AQP3 Facilitates Proliferation and Adipogenic Differentiation of Porcine Intramuscular Adipocytes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Care
2.2. Cell Culture
2.3. Transfection with siRNA
2.4. RNA Isolation and RT-qPCR
2.5. Western Blot
2.6. Oil Red O Staining
2.7. EdU Staining
2.8. CCK-8 Assay
2.9. Statistical Analysis
3. Results
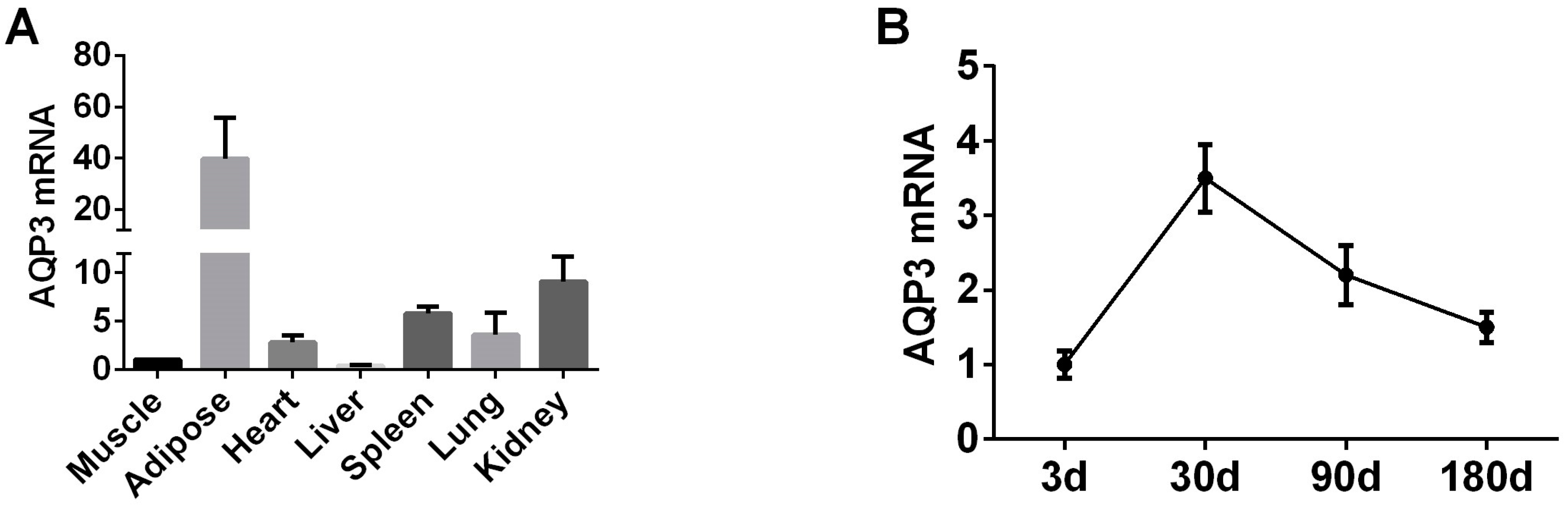
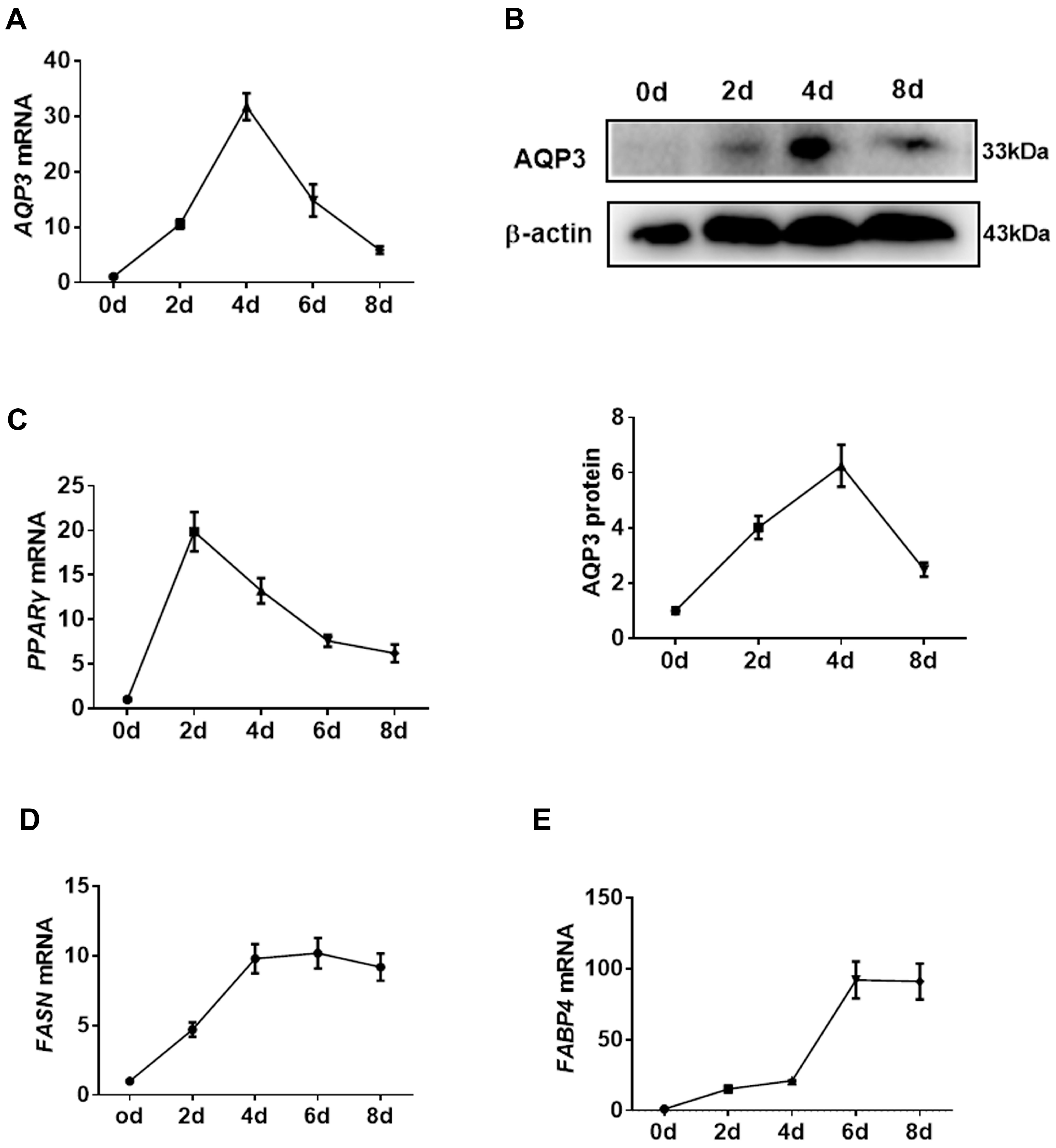
3.1. AQP3 Is Upregulated during Adipogenesis
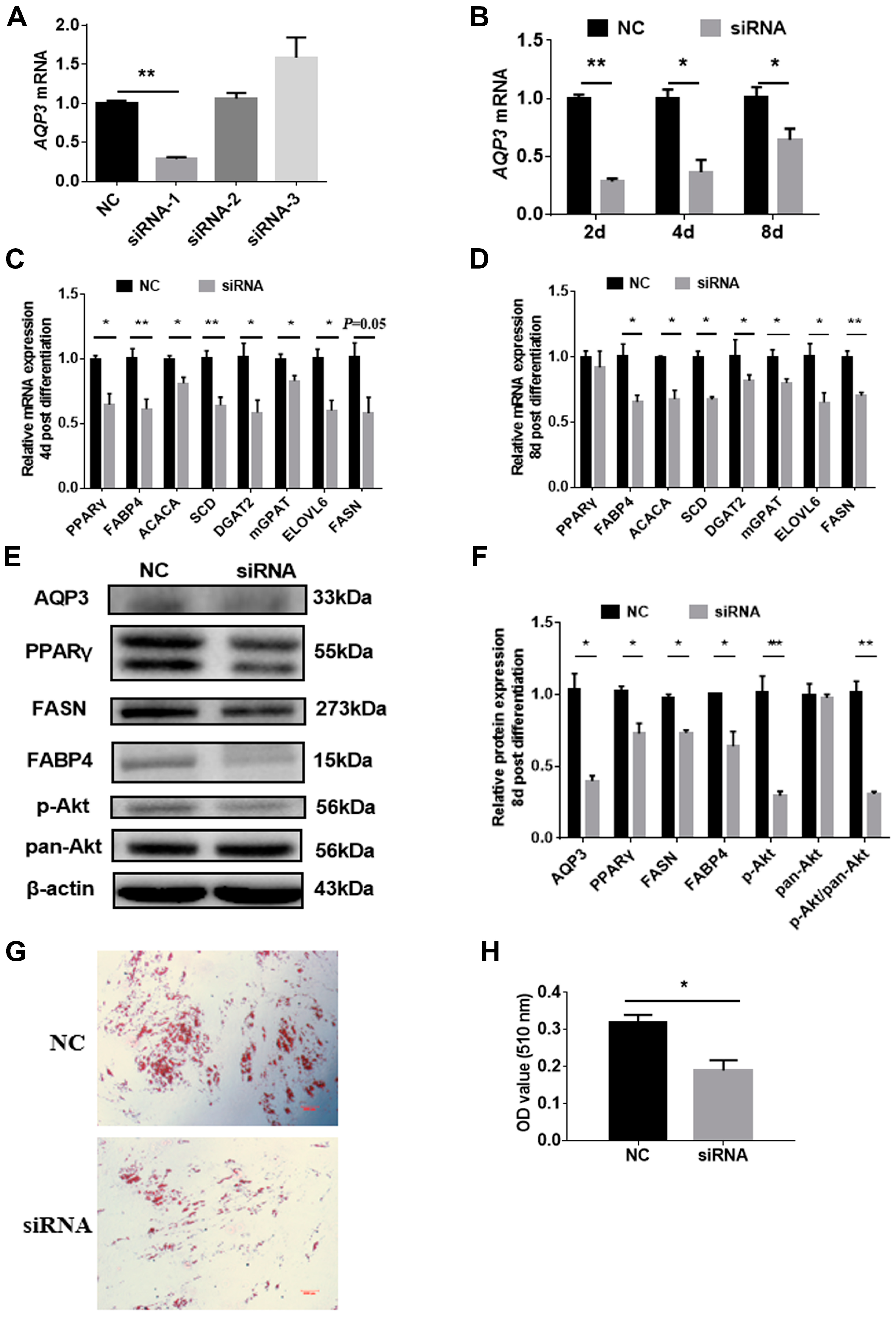
3.2. Knockdown of AQP3 Blunts Adipogenesis
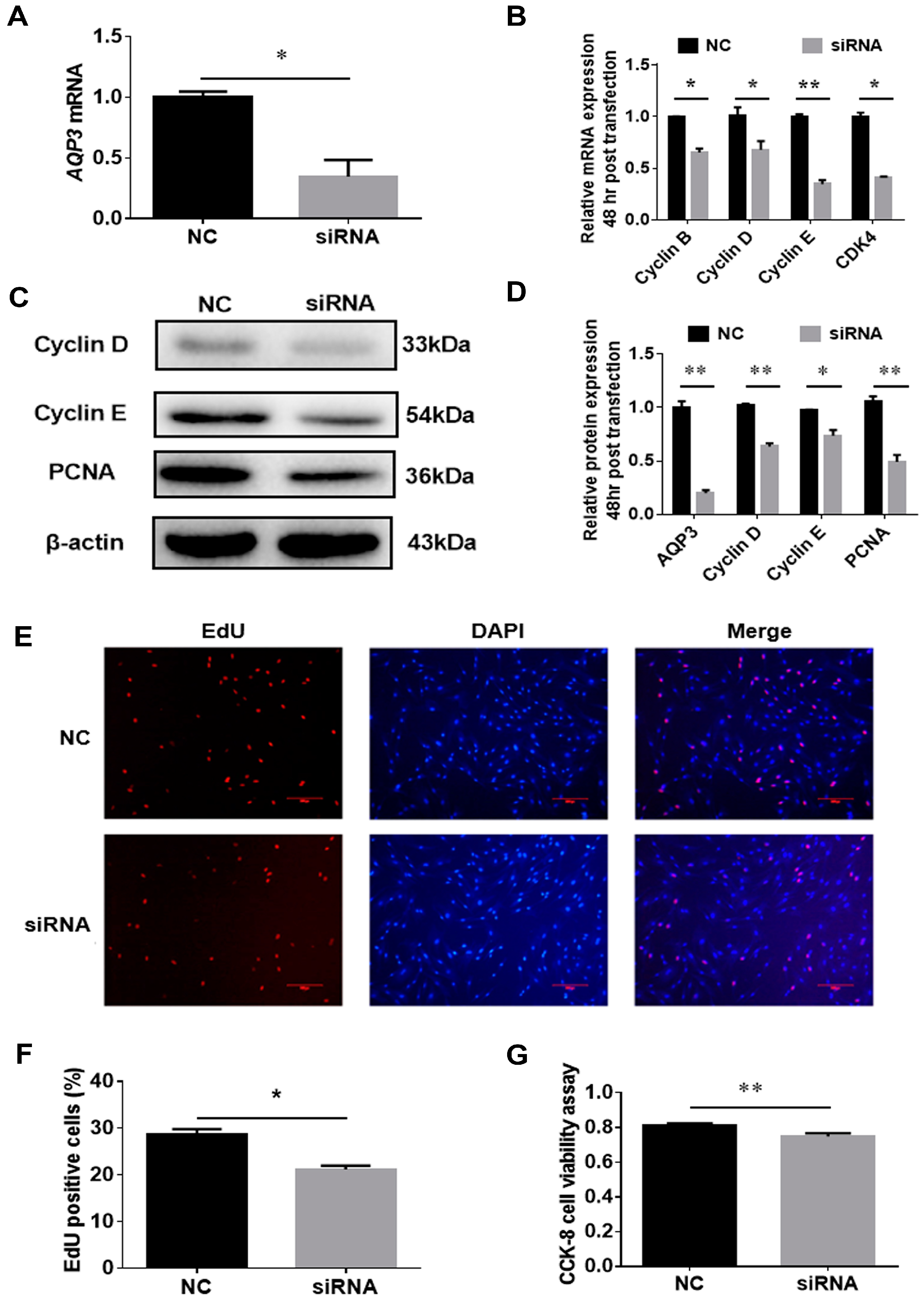
3.3. AQP3 Deletion Inhibits Proliferation
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hausman, G.J.; Basu, U.; Du, M.; Fernyhough-Culver, M.; Dodson, M.V. Intermuscular and intramuscular adipose tissues: Bad vs. good adipose tissues. Adipocyte 2014, 3, 242–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Listrat, A.; Lebret, B.; Louveau, I.; Astruc, T.; Bonnet, M.; Lefaucheur, L.; Picard, B.; Bugeon, J. How Muscle Structure and Composition Influence Meat and Flesh Quality. Scientific World J. 2016, 2016, 3182746. [Google Scholar] [CrossRef] [Green Version]
- Frank, D.; Joo, S.T.; Warner, R. Consumer Acceptability of Intramuscular Fat. Korean J. Food Sci. Anim. Resour. 2016, 36, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, I.V.; Soveral, G. Aquaporins in Obesity. Adv. Exp. Med. Biol. 2017, 969, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Iena, F.M.; Lebeck, J. Implications of Aquaglyceroporin 7 in Energy Metabolism. Int. J. Mol. Sci. 2018, 19, 154. