Genome-Wide Transcriptomic Analysis Reveals a Regulatory Network of Oxidative Stress-Induced Flowering Signals Produced in Litchi Leaves


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experiment Procedures
2.2. RNA Isolation, Library Construction and Sequencing
2.3. RNA-Seq Data Analysis
2.4. Quantitative Real-Time PCR Analysis
2.5. Construction of Gene Co-expression Networks
2.6. Statistical Analysis
3. Results
3.1. MV-Generated ROS Promote Flowering
3.2. Digital Transcriptomic Analysis
3.3. Identification of Differentially Expressed Genes
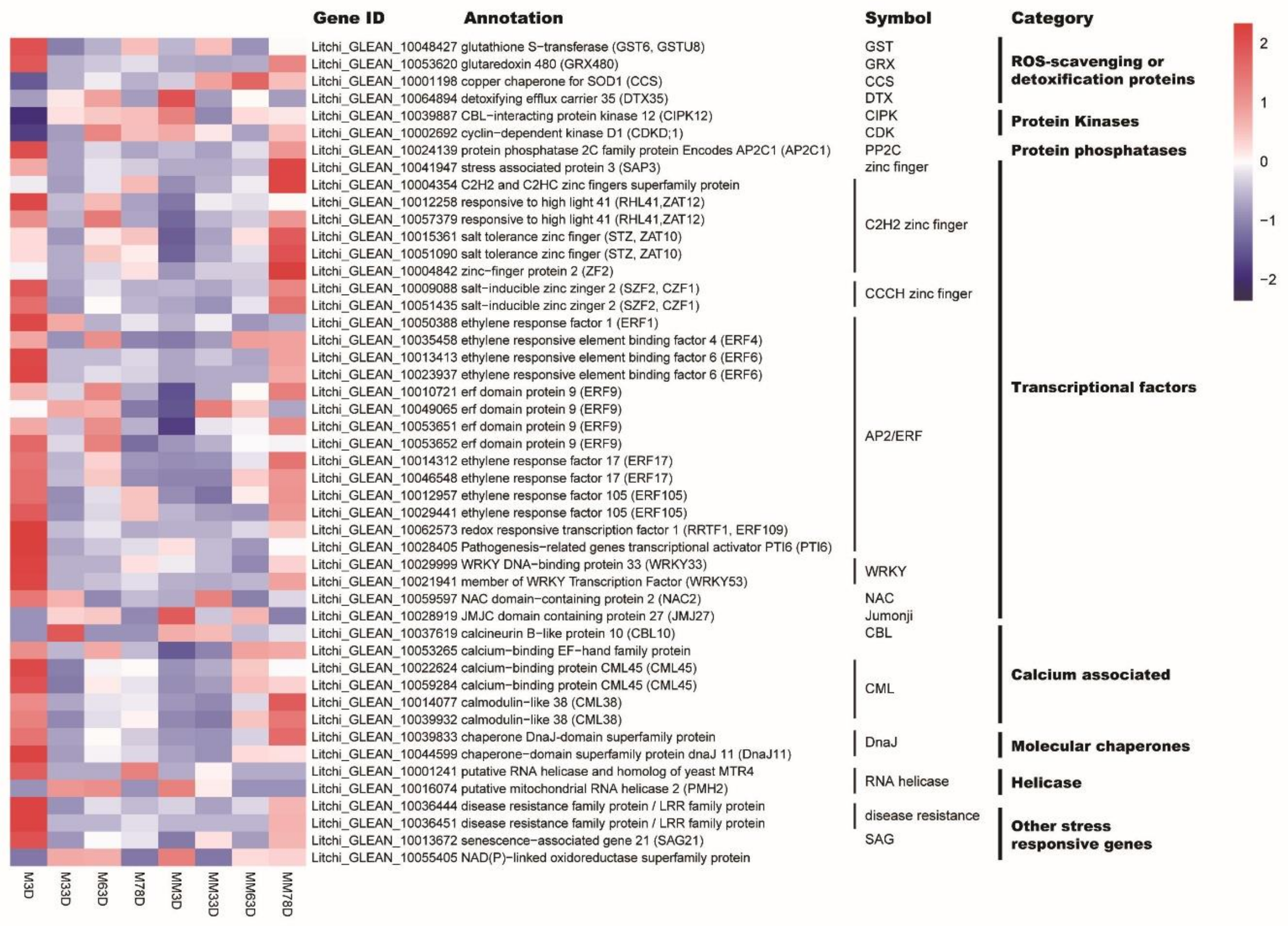
3.4. Identification of Genes Potentially Involved in Oxidative Stress-Induced Flowering
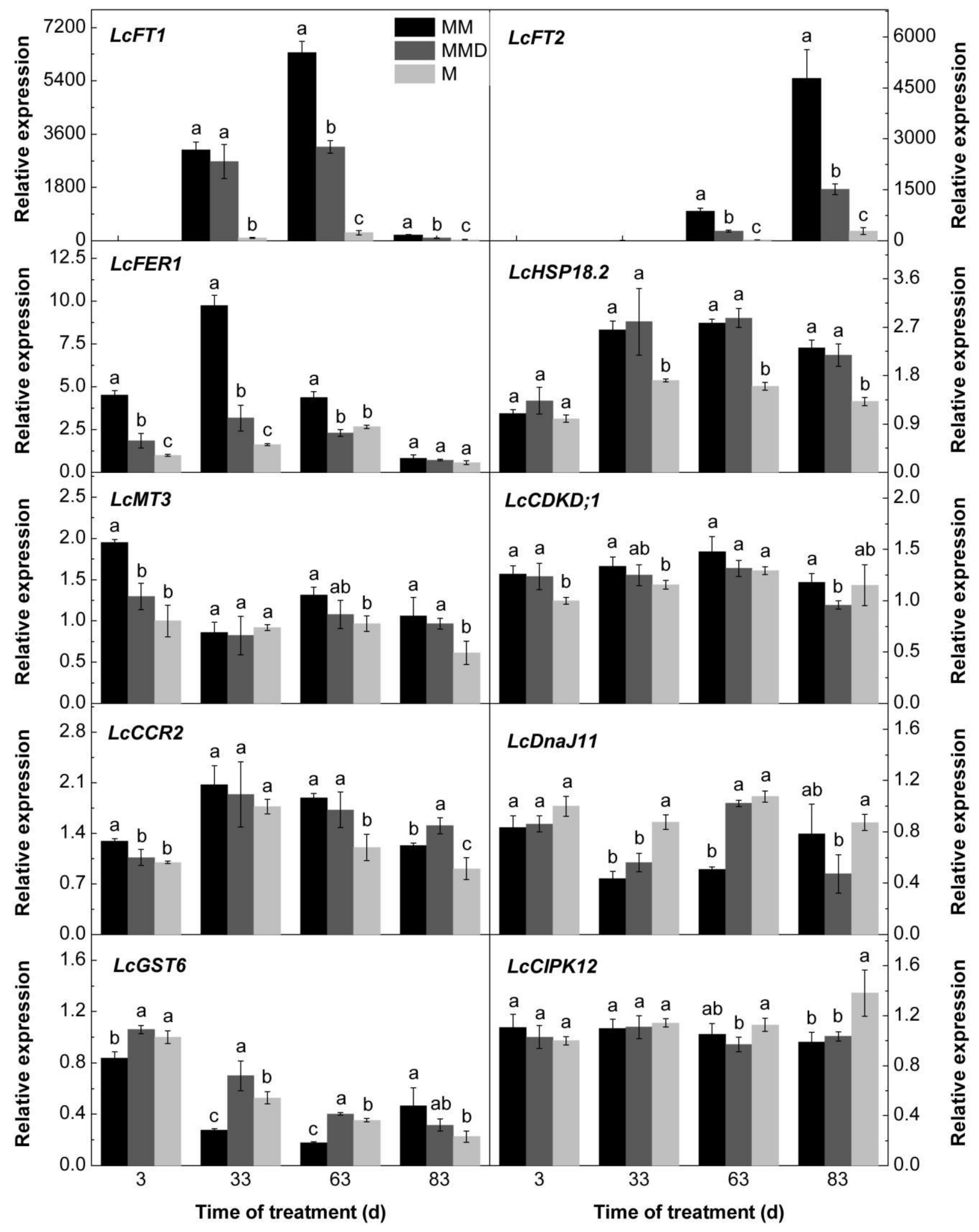
3.5. Confirm Gene Expression Using Real-time Quantitative Reverse Transcription PCR
3.6. Expression Levels of the Candidate Genes in Response to ROS and the ROS Scavenger
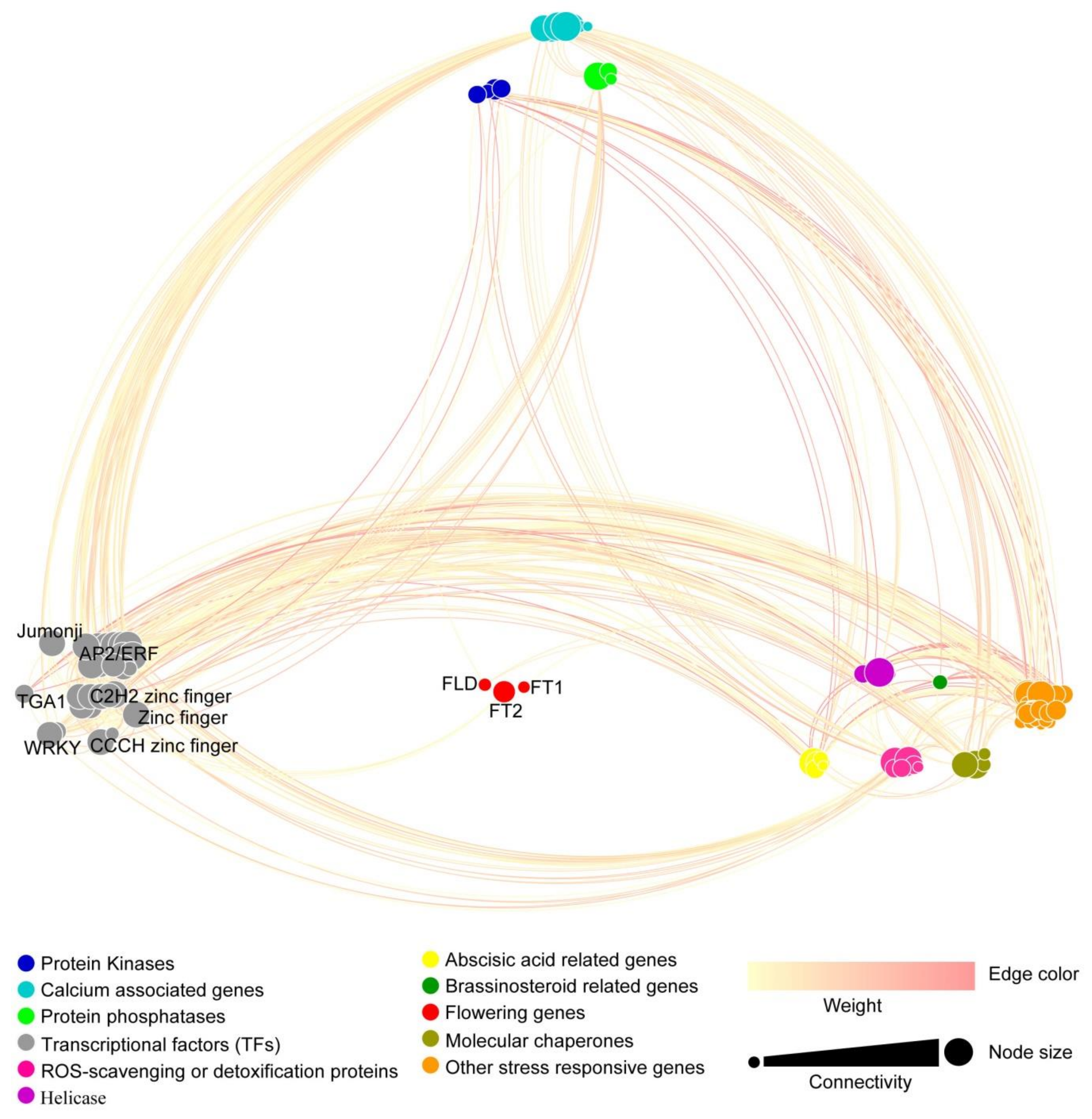
3.7. Co-Expression Network of the DEGs Expressed in Leaves
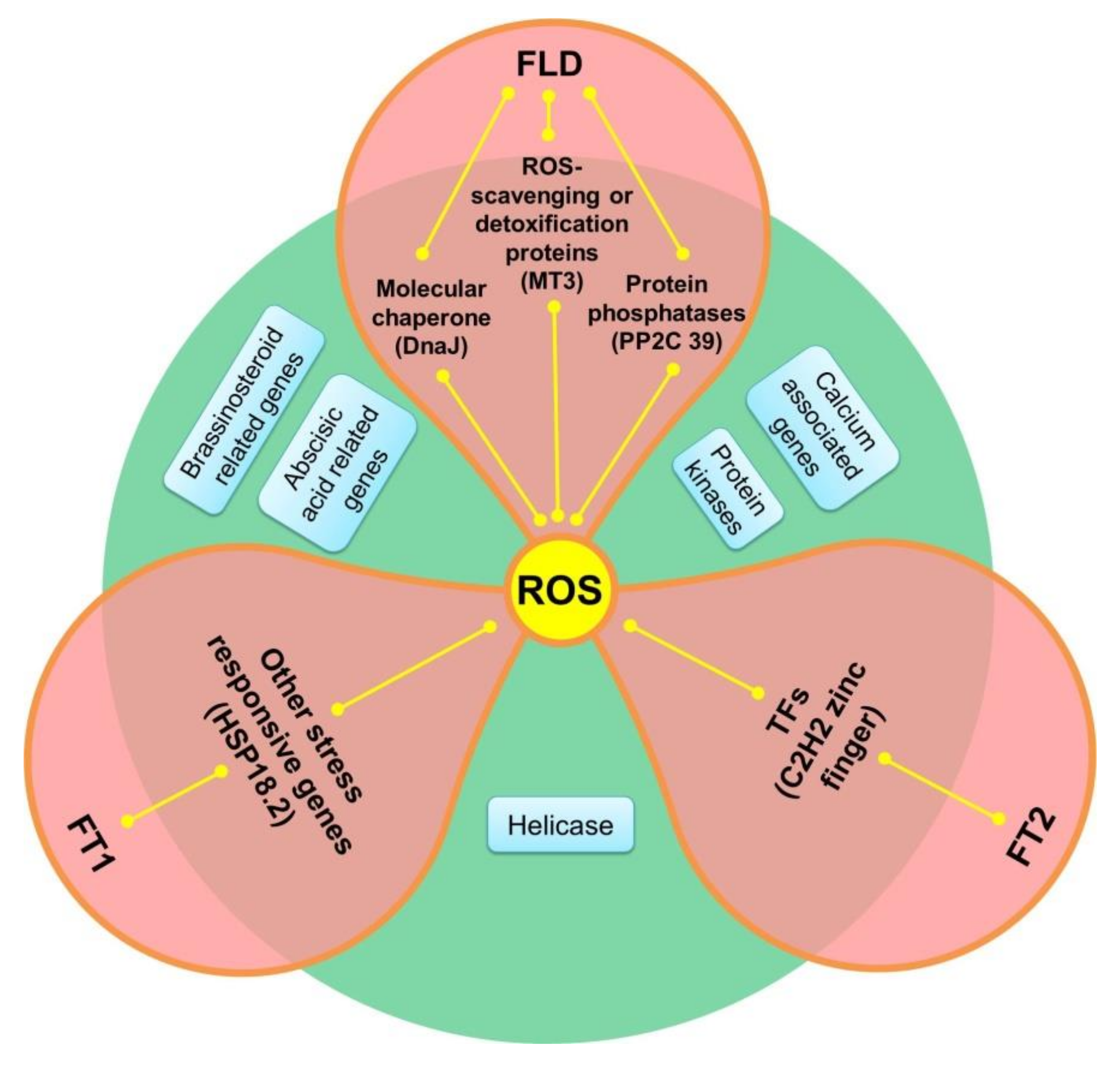
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Menzel, C.M.; Simpson, D.R. Effect of temperature on growth and flowering of litchi (Litchi chinensis Sonn.) cultivars. J. Hortic. Sci. 1988, 63, 349–360. [Google Scholar] [CrossRef]
- Chen, H.B.; Huang, H.B. Low temperature requirements for floral induction in lychee. Acta Hortic. 2005, 665, 195–202. [Google Scholar] [CrossRef]
