crit. Care 2009, 13, R119. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Venet, F.; Chung, C.S.; Lomas-Neira, J.; Ayala, A. Changes in dendritic cell function in the immune response to sepsis. Cell- & tissue-based therapy. Expert Opin. Biol. Ther. 2007, 7, 929–938. [Google Scholar]
- Nie, F.; Ding, F.; Chen, B.; Huang, S.; Liu, Q.; Xu, C. Dendritic cells aggregate inflammation in experimental osteoarthritis through a toll-like receptor (TLR)-dependent machinery response to challenges. Life Sci. 2019, 238, 116920. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhu, X.; Feng, Y.; Pang, W.; Qi, Z.; Cui, L.; Cao, Y. TLR4 and TLR9 signals stimulate protective immunity against blood-stage Plasmodium yoelii infection in mice. Exp. Parasitol. 2016, 170, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Flohe, S.B. Dendritic cells during polymicrobial sepsis rapidly mature but fail to initiate a protective Th1-type immune response. J. Leukoc. Biol. 2005, 79, 473–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appel, S.; Faivre, V.; Lukaszewicz, A.C.; Alves, A.; Charron, D.; Payen, D.; Haziot, A. Human Monocytes Differentiate into Dendritic Cells Subsets that Induce Anergic and Regulatory T Cells in Sepsis. PLoS ONE 2012, 7, e47209. [Google Scholar]
- Wang, H.-W.; Yang, W.; Gao, L.; Kang, J.-R.; Qin, J.-J.; Liu, Y.-P.; Lu, J.-Y. Adoptive transfer of bone marrow-derived dendritic cells decreases inhibitory and regulatory T-cell differentiation and improves survival in murine polymicrobial sepsis. Immunology 2015, 145, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Faivre, V.; Lukaszewicz, A.C.; Alves, A.; Charron, D.; Payen, D.; Haziot, A. Accelerated in vitro differentiation of blood monocytes into dendritic cells in human sepsis. Clin. Exp. Immunol. 2007, 147, 426–439. [Google Scholar] [CrossRef]
- Fan, X.; Liu, Z. Alterations of dendritic cells in sepsis: Featured role in immunoparalysis. 2015, 2015, 903720. [Google Scholar] [CrossRef]
- Shubin, N.J.; Chung, C.S.; Heffernan, D.S.; Irwin, L.R.; Monaghan, S.F.; Ayala, A. BTLA expression contributes to septic morbidity and mortality by inducing innate inflammatory cell dysfunction. J. Leukoc. Biol. 2012, 92, 593–603. [Google Scholar] [CrossRef] [Green Version]
- Cheng, T.; Bai, J.; Chung, C.-S.; Chen, Y.; Biron, B.M.; Ayala, A. Enhanced Innate Inflammation Induced by Anti-BTLA Antibody in Dual Insult Model of Hemorrhagic Shock/Sepsis. Shock 2016, 45, 40–49. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, Y.; Iwata, A.; Suzuki, K.; Suto, A.; Kawashima, S.; Saito, Y.; Owada, T.; Kobayashi, M.; Watanabe, N.; Nakajima, H. B and T lymphocyte attenuator inhibits LPS-induced endotoxic shock by suppressing Toll-like receptor 4 signaling in innate immune cells. Proc. Natl. Acad. Sci. USA 2013, 110, 5121–5126. [Google Scholar] [CrossRef] [Green Version]
- Wodnar-Filipowicz, A. Flt3 Ligand: Role in Control of Hematopoietic and Immune Functions of the Bone Marrow. Physiology 2003, 18, 247–251. [Google Scholar] [CrossRef]
- Li, J.; Chen, S.; Ge, J.; Lu, F.; Ren, S.; Zhao, Z.; Pu, X.; Chen, X.; Sun, J.; Gu, Y. A novel therapeutic vaccine composed of a rearranged human papillomavirus type 16 E6/E7 fusion protein and Fms-like tyrosine kinase-3 ligand induces CD8 + T cell responses and antitumor effect. Vaccine 2017, 35, 6459–6467. [Google Scholar] [CrossRef] [PubMed]
- Ramos, M.I.; Tak, P.P.; Lebre, M.C. Fms-like tyrosine kinase 3 ligand-dependent dendritic cells in autoimmune inflammation. Autoimmun. Rev. 2014, 13, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Toliver-Kinsky, T.E.; Cui, W.; Murphey, E.D.; Lin, C.; Sherwood, E.R. Enhancement of Dendritic Cell Production by Fms-Like Tyrosine Kinase-3 Ligand Increases the Resistance of Mice to a Burn Wound Infection. J. Immunol. 2004, 174, 404–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, L.; Bohannon, J.K.; Cui, W.; Vinish, M.; Toliver-Kinsky, T. Fms-like tyrosine kinase-3 ligand increases resistance to burn wound infection through effects on plasmacytoid dendritic cells. BMC Immunol. 2017, 18, 9. [Google Scholar] [CrossRef] [Green Version]