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, A.; Catalan, V.; Gomez-Ambrosi, J.; Garcia-Navarro, S.; Rotellar, F.; Valenti, V.; Silva, C.; Gil, M.J.; Salvador, J.; Burrell, M.A.; et al. Insulin- and leptin-mediated control of aquaglyceroporins in human adipocytes and hepatocytes is mediated via the PI3K/Akt/mTOR signaling cascade. J. Clin. Endocrinol. Metab. 2011, 96, E586–E597. [Google Scholar] [CrossRef] [Green Version]
- Laforenza, U.; Scaffino, M.F.; Gastaldi, G. Aquaporin-10 represents an alternative pathway for glycerol efflux from human adipocytes. PLoS ONE 2013, 8, e54474. [Google Scholar] [CrossRef]
- Madeira, A.; de Almeida, A.; de Graaf, C.; Camps, M.; Zorzano, A.; Moura, T.F.; Casini, A.; Soveral, G. A gold coordination compound as a chemical probe to unravel aquaporin-7 function. Chembiochem 2014, 15, 1487–1494. [Google Scholar] [CrossRef]
- Rodriguez, R.A.; Liang, H.; Chen, L.Y.; Plascencia-Villa, G.; Perry, G. Single-channel permeability and glycerol affinity of human aquaglyceroporin AQP3. Biochim. Biophys. Acta Biomembr. 2019, 1861, 768–775. [Google Scholar] [CrossRef]
- Tardelli, M.; Claudel, T.; Bruschi, F.V.; Moreno-Viedma, V.; Trauner, M. Adiponectin regulates AQP3 via PPARalpha in human hepatic stellate cells. Biochem. Biophys. Res. Commun. 2017, 490, 51–54. [Google Scholar] [CrossRef]
- Rodriguez, A.; Moreno, N.R.; Balaguer, I.; Mendez-Gimenez, L.; Becerril, S.; Catalan, V.; Gomez-Ambrosi, J.; Portincasa, P.; Calamita, G.; Soveral, G.; et al. Leptin administration restores the altered adipose and hepatic expression of aquaglyceroporins improving the non-alcoholic fatty liver of ob/ob mice. Sci. Rep. 2015, 5, 12067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzi, C.; Holdt, L.M.; Fiorito, G.; Tsai, P.C.; Kretschmer, A.; Wahl, S.; Guarrera, S.; Teupser, D.; Spector, T.D.; Iacoviello, L.; et al. Epigenetic Signatures at AQP3 and SOCS3 Engage in