- Zhou, B.Y.; Chen, H.B.; Wu, C.B. An overview on natural triggers and stress signals in relation to flowering in Litchi chinensis and Dimocarpus longan. Acta Hortic. 2014, 1029, 137–144. [Google Scholar] [CrossRef]
- Zhang, H.; Shen, J.; Wei, Y.; Chen, H. Transcriptome profiling of litchi leaves in response to low temperature reveals candidate regulatory genes and key metabolic events during floral induction. BMC Genom. 2017, 18, 363. [Google Scholar] [CrossRef] [Green Version]
- Corbesier, L.; Vincent, C.; Jang, S.; Fornara, F.; Fan, Q.; Searle, I.; Giakountis, A.; Farrona, S.; Gissot, L.; Turnbull, C.; et al. FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis. Science 2007, 316, 1030–1033. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.J.; Klejnot, J.; Yu, X.H.; Liu, X.M.; Lin, C.T. Florigen (II): It is a mobile protein. J. Integr. Plant Biol. 2007, 49, 1665–1669. [Google Scholar] [CrossRef]
- Lin, M.K.; Belanger, H.; Lee, Y.J.; Varkonyi-Gasic, E.; Taoka, K.I.; Miura, E.; Xoconostle-Cazares, B.; Gendler, K.; Jorgensen, R.A.; Phinney, B.; et al. Flowering Locus T protein may act as the long-distance florigenic signal in the cucurbits. Plant Cell 2007, 19, 1488–1506. [Google Scholar] [CrossRef] [Green Version]
- Tamaki, S.; Matsuo, S.; Wong, H.L.; Yokoi, S.; Shimamoto, K. Hd3a protein is a mobile flowering signal in rice. Science 2007, 316, 1033–1036. [Google Scholar] [CrossRef]