- Patil, N.K.; Bohannon, J.K.; Luan, L.; Guo, Y.; Fensterheim, B.; Hernandez, A.; Wang, J.; Sherwood, E.R. Flt3 Ligand Treatment Attenuates T Cell Dysfunction and Improves Survival in a Murine Model of Burn Wound Sepsis. Shock 2017, 47, 40–51. [Google Scholar] [CrossRef]
- Hundeshagen, G.; Cui, W.; Musgrove, L.; Cherry, A.; Lee, S.-J.; Cox, R.A.; Toliver-Kinsky, T. Fms-Like Tyrosine Kinase-3 Ligand Attenuates Local and Systemic Infection in a Model of Post-Burn Pneumonia. Shock 2017, 1, 721–727. [Google Scholar] [CrossRef]
- Grailer, J.J.; Fattahi, F.; Dick, R.S.; Zetoune, F.S.; Ward, P.A. Cutting Edge: Critical Role for C5aRs in the Development of Septic Lymphopenia in Mice. J. Immunol. 2015, 194, 868–872. [Google Scholar] [CrossRef] [Green Version]
- Aballay, A.; Flierl, M.A.; Rittirsch, D.; Chen, A.J.; Nadeau, B.A.; Day, D.E.; Sarma, J.V.; Huber-Lang, M.S.; Ward, P.A. The Complement Anaphylatoxin C5a Induces Apoptosis in Adrenomedullary Cells during Experimental Sepsis. PLoS ONE 2008, 3, e2560. [Google Scholar]
- Rittirsch, D.; Flierl, M.A.; Nadeau, B.A.; Day, D.E.; Huber-Lang, M.; Mackay, C.R.; Zetoune, F.S.; Gerard, N.P.; Cianflone, K.; Köhl, J.; et al. Functional roles for C5a receptors in sepsis. Nat. Med. 2008, 14, 551–557. [Google Scholar] [CrossRef]
- Ma, N.; Xing, C.; Xiao, H.; Wang, Y.; Wang, K.; Hou, C.; Han, G.; Chen, G.; Marrero, B.; Wang, Y.; et al. C5a regulates IL-12+ DC migration to induce pathogenic Th1 and Th17 cells in sepsis. PLoS ONE 2013, 8, e69779. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Wang, J. Programmed cell death of dendritic cells in immune regulation. Immunol. Rev. 2010, 236, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Bhan, C.; Dipankar, P.; Chakraborty, P.; Sarangi, P.P. Role of cellular events in the pathophysiology of sepsis. Inflamm. Res. 2016, 65, 853–868. [Google Scholar] [CrossRef] [PubMed]
- Iwata, A.; Stevenson, V.M.; Minard, A.; Tasch, M.; Tupper, J.; Lagasse, E.; Weissman, I.; Harlan, J.M.; Winn, R.K. Over-Expression of Bcl-2 Provides Protection in Septic Mice by a trans Effect. J. Immunol. 2003, 171, 3136–3141. [Google Scholar] [CrossRef] [PubMed]
- Gautier, E.L.; Huby, T.; Saint-Charles, F.; Ouzilleau, B.; Chapman, M.J.; Lesnik, P. Enhanced Dendritic Cell Survival Attenuates Lipopolysaccharide-Induced Immunosuppression and Increases Resistance to Lethal Endotoxic Shock. J. Immunol. 2008, 180, 6941–6946. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Huang, X.; Lu, C.; Cairo, M.S.; Zhou, X. MicroRNA-146a and MicroRNA-146b Regulate Human Dendritic Cell Apoptosis and Cytokine Production by Targeting TRAF6 and IRAK1 Proteins. J. Biol. Chem. 2015, 290, 2831–2841. [Google Scholar] [CrossRef] [Green Version]
- Koppi, T.A.; Tough-Bement, T.; Lewinsohn, D.M.; Lynch, D.H.; Alderson, M.R. CD40 ligand inhibits Fas/CD95-mediated apoptosis of human blood-derived dendritic cells. Eur. J. Immunol. 1997, 27, 3161–3165. [Google Scholar] [CrossRef]
- Michels, M.; Danieslki, L.G.; Vieira, A.; Florentino, D.; Dall’Igna, D.; Galant, L.; Sonai, B.; Vuolo, F.; Mina, F.; Pescador, B.; et al. CD40-CD40 Ligand Pathway is a Major Component of Acute Neuroinflammation and Contributes to Long-term Cognitive Dysfunction after Sepsis. Mol. Med. (Camb. Mass.) 2015, 21, 219–226. [Google Scholar] [CrossRef]
- Sinistro, A.; Almerighi, C.; Ciaprini, C.; Natoli, S.; Sussarello, E.; Di Fino, S.; Calo-Carducci, F.; Rocchi, G.; Bergamini, A. Downregulation of CD40 Ligand Response in Monocytes from Sepsis Patients. Clin. Vaccine Immunol. 2008, 15, 1851–1858. [Google Scholar] [CrossRef] [Green Version]
- Wong, B.R.; Josien, R.; Lee, S.Y.; Sauter, B.; Li, H.L.; Steinman, R.M.; Choi, Y. TRANCE (tumor necrosis factor [TNF]-related activation-induced cytokine), a new TNF family member predominantly expressed in T cells, is a dendritic cell-specific survival factor. J. Exp. Med. 1997, 186, 2075–2080. [Google Scholar] [CrossRef]