Low-Grade Inflammation across Different Tissues. PLoS ONE 2016, 11, e0166015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J. Identification of Key Genes for Differential Deposition and Functional Analysis of AQP3 in Porcine Intramusclar Fat. Master’s Thesis, NWAFU, Yangling, China, May 2018. [Google Scholar]
- Arif, M.; Kitchen, P.; Conner, M.T.; Hill, E.J.; Nagel, D.; Bill, R.M.; Dunmore, S.J.; Armesilla, A.L.; Gross, S.; Carmichael, A.R.; et al. Downregulation of aquaporin 3 inhibits cellular proliferation, migration and invasion in the MDA-MB-231 breast cancer cell line. Oncol. Lett. 2018, 16, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Marlar, S.; Jensen, H.H.; Login, F.H.; Nejsum, L.N. Aquaporin-3 in Cancer. Int. J. Mol. Sci. 2017, 18, 2106. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Tao, C.; Yuan, C.; Ren, J.; Yang, M.; Ying, H. AQP3 small interfering RNA and PLD2 small interfering RNA inhibit the proliferation and promote the apoptosis of squamous cell carcinoma. Mol. Med. Rep. 2017, 16, 1964–1972. [Google Scholar] [CrossRef] [Green Version]
- Xiong, G.; Chen, X.; Zhang, Q.; Fang, Y.; Chen, W.; Li, C.; Zhang, J. RNA interference influenced the proliferation and invasion of XWLC-05 lung cancer cells through inhibiting aquaporin 3. Biochem. Biophys. Res. Commun. 2017, 485, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, B.; Zhang, L.; Chen, L.; Sun, G.; Zhang, Q.; Wang, J.; Zhi, X.; Wang, L.; Xu, Z.; et al. The proliferation impairment induced by AQP3 deficiency is the result of glycerol uptake and metabolism inhibition in gastric cancer cells. Tumour Biol. 2016, 37, 9169–9179. [Google Scholar] [CrossRef]