- Zhou, B.; Li, N.; Zhang, Z.; Huang, X.; Chen, H.; Hu, Z.; Pang, X.; Liu, W.; Lu, Y. Hydrogen peroxide and nitric oxide promote reproductive growth in Litchi chinensis. Biol. Plant. 2012, 56, 321–329. [Google Scholar] [CrossRef]
- Liu, W.W.; Kim, H.J.; Chen, H.B.; Lu, X.Y.; Zhou, B.Y. Identification of MV-generated ROS responsive EST clones in floral buds of Litchi chinensis Sonn. Plant Cell Rep. 2013, 32, 1361–1372. [Google Scholar] [CrossRef]
- Lu, X.; Li, J.; Chen, H.; Hu, J.; Liu, P.; Zhou, B. RNA-seq analysis of apical meristem reveals integrative regulatory network of ROS and chilling potentially related to flowering in Litchi chinensis. Sci. Rep. 2017, 7, 10183. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Song, F.; Zhu, J.; Zhang, S.; Yang, Y.; Chen, T.; Tang, B.; Dong, L.; Ding, N.; Zhang, Q.; et al. GSA: Genome sequence archive. Genom. Proteom. Bioinform. 2017, 15, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhao, W.; Xiao, J.; Bao, Y.; Wang, F.; Hao, L.; Zhu, J.; Chen, T.; Zhang, S.; Chen, X.; et al. Database resources of the BIG Data Center in 2019. Nucleic Acids Res. 2019, 47, D8–D14. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Chan, Z. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudawan, B.; Chang, C.S.; Chao, H.F.; Ku, M.S.; Yen, Y.F. Hydrogen cyanamide breaks grapevine bud dormancy in the summer through transient activation of gene expression and accumulation of reactive oxygen and nitrogen species. BMC Plant Biol. 2016, 16, 202. [Google Scholar] [CrossRef] [Green Version]