- Cremer, I. Long-lived immature dendritic cells mediated by TRANCE-RANK interaction. Blood 2002, 100, 3646–3655. [Google Scholar] [CrossRef] [Green Version]
- Alcain, J.; Podaza, E.; Gori, M.S.; Salamone, G.; Vermeulen, M. Modulation of Dendritic Cell Apoptosis and CD8+ Cytotoxicity by Histamine: Role of Protein Kinase C. Mediat. Inflamm. 2017, 2017, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.-T.; Yao, Y.-M.; Yao, F.-H.; Huang, L.-F.; Dong, N.; Yu, Y.; Sheng, Z.-Y. Association between High-Mobility Group Box-1 Protein Release and Immune Function of Dendritic Cells in Thermal Injury. J. Interferon Cytokine Res. 2010, 30, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.-S. Role of high mobility group box 1 in inflammatory disease: Focus on sepsis. Arch. Pharmacal Res. 2012, 35, 1511–1523. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Choi, J.G.; Abraham, S.; Wu, H.; Diaz, D.; Terreros, D.; Shankar, P.; Manjunath, N. Human macrophage and dendritic cell-specific silencing of high-mobility group protein B1 ameliorates sepsis in a humanized mouse model. Proc. Natl. Acad. Sci. USA 2012, 109, 21052–21057. [Google Scholar] [CrossRef] [Green Version]
- Meng, Y.; Zhao, Z.; Zhu, W.; Yang, T.; Deng, X.; Bao, R. CD155 blockade improves survival in experimental sepsis by reversing dendritic cell dysfunction. Biochem. Biophys. Res. Commun. 2017, 490, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Pene, F.; Courtine, E.; Ouaaz, F.; Zuber, B.; Sauneuf, B.; Sirgo, G.; Rousseau, C.; Toubiana, J.; Balloy, V.; Chignard, M.; et al. Toll-Like Receptors 2 and 4 Contribute to Sepsis-Induced Depletion of Spleen Dendritic Cells. Infect. Immun. 2009, 77, 5651–5658. [Google Scholar] [CrossRef] [Green Version]
- Perrin-Cocon, L.; Aublin-Gex, A.; Sestito, S.E.; Shirey, K.A.; Patel, M.C.; Andre, P.; Blanco, J.C.; Vogel, S.N.; Peri, F.; Lotteau, V. TLR4 antagonist FP7 inhibits LPS-induced cytokine production and glycolytic reprogramming in dendritic cells, and protects mice from lethal influenza infection. Sci. Rep. 2017, 7, 40791. [Google Scholar] [CrossRef]
- Deng, M.; Ma, T.; Yan, Z.; Zettel, K.R.; Scott, M.J.; Liao, H.; Frank, A.; Morelli, A.E.; Sodhi, C.P.; Hackam, D.J.; et al. Toll-like Receptor 4 Signaling on Dendritic Cells Suppresses Polymorphonuclear Leukocyte CXCR2 Expression and Trafficking via Interleukin 10 During Intra-abdominal Sepsis. J. Infect. Dis. 2016, 213, 1280–1288. [Google Scholar] [CrossRef] [Green Version]
- Barrenschee, M.; Lex, D.; Uhlig, S. Effects of the TLR2 agonists MALP-2 and Pam3Cys in isolated mouse lungs. PLoS ONE 2010, 5, e13889. [Google Scholar] [CrossRef] [Green Version]
- Raby, A.C.; Holst, B.; Le Bouder, E.; Diaz, C.; Ferran, E.; Conraux, L.; Guillemot, J.C.; Coles, B.; Kift-Morgan, A.; Colmont, C.S.; et al. Targeting the TLR Co-Receptor CD14 with TLR2-Derived Peptides Modulates Immune Responses to Pathogens. Sci. Transl. Med. 2013, 5, ra64–ra185. [Google Scholar] [CrossRef]
- Baumann, C.L.; Aspalter, I.M.; Sharif, O.; Pichlmair, A.; Blüml, S.; Grebien, F.; Bruckner, M.; Pasierbek, P.; Aumayr, K.; Planyavsky, M.; et al. CD14 is a coreceptor of Toll-like receptors 7 and 9. J. Exp. Med. 2010, 207, 2689–2701. [Google Scholar] [CrossRef]
- Atalan, N.; Acar, L.; Yapici, N.; Kudsioglu, T.; Ergen, A.; Yilmaz, S.G.; Isbir, T. The Relationship Between Sepsis-induced Immunosuppression and Serum Toll-like Receptor 9 Level. Vivo 2018, 32, 1653–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eugenin, E.A.; Tan, T.L.; Goh, Y.Y. The role of group IIA secretory phospholipase A2 (sPLA2-IIA) as a biomarker for the diagnosis of sepsis and bacterial infection in adults—A systematic review. PLoS ONE 2017, 12, e0180554. [Google Scholar]
