Immunoreactive Proteins in the Esophageal Gland Cells of Anisakis Simplex Sensu Stricto Detected by MALDI-TOF/TOF Analysis


 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Parasite Material
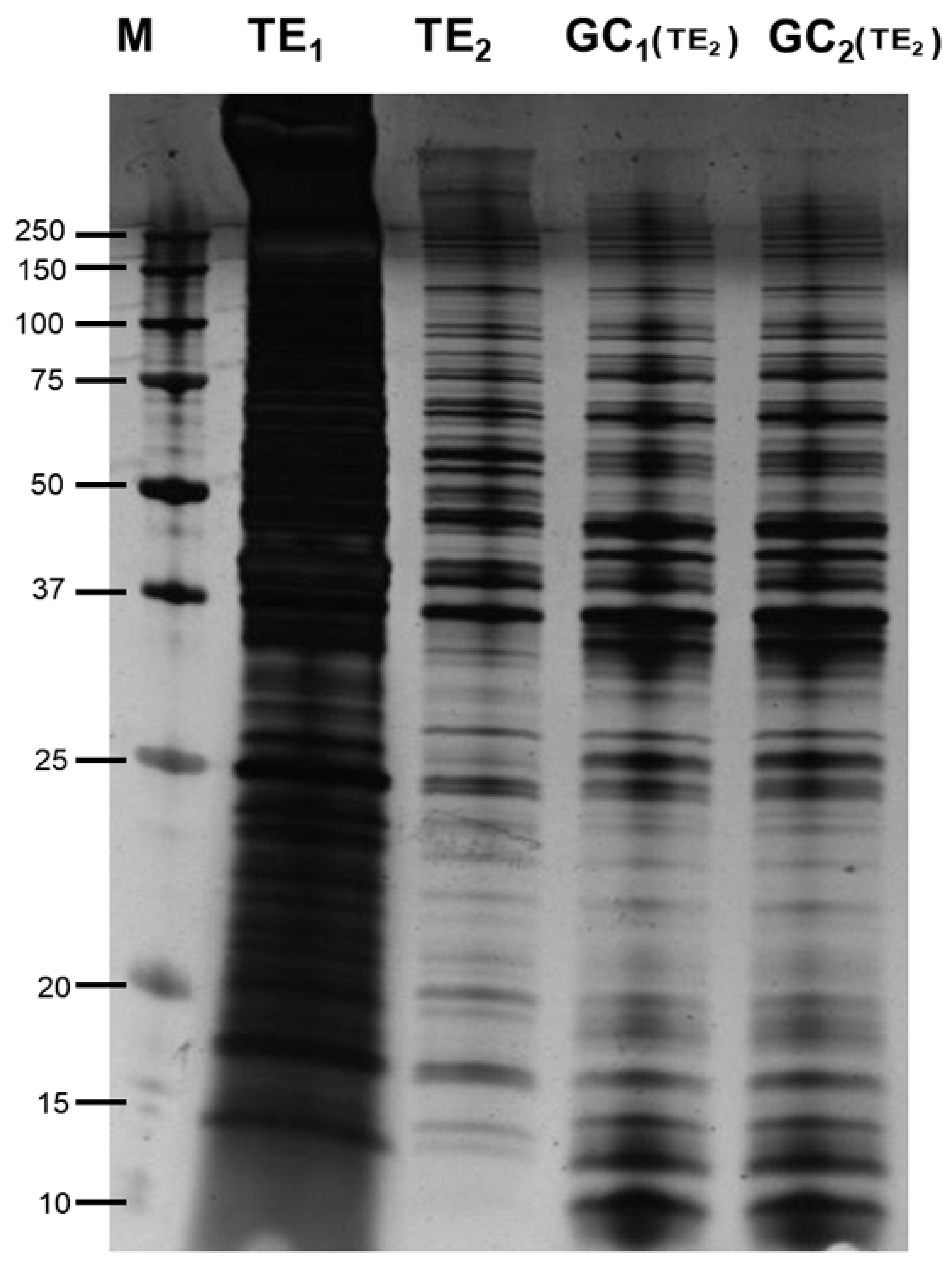
2.2. 1D SDS-PAGE
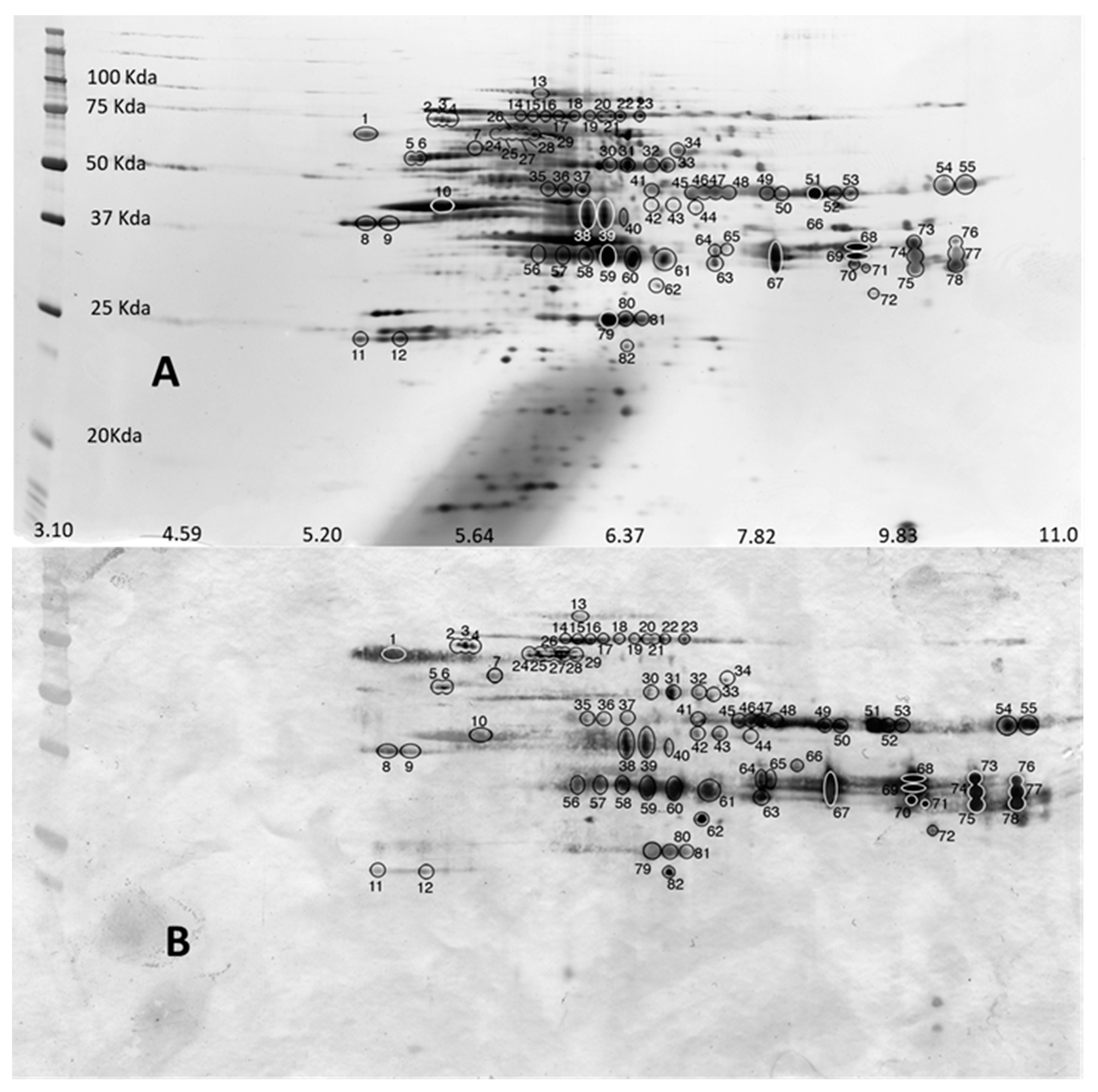
2.3. Two-Dimensional Gel Electrophoresis
2.4. Silver Staining
2.5. Image Acquisition and Analysis
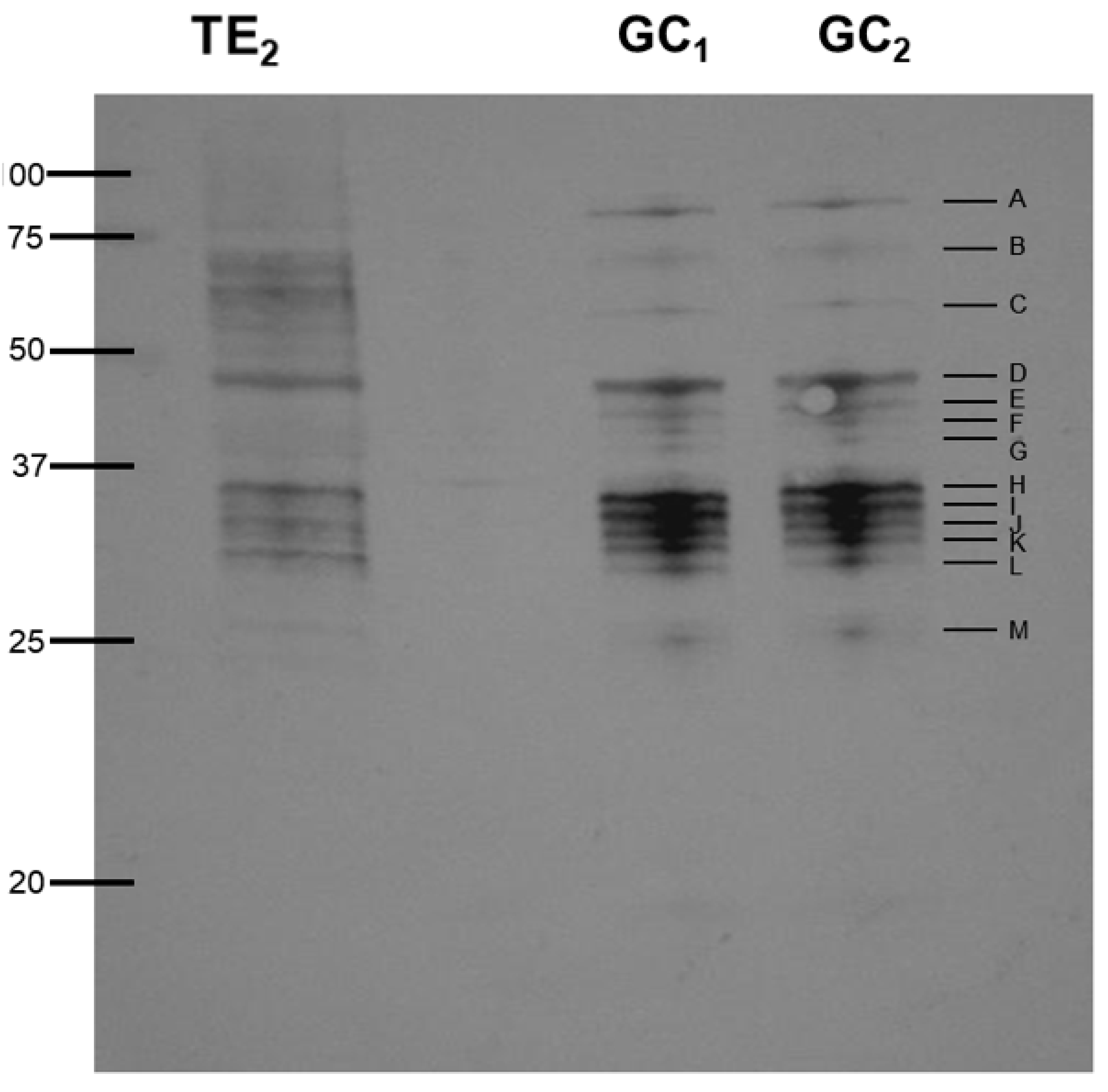
2.6. Immunoblotting
2.7. In-Gel Protein Digestion and Sample Preparation
2.8. MALDI Peptide Mass Fingerprinting, MS/MS Analysis and Database Mining
3. Results
3.1. Silver Staining and Western Blotting