- Takeno, K. Stress-Induced Flowering. In Abiotic Stress Responses in Plants: Metabolism, Productivity and Sustainability; Ahmad, P., Prasad, M.N.V., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 331–346. [Google Scholar]
- Yang, Z.; Zhong, X.; Fan, Y.; Wang, H.; Li, J.; Huang, X. Burst of reactive oxygen species in pedicel-mediated fruit abscission after carbohydrate supply was cut off in longan (Dimocarpus longan). Front. Plant Sci. 2015, 6, 360. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Shi, X.; Qian, M.; Zheng, L.; Lian, C.; Xia, Y.; Shen, Z. Copper-induced hydrogen peroxide upregulation of a metallothionein gene, OsMT2c, from Oryza sativa L. confers copper tolerance in Arabidopsis thaliana. J. Hazard. Mater. 2015, 294, 99–108. [Google Scholar] [CrossRef]
- Ding, F.; Zhang, S.; Chen, H.; Su, Z.; Zhang, R.; Xiao, Q.; Li, H. Promoter difference of LcFT1 is a leading cause of natural variation of flowering timing in different litchi cultivars (Litchi chinensis Sonn.). Plant Sci. 2015, 241, 128–137. [Google Scholar] [CrossRef]
- Shen, J.; Xiao, Q.; Qiu, H.; Chen, C.; Chen, H. Integrative effect of drought and low temperature on litchi (Litchi chinensis Sonn.) floral initiation revealed by dynamic genome-wide transcriptome analysis. Sci. Rep. 2016, 6, 32005. [Google Scholar] [CrossRef]