- Perrin-Cocon, L.; Agaugué, S.; Coutant, F.; Masurel, A.; Bezzine, S.; Lambeau, G.; André, P.; Lotteau, V. Secretory phospholipase A2 induces dendritic cell maturation. Eur. J. Immunol. 2004, 34, 2293–2302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Varambally, S.; Maher, C.A.; Cao, Q.; Chockley, P.; Toubai, T.; Malter, C.; Nieves, E.; Tawara, I.; Wang, Y.; et al. Targeting of microRNA-142-3p in dendritic cells regulates endotoxin-induced mortality. Blood 2011, 117, 6172–6183. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Potter, C.S.; Sundberg, J.P.; Hogenesch, H. SHARPIN is a key regulator of immune and inflammatory responses. J. Cell. Mol. Med. 2012, 16, 2271–2279. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, S.K.; Wang, Z.; Sokolovska, A.; Seymour, R.; Sundberg, J.P.; HogenEsch, H. SHARPIN Is Essential for Cytokine Production, NF-κB Signaling, and Induction of Th1 Differentiation by Dendritic Cells. PLoS ONE 2012, 7, e31809. [Google Scholar]
- Nastase, M.-V.; Zeng-Brouwers, J.; Frey, H.; Hsieh, L.T.-H.; Poluzzi, C.; Beckmann, J.; Schroeder, N.; Pfeilschifter, J.; Lopez-Mosqueda, J.; Mersmann, J.; et al. An Essential Role for SHARPIN in the Regulation of Caspase 1 Activity in Sepsis. Am. J. Pathol. 2016, 186, 1206–1220. [Google Scholar] [CrossRef] [Green Version]
- Marrack, P.; Li, C.C.; Munitic, I.; Mittelstadt, P.R.; Castro, E.; Ashwell, J.D. Suppression of Dendritic Cell-Derived IL-12 by Endogenous Glucocorticoids Is Protective in LPS-Induced Sepsis. PLoS Biol. 2015, 13, e1002269. [Google Scholar]
- Robinson, R. Glucocorticoids Reduce Sepsis by Diminishing Dendritic Cell Responses. PLoS Biol. 2015, 13, e1002270. [Google Scholar] [CrossRef]
- Bohannon, J.; Cui, W.; Cox, R.; Przkora, R.; Sherwood, E.; Toliver-Kinsky, T. Prophylactic Treatment with Fms-Like Tyrosine Kinase-3 Ligand after Burn Injury Enhances Global Immune Responses to Infection. J. Immunol. 2008, 180, 3038–3048. [Google Scholar] [CrossRef] [Green Version]
- Beshara, R.; Sencio, V. Alteration of Flt3-Ligand-dependent de novo generation of conventional dendritic cells during influenza infection contributes to respiratory bacterial superinfection. PLoS pathog. 2018, 14, e1007360. [Google Scholar] [CrossRef]
- Peck-Palmer, O.M.; Unsinger, J.; Chang, K.C.; McDonough, J.S.; Perlman, H.; McDunn, J.E.; Hotchkiss, R.S. Modulation of the Bcl-2 Family Blocks Sepsis-Induced Depletion of Dendritic Cells and Macrophages. Shock (AugustaGa.) 2009, 31, 359–366. [Google Scholar] [CrossRef] [Green Version]
- Raby, A.C.; Labeta, M.O. Therapeutic Boosting of the Immune Response: Turning to CD14 for Help. Curr. Pharm. Biotechnol. 2016, 17, 414–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramoner, R. Dendritic-cell activation by secretory phospholipase A2. Blood 2005, 105, 3583–3587. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Gharbi, A.; You, J.S.; Han, H.D.; Kang, T.H.; Hong, S.H.; Park, W.S.; Jung, I.D.; Park, Y.M. Drug repositioning of TANK-binding kinase 1 inhibitor CYT387 as an alternative for the treatment of Gram-negative bacterial sepsis. Int. Immunopharmacol. 2019, 73, 482–490. [Google Scholar] [CrossRef] [PubMed]
- Peters van Ton, A.M.; Kox, M.; Abdo, W.F.; Pickkers, P. Precision Immunotherapy for Sepsis. Front. Immunol. 2018, 9, 1926. [Google Scholar] [CrossRef]
- Watanabe, E.; Thampy, L.K.; Hotchkiss, R.S. Immunoadjuvant therapy in sepsis: Novel strategies for immunosuppressive sepsis coming down the pike. Acute Med. Surg. 2018, 5, 309–315. [Google Scholar] [CrossRef]
- Oh, B.M.; Lee, S.J.; Park, G.L.; Hwang, Y.S.; Lim, J.; Park, E.S.; Lee, K.H.; Kim, B.Y.; Kwon, Y.T.; Cho, H.J.; et al. Erastin Inhibits Septic Shock and Inflammatory Gene Expression via Suppression of the NF-kappaB Pathway. J. Clin. Med. 2019, 8, 2210. [Google Scholar] [CrossRef] [Green Version]
- Santacroce, L.; Charitos, I.A.; Bottalico, L. A successful history: Probiotics and their potential as antimicrobials. Expert Rev. Anti-Infect. Ther. 2019, 17, 635–645. [Google Scholar] [CrossRef]