- Chen, L.; Li, Z.; Zhang, Q.; Wei, S.; Li, B.; Zhang, X.; Zhang, L.; Li, Q.; Xu, H.; Xu, Z. Silencing of AQP3 induces apoptosis of gastric cancer cells via downregulation of glycerol intake and downstream inhibition of lipogenesis and autophagy. OncoTargets Ther. 2017, 10, 2791–2804. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Huang, K.; Chen, F.; Li, W.; Sun, S.; Shi, X.E.; Yang, G. Verification of suitable and reliable reference genes for quantitative real-time PCR during adipogenic differentiation in porcine intramuscular stromal-vascular cells. Animal 2016, 10, 947–952. [Google Scholar] [CrossRef]
- He, L.; Huang, N.; Li, H.; Tian, J.; Zhou, X.; Li, T.; Yao, K.; Wu, G.; Yin, Y. AMPK/α-Ketoglutarate Axis Regulates Intestinal Water and Ion Homeostasis in Young Pigs. J. Agric. Food Chem. 2017, 65, 2287–2298. [Google Scholar] [CrossRef]
- Liu, J.B.; Chen, D.W.; Yu, B.; Mao, X.B. Effect of maternal folic acid supplementation on hepatic one-carbon unit associated gene expressions in newborn piglets. Mol. Biol. Rep. 2011, 38, 3849–3856. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Song, Z.Y.; Pu, L.; Yang, H.; Zheng, J.M.; Zhang, Z.Y.; Shi, X.E.; Yang, G.S. Retinol binding protein 4 affects the adipogenesis of porcine preadipocytes through insulin signaling pathways. Biochem. Cell Biol. 2013, 91, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Zhang, S.; Guan, W.; Chen, F.; Zhang, Y.; Chen, J.; Liu, Y. Metabolic transition of milk triacylglycerol synthesis in response to varying levels of palmitate in porcine mammary epithelial cells. Genes Nutr. 2018, 13, 18. [Google Scholar] [CrossRef]