- Sudre, D.; Gutierrez-Carbonell, E.; Lattanzio, G.; Rellán-Álvarez, R.; Gaymard, F.; Wohlgemuth, G.; Fiehn, O.; Álvarez-Fernández, A.; Zamarreño, A.M.; Bacaicoa, E.; et al. Iron-dependent modifications of the flower transcriptome, proteome, metabolome, and hormonal content in an Arabidopsis ferritin mutant. J. Exp. Bot. 2013, 64, 2665–2688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Streitner, C.; Danisman, S.; Wehrle, F.; Schöning, J.C.; Alfano, J.R.; Staiger, D. The small glycine-rich RNA binding protein AtGRP7 promotes floral transition in Arabidopsis thaliana. Plant J. 2008, 56, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.H.; Kwak, K.J.; Kim, M.K.; Park, S.J.; Yang, K.Y.; Kang, H. Expression of Arabidopsis glycine-rich RNA-binding protein AtGRP2 or AtGRP7 improves grain yield of rice (Oryza sativa) under drought stress conditions. Plant Sci. 2014, 214, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Hassinen, V.H.; Tervahauta, A.I.; Schat, H.; Kärenlampi, S.O. Plant metallothioneins-metal chelators with ROS scavenging activity? Plant Biol. 2011, 13, 225–232. [Google Scholar] [CrossRef]
- Lee, J.; Donghwan, S.; Won-yong, S.; Inhwan, H.; Youngsook, L. Arabidopsis metallothioneins 2a and 3 enhance resistance to cadmium when expressed in Vicia faba guard cells. Plant Mol. Biol. 2004, 54, 805–815. [Google Scholar] [CrossRef]