- Alkhateeb, T.; Kumbhare, A.; Bah, I.; Youssef, D.; Yao, Z.Q.; McCall, C.E.; El Gazzar, M. S100A9 maintains myeloid-derived suppressor cells in chronic sepsis by inducing miR-21 and miR-181b. Mol. Immunol. 2019, 112, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Seymour, C.W.; Rosengart, M.R. Septic Shock: Advances in Diagnosis and Treatment. JAMA 2015, 314, 708–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peake, S.L.; Delaney, A.; Bailey, M.; Bellomo, R.; Cameron, P.A.; Cooper, D.J.; Higgins, A.M.; Holdgate, A.; Howe, B.D.; Webb, S.A.; et al. Goal-directed resuscitation for patients with early septic shock. N. Engl. J. Med. 2014, 371, 1496–1506. [Google Scholar] [PubMed] [Green Version]
- Leentjens, J.; Kox, M.; van der Hoeven, J.G.; Netea, M.G.; Pickkers, P. Immunotherapy for the Adjunctive Treatment of Sepsis: From Immunosuppression to Immunostimulation. Time for a Paradigm Change? Am. J. Respir. Crit. Care Med. 2013, 187, 1287–1293. [Google Scholar] [CrossRef]
- Schrijver, I.T.; Theroude, C.; Roger, T. Myeloid-Derived Suppressor Cells in Sepsis. Front. Immunol. 2019, 10, 327. [Google Scholar] [CrossRef] [Green Version]
- Venet, F.; Demaret, J.; Blaise, B.J.; Rouget, C.; Girardot, T.; Idealisoa, E.; Rimmelé, T.; Mallet, F.; Lepape, A.; Textoris, J.; et al. IL-7 Restores T Lymphocyte Immunometabolic Failure in Septic Shock Patients through mTOR Activation. J. Immunol. 2017, 199, 1606–1615. [Google Scholar] [CrossRef]
- Mackall, C.L.; Fry, T.J.; Gress, R.E. Harnessing the biology of IL-7 for therapeutic application. Nat. Rev. Immunol. 2011, 11, 330–342. [Google Scholar] [CrossRef]
- Hotchkiss, R.S.; Sherwood, E.R. Immunology. Getting sepsis therapy right. Science 2015, 347, 1201–1202. [Google Scholar] [CrossRef] [Green Version]
- Unsinger, J.; McGlynn, M.; Kasten, K.R.; Hoekzema, A.S.; Watanabe, E.; Muenzer, J.T.; McDonough, J.S.; Tschoep, J.; Ferguson, T.A.; McDunn, J.E.; et al. IL-7 Promotes T Cell Viability, Trafficking, and Functionality and Improves Survival in Sepsis. J. Immunol. 2010, 184, 3768–3779. [Google Scholar] [CrossRef] [Green Version]
- Shindo, Y.; Unsinger, J.; Burnham, C.-A.; Green, J.M.; Hotchkiss, R.S. Interleukin-7 and Anti–Programmed Cell Death 1 Antibody Have Differing Effects to Reverse Sepsis-Induced Immunosuppression. Shock (AugustaGa.) 2015, 43, 334–343. [Google Scholar] [CrossRef] [Green Version]
- Ariyasu, R.; Horiike, A.; Yoshizawa, T.; Dotsu, Y.; Koyama, J.; Saiki, M.; Sonoda, T.; Nishikawa, S.; Kitazono, S.; Yanagitani, N.; et al. Adrenal Insufficiency Related to Anti-Programmed Death-1 Therapy. Anticancer Res. 2017, 37, 4229–4232. [Google Scholar] [PubMed]
- Monneret, G.; Gossez, M.; Venet, F. Sepsis in PD-1 light. Crit. Care 2016, 20, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhou, Y.; Lou, J.; Li, J.; Bo, L.; Zhu, K.; Wan, X.; Deng, X.; Cai, Z. PD-L1 blockade improves survival in experimental sepsis by inhibiting lymphocyte apoptosis and reversing monocyte dysfunction. Crit. Care 2010, 14, R220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shindo, Y.; McDonough, J.S.; Chang, K.C.; Ramachandra, M.; Sasikumar, P.G.; Hotchkiss, R.S. Anti-PD-L1 peptide improves survival in sepsis. J. Surg. Res. 2017, 208, 33–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, K.C.; Burnham, C.A.; Compton, S.M.; Rasche, D.P.; Mazuski, R.J.; McDonough, J.S.; Unsinger, J.; Korman, A.J.; Green, J.M.; Hotchkiss, R.S. Blockade of the negative co-stimulatory molecules PD-1 and CTLA-4 improves survival in primary and secondary fungal sepsis. Crit. Care (Lond. Engl.) 2013, 17, R85. [Google Scholar] [CrossRef] [Green Version]
- Brahmamdam, P.; Inoue, S.; Unsinger, J.; Chang, K.C.; McDunn, J.E.; Hotchkiss, R.S. Delayed administration of anti-PD-1 antibody reverses immune dysfunction and improves survival during sepsis. J. Leukoc. Biol. 2010, 88, 233–240. [Google Scholar] [CrossRef]