- Yang, Y.; Ju, D.; Zhang, M.; Yang, G. Interleukin-6 stimulates lipolysis in porcine adipocytes. Endocrine 2008, 33, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Y.H.; Wang, L.; Riaz, H.; Wu, J.B.; Yuan, Y.F.; Han, L.; Wang, Y.L.; Zhao, Y.; Dan, Y.; Huo, L.J. Knockdown of CEBPbeta by RNAi in porcine granulosa cells resulted in S phase cell cycle arrest and decreased progesterone and estradiol synthesis. J. Steroid. Biochem. Mol. Biol. 2014, 143, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Dozois, C.M.; Oswald, E.; Gautier, N.; Serthelon, J.P.; Fairbrother, J.M.; Oswald, I.P. A reverse transcription-polymerase chain reaction method to analyze porcine cytokine gene expression. Vet. Immunol. Immunopathol. 1997, 58, 287–300. [Google Scholar] [CrossRef]
- Li, X.; Lei, T.; Xia, T.; Chen, X.; Feng, S.; Chen, H.; Chen, Z.; Peng, Y.; Yang, Z. Molecular characterization, chromosomal and expression patterns of three aquaglyceroporins (AQP3, 7, 9) from pig. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008, 149, 468–476. [Google Scholar] [CrossRef]
- Kishida, K.; Kuriyama, H.; Funahashi, T.; Shimomura, I.; Kihara, S.; Ouchi, N.; Nishida, M.; Nishizawa, H.; Matsuda, M.; Takahashi, M.; et al. Aquaporin adipose, a putative glycerol channel in adipocytes. J. Biol. Chem. 2000, 275, 20896–20902. [Google Scholar] [CrossRef] [Green Version]
- Maeda, N.; Funahashi, T.; Hibuse, T.; Nagasawa, A.; Kishida, K.; Kuriyama, H.; Nakamura, T.; Kihara, S.; Shimomura, I.; Matsuzawa, Y. Adaptation to fasting by glycerol transport through aquaporin 7 in adipose tissue. Proc. Natl. Acad. Sci. USA 2004, 101, 17801–17806. [Google Scholar] [CrossRef] [Green Version]
- Miranda, M.; Escote, X.; Ceperuelo-Mallafre, V.; Alcaide, M.J.; Simon, I.; Vilarrasa, N.; Wabitsch, M.; Vendrell, J. Paired subcutaneous and visceral adipose tissue aquaporin-7 expression in human obesity and type 2 diabetes: Differences and similarities between depots. J. Clin. Endocrinol. Metab. 2010, 95, 3470–3479. [Google Scholar] [CrossRef] [Green Version]