- Xue, T.; Li, X.; Zhu, W.; Wu, C.; Yang, G.; Zheng, C. Cotton metallothionein GhMT3a, a reactive oxygen species scavenger, increased tolerance against abiotic stress in transgenic tobacco and yeast. J. Exp. Bot. 2008, 60, 339–349. [Google Scholar] [CrossRef]
- Mekawy, A.M.M.; Assaha, D.V.; Munehiro, R.; Kohnishi, E.; Nagaoka, T.; Ueda, A.; Saneoka, H. Characterization of type 3 metallothionein-like gene (OsMT-3a) from rice, revealed its ability to confer tolerance to salinity and heavy metal stresses. Environ. Exp. Bot. 2018, 147, 157–166. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Percentage of Flowering Trees (%) | Length of Panicle/cm | Width of Panicle/cm | Max Spike Nodes per Panicle | No. of Flowers per Panicle |
|---|---|---|---|---|---|
| MM | 80 | 12.61 ± 1.97 | 5.68 ± 1.79 | 11.35 ± 1.10 * | 749.48 ± 180.41 * |
| MMD | 80 | 10.99 ± 1.19 | 2.62 ± 0.53 | 7.20 ± 1.06 | 202.32 ± 73.29 |
| M | 0 | - | - | - | - |
| Sample | Raw Reads | Clean Reads | Clean Reads/Raw Reads (%) | Alignment Rate (%) |
|---|---|---|---|---|
| M3D-1 | 7838012 | 6906535 | 88.115902 | 85.87 |
| M3D-2 | 7885494 | 7206931 | 91.394794 | 87.22 |
| M33D-1 | 7768367 | 7505505 | 96.616252 | 87.9 |
| M33D-2 | 6978483 | 6833631 | 97.924305 | 88.35 |
| M63D-1 | 10799631 | 10360566 | 95.934444 | 88.45 |
| M63D-2 | 9667348 | 9354050 | 96.759215 | 88.85 |
| M78D-1 | 8278464 | 7879395 | 95.179432 | 85.84 |
| M78D-2 | 9337570 | 8769018 | 93.911135 | 88.83 |
| MM3D-1 | 7864489 | 7701724 | 97.93038 | 87.84 |
| MM3D-2 | 11179790 | 10754878 | 96.199285 | 86.8 |