- Patera, A.C.; Drewry, A.M.; Chang, K.; Beiter, E.R.; Osborne, D.; Hotchkiss, R.S. Frontline Science: Defects in immune function in patients with sepsis are associated with PD-1 or PD-L1 expression and can be restored by antibodies targeting PD-1 or PD-L1. J. Leukoc. Biol. 2016, 100, 1239–1254. [Google Scholar] [CrossRef]
- Jarvis, J.N.; Meintjes, G.; Rebe, K.; Williams, G.N.; Bicanic, T.; Williams, A.; Schutz, C.; Bekker, L.-G.; Wood, R.; Harrison, T.S. Adjunctive interferon-γ immunotherapy for the treatment of HIV-associated cryptococcal meningitis. Aids 2012, 26, 1105–1113. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Oh, S.J.; Ahn, S.; Chung, D.H. IFN-γ-producing NKT cells exacerbate sepsis by enhancing C5a generation via IL-10-mediated inhibition of CD55 expression on neutrophils. Eur. J. Immunol. 2014, 44, 2025–2035. [Google Scholar] [CrossRef]
- Nalos, M.; Santner-Nanan, B.; Parnell, G.; Tang, B.; McLean, A.S.; Nanan, R. Immune effects of interferon γ in persistent staphylococcal sepsis. Am. J. Respir. Crit. Care Med. 2012, 185, 110–112. [Google Scholar] [CrossRef]
- Patil, N.K.; Bohannon, J.K.; Sherwood, E.R. Immunotherapy: A promising approach to reverse sepsis-induced immunosuppression. Pharmacol. Res. 2016, 111, 688–702. [Google Scholar] [CrossRef] [PubMed]
- Delano, M.J.; Ward, P.A. Sepsis-induced immune dysfunction: Can immune therapies reduce mortality? J. Clin. Investig. 2016, 126, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Priyanka Gupta, R.S. Om Shankar Chaurasia, Anuj Sethi, Role of Granulocyte Colony Stimulating Factor (G-Csf) in Neonatal Sepsis with Neutropenia. Peoples J. Sci. Res. 2016, 9, 7–13. [Google Scholar]
- Cheng, A.C.; Limmathurotsakul, D.; Chierakul, W.; Getchalarat, N.; Wuthiekanun, V.; Stephens, D.P.; Day, N.P.J.; White, N.J.; Chaowagul, W.; Currie, B.J.; et al. A Randomized Controlled Trial of Granulocyte Colony-Stimulating Factor for the Treatment of Severe Sepsis Due to Melioidosis in Thailand. Clin. Infect. Dis. 2007, 45, 308–314. [Google Scholar] [CrossRef]
- Becher, B.; Tugues, S.; Greter, M. GM-CSF: From Growth Factor to Central Mediator of Tissue Inflammation. Immunity 2016, 45, 963–973. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, P.; Budnick, I.; Singh, M.; Thiruppathi, M.; Alharshawi, K.; Elshabrawy, H.; Holterman, M.J.; Prabhakar, B.S. Dual Role of GM-CSF as a Pro-Inflammatory and a Regulatory Cytokine: Implications for Immune Therapy. J. Interferon Cytokine Res. 2015, 35, 585–599. [Google Scholar] [CrossRef] [Green Version]
- Marlow, N.; Morris, T.; Brocklehurst, P.; Carr, R.; Cowan, F.; Patel, N.; Petrou, S.; Redshaw, M.; Modi, N.; Doré, C.J. A randomised trial of granulocyte-macrophage colony-stimulating factor for neonatal sepsis: Childhood outcomes at 5 years. Arch. Dis. Child. Fetal Neonatal Ed. 2015, 100, F320–F326. [Google Scholar] [CrossRef]
- Bo, L.; Wang, F.; Zhu, J.; Li, J.; Deng, X. Granulocyte-colony stimulating factor (G-CSF) and granulocyte-macrophage colony stimulating factor (GM-CSF) for sepsis: A meta-analysis. Crit. Care (Lond. Engl.) 2011, 15, R58. [Google Scholar] [CrossRef] [Green Version]
- Fink, M.P.; Warren, H.S. Strategies to improve drug development for sepsis. Nat. Rev. Drug Discov. 2014, 13, 741–758. [Google Scholar] [CrossRef]
- Guo, Y.; Luan, L.; Patil, N.K.; Wang, J.; Bohannon, J.K.; Rabacal, W.; Fensterheim, B.A.; Hernandez, A.; Sherwood, E.R. IL-15 Enables Septic Shock by Maintaining NK Cell Integrity and Function. J. Immunol. 2017, 198, 1320–1333. [Google Scholar] [CrossRef] [Green Version]
- Inoue, S.; Unsinger, J.; Davis, C.G.; Muenzer, J.T.; Ferguson, T.A.; Chang, K.; Osborne, D.F.; Clark, A.T.; Coopersmith, C.M.; McDunn, J.E.; et al. IL-15 Prevents Apoptosis, Reverses Innate and Adaptive Immune Dysfunction, and Improves Survival in Sepsis. J. Immunol. 2009, 184, 1401–1409. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Luan, L.; Rabacal, W.; Bohannon, J.K.; Fensterheim, B.A.; Hernandez, A.; Sherwood, E.R. IL-15 Superagonist–Mediated Immunotoxicity: Role of NK Cells and IFN-γ. J. Immunol. 2015, 195, 2353–2364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattei, F.; Patil, N.K.; Luan, L.; Bohannon, J.K.; Guo, Y.; Hernandez, A.; Fensterheim, B.; Sherwood, E.R. IL-15 Superagonist Expands mCD8+ T, NK and NKT Cells after Burn Injury but Fails to Improve Outcome during Burn Wound Infection. PLoS ONE 2016, 11, e0148452. [Google Scholar]