- Chiadak, J.D.; Arsenijevic, T.; Gregoire, F.; Bolaky, N.; Delforge, V.; Perret, J.; Delporte, C. Involvement of JNK/NFkappaB Signaling Pathways in the Lipopolysaccharide-Induced Modulation of Aquaglyceroporin Expression in 3T3-L1 Cells Differentiated into Adipocytes. Int. J. Mol. Sci. 2016, 17, 1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhary, V.; Olala, L.O.; Kagha, K.; Pan, Z.Q.; Chen, X.; Yang, R.; Cline, A.; Helwa, I.; Marshall, L.; Kaddour-Djebbar, I.; et al. Regulation of the Glycerol Transporter, Aquaporin-3, by Histone Deacetylase-3 and p53 in Keratinocytes. J. Investig. Dermatol. 2017, 137, 1935–1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janani, C.; Ranjitha Kumari, B.D. PPAR γ gene—A review. Diabetes Metab. Syndr. 2015, 9, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Kishida, K.; Shimomura, I.; Nishizawa, H.; Maeda, N.; Kuriyama, H.; Kondo, H.; Matsuda, M.; Nagaretani, H.; Ouchi, N.; Hotta, K.; et al. Enhancement of the aquaporin adipose gene expression by a peroxisome proliferator-activated receptor γ. J. Biol. Chem. 2001, 276, 48572–48579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tardelli, M.; Bruschi, F.V.; Claudel, T.; Moreno-Viedma, V.; Halilbasic, E.; Marra, F.; Herac, M.; Stulnig, T.M.; Trauner, M. AQP3 is regulated by PPARgamma and JNK in hepatic stellate cells carrying PNPLA3 I148M. Sci. Rep. 2017, 7, 14661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiadak, J.D.; Gena, P.; Gregoire, F.; Bolaky, N.; Delforge, V.; Perret, J.; Calamita, G.; Delporte, C. Lipopolysaccharide Modifies Glycerol Permeability and Metabolism in 3T3-L1 Adipocytes. Int. J. Mol. Sci. 2017, 18, 2566. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Jiao, L.; Li, T.; Wang, H.; Wei, W.; Qian, H. Expression of AQP3 and AQP5 as a prognostic marker in triple-negative breast cancer. Oncol. Lett. 2018, 16, 2661–2667. [Google Scholar] [CrossRef] [Green Version]