| MM33D-1 | 8149987 | 7930713 | 97.309517 | 86.79 |
| MM33D-2 | 5167534 | 4951442 | 95.818276 | 89.31 |
| MM63D-1 | 9581811 | 8874327 | 92.616385 | 87.28 |
| MM63D-2 | 12149900 | 11554502 | 95.099565 | 85.2 |
| MM78D-1 | 9088200 | 8815661 | 97.001177 | 89.61 |
| MM78D-2 | 11964902 | 10606541 | 88.64712 | 86.02 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, X.; Yu, S.; Lü, P.; Chen, H.; Zhong, S.; Zhou, B. Genome-Wide Transcriptomic Analysis Reveals a Regulatory Network of Oxidative Stress-Induced Flowering Signals Produced in Litchi Leaves. Genes 2020, 11, 324. https://doi.org/10.3390/genes11030324
Lu X, Yu S, Lü P, Chen H, Zhong S, Zhou B. Genome-Wide Transcriptomic Analysis Reveals a Regulatory Network of Oxidative Stress-Induced Flowering Signals Produced in Litchi Leaves. Genes. 2020; 11(3):324. https://doi.org/10.3390/genes11030324
Chicago/Turabian StyleLu, Xingyu, Sheng Yu, Peitao Lü, Houbin Chen, Silin Zhong, and Biyan Zhou. 2020. "Genome-Wide Transcriptomic Analysis Reveals a Regulatory Network of Oxidative Stress-Induced Flowering Signals Produced in Litchi Leaves" Genes 11, no. 3: 324. https://doi.org/10.3390/genes11030324
APA StyleLu, X., Yu, S., Lü, P., Chen, H., Zhong, S., & Zhou, B. (2020). Genome-Wide Transcriptomic Analysis Reveals a Regulatory Network of Oxidative Stress-Induced Flowering Signals Produced in Litchi Leaves. Genes, 11(3), 324. https://doi.org/10.3390/genes11030324