- Malik, A.; Kanneganti, T.D. Function and regulation of IL-1alpha in inflammatory diseases and cancer. Immunol. Rev. 2018, 281, 124–137. [Google Scholar] [CrossRef]
- Ge, Y.; Huang, M.; Yao, Y.M. Recent advances in the biology of IL-1 family cytokines and their potential roles in development of sepsis. Cytokine Growth Factor Rev. 2019, 45, 24–34. [Google Scholar] [CrossRef]
- Benjamin, J.T.; Moore, D.J.; Bennett, C.; van der Meer, R.; Royce, A.; Loveland, R.; Wynn, J.L. Cutting Edge: IL-1alpha and Not IL-1beta Drives IL-1R1-Dependent Neonatal Murine Sepsis Lethality. J. Immunol. 2018, 201, 2873–2878. [Google Scholar] [CrossRef] [Green Version]
- Cross, A.S. IL-18/IL-1/IL-17A axis: A novel therapeutic target for neonatal sepsis? Cytokine 2016, 86, 1–3. [Google Scholar] [CrossRef]
- Pfeiffer, D.; Rossmanith, E.; Lang, I.; Falkenhagen, D. miR-146a, miR-146b, and miR-155 increase expression of IL-6 and IL-8 and support HSP10 in an In vitro sepsis model. PLoS ONE 2017, 12, e0179850. [Google Scholar] [CrossRef] [Green Version]
- Qiu, X.; Zhang, L.; Tong, Y.; Qu, Y.; Wang, H.; Mu, D. Interleukin-6 for early diagnosis of neonatal sepsis with premature rupture of the membranes: A meta-analysis. Medicine 2018, 97, e13146. [Google Scholar] [CrossRef]
- Hall, M.W.; Knatz, N.L.; Vetterly, C.; Tomarello, S.; Wewers, M.D.; Volk, H.D.; Carcillo, J.A. Immunoparalysis and nosocomial infection in children with multiple organ dysfunction syndrome. Intensive Care Med. 2010, 37, 525–532. [Google Scholar] [CrossRef] [Green Version]
- Meisel, C.; Schefold, J.C.; Pschowski, R.; Baumann, T.; Hetzger, K.; Gregor, J.; Weber-Carstens, S.; Hasper, D.; Keh, D.; Zuckermann, H.; et al. Granulocyte-macrophage colony-stimulating factor to reverse sepsis-associated immunosuppression: A double-blind, randomized, placebo-controlled multicenter trial. Am. J. Respir. Crit. Care Med. 2009, 180, 640–648. [Google Scholar] [CrossRef] [PubMed]
- Hotchkiss, R.S.; Monneret, G.; Payen, D. Immunosuppression in sepsis: A novel understanding of the disorder and a new therapeutic approach. Lancet Infect. Dis. 2013, 13, 260–268. [Google Scholar] [CrossRef] [Green Version]
- Unsinger, J.; Burnham, C.A.; McDonough, J.; Morre, M.; Prakash, P.S.; Caldwell, C.C.; Dunne, W.M., Jr.; Hotchkiss, R.S. Interleukin-7 ameliorates immune dysfunction and improves survival in a 2-hit model of fungal sepsis. J. Infect. Dis. 2012, 206, 606–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venet, F.; Lukaszewicz, A.-C.; Payen, D.; Hotchkiss, R.; Monneret, G. Monitoring the immune response in sepsis: A rational approach to administration of immunoadjuvant therapies. Curr. Opin. Immunol. 2013, 25, 477–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marciano, B.E.; Wesley, R.; De Carlo, E.S.; Anderson, V.L.; Barnhart, L.A.; Darnell, D.; Malech, H.L.; Gallin, J.I.; Holland, S.M. Long-Term Interferon- Therapy for Patients with Chronic Granulomatous Disease. Clin. Infect. Dis. 2004, 39, 692–699. [Google Scholar] [CrossRef]
- Kalvelage, C.; Zacharowski, K.; Bauhofer, A.; Gockel, U.; Adamzik, M.; Nierhaus, A.; Kujath, P.; Eckmann, C.; Pletz, M.W.; Bracht, H.; et al. Personalized medicine with IgGAM compared with standard of care for treatment of peritonitis after infectious source control (the PEPPER trial): Study protocol for a randomized controlled trial. Trials 2019, 20, 156. [Google Scholar] [CrossRef]