- Galan-Cobo, A.; Ramirez-Lorca, R.; Serna, A.; Echevarria, M. Overexpression of AQP3 Modifies the Cell Cycle and the Proliferation Rate of Mammalian Cells in Culture. PLoS ONE 2015, 10, e0137692. [Google Scholar] [CrossRef]
- Sarantopoulos, C.N.; Banyard, D.A.; Ziegler, M.E.; Sun, B.; Shaterian, A.; Widgerow, A.D. Elucidating the Preadipocyte and Its Role in Adipocyte Formation: A Comprehensive Review. Stem. Cell Rev. Rep. 2018, 14, 27–42. [Google Scholar] [CrossRef]
- Lebeck, J. Metabolic impact of the glycerol channels AQP7 and AQP9 in adipose tissue and liver. J. Mol. Endocrinol. 2014, 52, R165–R178. [Google Scholar] [CrossRef] [Green Version]
- Madeira, A.; Mosca, A.F.; Moura, T.F.; Soveral, G. Aquaporin-5 is expressed in adipocytes with implications in adipose differentiation. IUBMB Life 2015, 67, 54–60. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Primer Sequences | Production Length/bp |
|---|---|---|---|
| AQP3 | NM_001110172.1 | F: CACCTCCATGGGCTTCAACT | 278 |
| R: TGCCCATTCGCATCTACTCC | |||
| PPARγ | NM_214379 | F: AGGACTACCAAAGTGCCATCAAA | 142 |
| R: GAGGCTTTATCCCCACAGACAC | |||
| aP2 | NM_001002817.1 | F: GAGCACCATAACCTTAGATGGA | 121 |
| R: AAATTCTGGTAGCCGTGACA | |||
| FASN | NM_001099930.1 | F: GTCCTGCTGAAGCCTAACTC | 206 |
| R: TCCTTGGAACCGTCTGTG | |||
| SCD | NM_213781.1 | F: ACAAGAGGCCAAGACAAGTTCC | 142 |
| R: GCTGTAGGGAATGCTGGTTAGTTT | |||
| ACACA | NM_001114269.1 | F: TCCCAGTGCAAGCAGTATG | 211 |
| R: TGCCAATCCACACGAAGAC | |||