- Schlosser, K.; Wang, J.P.; Dos Santos, C.; Walley, K.R.; Marshall, J.; Fergusson, D.A.; Winston, B.W.; Granton, J.; Watpool, I.; Stewart, D.J.; et al. Effects of Mesenchymal Stem Cell Treatment on Systemic Cytokine Levels in a Phase 1 Dose Escalation Safety Trial of Septic Shock Patients. Crit. Care Med. 2019, 47, 918–925. [Google Scholar] [CrossRef]
- Steinhagen, F.; Schmidt, S.V.; Schewe, J.C.; Peukert, K.; Klinman, D.M.; Bode, C. Immunotherapy in sepsis—Brake or accelerate? Pharmacol. Ther. 2020, 107476. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. Immunotherapeutic implications of IL-6 blockade for cytokine storm. Immunotherapy 2016, 8, 959–970. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.-D.; Li, T.; Ji, X.-Y. Dendritic Cells in Sepsis: Pathological Alterations and Therapeutic Implications. J. Immunol. Res. 2017, 2017, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolling, Y.; Salva, S.; Villena, J.; Alvarez, S. Are the immunomodulatory properties of Lactobacillus rhamnosus CRL1505 peptidoglycan common for all Lactobacilli during respiratory infection in malnourished mice? PLoS ONE 2018, 13, e0194034. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Immunotherapy Agents | Major Functions | Ref. |
|---|---|---|
| FLT3L | Increasing the numbers of DCs | [31,33,34,36,71,72] |
| BCL-2 | Inhibiting Fas-mediated DC apoptosis | [73] |
| CD40L | Inhibiting Fas-mediated DC apoptosis | [47,48] |
| TRANCE | Inhibiting Fas-mediated DC apoptosis | [50] |
| Histamine | Inhibiting DC apoptosis | [51] |
| Anti-HMGB1 antibody | Reducing cytokine storm | [52,53,54] |
| Anti-CD155 antibody | Reverse DC dysfunction | [55] |
| Anti-C5a antibody | Improving survival of sepsis | [37] |
| TLR2-derived peptide | Promoting DC maturation | [74] |
| sPLA2 | Increasing the IFN-γsecretion | [63,64,75] |
| miR-142-3p | Promoting the expression of IL-6 and then reducing endotoxin-mediated mortality | [65] |
| SHARPIN | Induction of Th1 differentiation by DCs | [67,68] |
| TLR4 agonist | Inhibiting LPS-induced cytokine production | [57,59] |
| Glucocorticoids | Reducing IL-12 production of DCs | [69,70] |
| CYT387 | Inhibiting LPS-induced cytokine production | [76] |
| Immunotherapy | Major Functions | Ref. |
|---|---|---|
| GM-CSF | Improving the production and function of neutrophils and monocytes. | [120,121] |
| IL-7 | Inducing the proliferation of naive and memory T cells; decreasing Sepsis-induced lymphocyte apoptosis and reversing sepsis-induced depression of interferon γ | [89,122,123] |
| IL-15 | Increasing NK cell, T cell, NKT cell proliferation and activation | [111,112] |
| IFN-γ | Reversing monocyte dysfunction; Increasing the numbers of IL-17-expressing CD4+ T cells | [99,124,125] |
| IgGAM | Improving pathogen recognition and anti-apoptotic effects | [126] |
| Mesenchymalstem cells | Augmenting bacterial clearance | [127,128] |
| PD1/PDL1-specific antibodies | Improving IFN-γ production and decreasing apoptosis of T cells. | [96] |
| IL-1ra | Preventing IL-1β-induced septic shock | [115] |
| anti-IL-6 antibody | Improving survival in sepsis model. | [129] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, G.; Li, X.; Zhang, L.; Elgaili Abdalla, A.; Teng, T.; Li, Y. Crosstalk between Dendritic Cells and Immune Modulatory Agents against Sepsis. Genes 2020, 11, 323. https://doi.org/10.3390/genes11030323
Wang G, Li X, Zhang L, Elgaili Abdalla A, Teng T, Li Y. Crosstalk between Dendritic Cells and Immune Modulatory Agents against Sepsis. Genes. 2020; 11(3):323. https://doi.org/10.3390/genes11030323
Chicago/Turabian StyleWang, Guoying, Xianghui Li, Lei Zhang, Abualgasim Elgaili Abdalla, Tieshan Teng, and Yanzhang Li. 2020. "Crosstalk between Dendritic Cells and Immune Modulatory Agents against Sepsis" Genes 11, no. 3: 323. https://doi.org/10.3390/genes11030323
APA StyleWang, G., Li, X., Zhang, L., Elgaili Abdalla, A., Teng, T., & Li, Y. (2020). Crosstalk between Dendritic Cells and Immune Modulatory Agents against Sepsis. Genes, 11(3), 323. https://doi.org/10.3390/genes11030323