| mGPAT | XM_005671462.3 | F: ACTATCTCCTGCTCACTTTCA | 146 |
| R: CGTCTCATCTAGCCTCCGTC | |||
| CD36 | NM_001044622.1 | F: ATCGTGCCTATCCTCTGG | 103 |
| R: CCAGGCCAAGGAGGTTAA | |||
| C/EBPα | XM_003127015.4 | F: AACAACTGAGCCGCGAACTG | 181 |
| R: GCTCCGGCAGTCTTGAGAT | |||
| DGAT2 | NM_001160080.1 | F: GCAGGTGATCTTTGAGGAGG | 140 |
| R: GCTTGGAGTAGGGCATGAG | |||
| ELOVL6 | XM_021100708.1 | F: ACCACATCACTGTGCTCCTC | 95 |
| R: CGAGTGCACGCCATAGTTCA | |||
| Cyclin B | NM_001170768.1 | F: AATCCCTTCTTGTGGTTA | 104 |
| R: CTTAGATGTGGCATACTTG | |||
| Cyclin E | XM_005653265.2 | F: AGAAGGAAAGGGATGCGAAGG | 173 |
| R: CCAAGGCTGATTGCCACACT | |||
| Cyclin D | XM_021082686.1 | F: TACACCGACAACTCCATCCG | 224 |
| R: GAGGGCGGGTTGGAAATGAA | |||
| β-actin | XM_021086047.1 | F: GGACTTCGAGCAGGAGATGG | 138 |
| R: AGGAAGGAGGGCTGGAAGAG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Yang, J.; Yao, Y.; Shi, X.; Yang, G.; Li, X. AQP3 Facilitates Proliferation and Adipogenic Differentiation of Porcine Intramuscular Adipocytes. Genes 2020, 11, 453. https://doi.org/10.3390/genes11040453
Wang X, Yang J, Yao Y, Shi X, Yang G, Li X. AQP3 Facilitates Proliferation and Adipogenic Differentiation of Porcine Intramuscular Adipocytes. Genes. 2020; 11(4):453. https://doi.org/10.3390/genes11040453
Chicago/Turabian StyleWang, Xiaoyu, Jing Yang, Ying Yao, Xin’E Shi, Gongshe Yang, and Xiao Li. 2020. "AQP3 Facilitates Proliferation and Adipogenic Differentiation of Porcine Intramuscular Adipocytes" Genes 11, no. 4: 453. https://doi.org/10.3390/genes11040453
APA StyleWang, X., Yang, J., Yao, Y., Shi, X., Yang, G., & Li, X. (2020). AQP3 Facilitates Proliferation and Adipogenic Differentiation of Porcine Intramuscular Adipocytes. Genes, 11(4), 453. https://doi.org/10.3390/genes11040453