3.2. MALDI Peptide Mass Fingerprinting, MS/MS Analysis and Database Mining
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- González Quijada, S.; González Escudero, R.; Arias García, L.; Gil Martín, A.R.; Vicente Serrano, J.; Corral Fernández, E. Anisakiasis gastrointestinal manifestations: Description of 42 cases. Rev. Clin. Esp. 2005, 205, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Daschner, A.; Pascual, C.Y. Anisakis simplex: Sensitization and clinical allergy. Curr. Opin. Allergy Clin. Immunol. 2005, 5, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Audicana, M.T.; Kennedy, M.W. Anisakis simplex: From obscure infectious worm to Inducer of immune hypersensitivity. Clin. Microbiol. Rev. 2008, 21, 360–379. [Google Scholar] [CrossRef] [Green Version]
- Montoro, A.; Pretejer, M.J.; Chivato, T.; Laguna, R.; Cuellar, C. Recidivous acute urticaria caused by Anisakis simplex. Allergy 1997, 52, 985–991. [Google Scholar] [CrossRef] [PubMed]
- Audicana, L.; Audicana, M.T.; Fernandez de Corres, L.; Kennedy, M.W. Cooking and freezing may not protect against allergic reactions to ingested Anisakis simplex antigens in humans. Vet. Rec. 1997, 140, 235. [Google Scholar] [CrossRef]
- Moneo, I.; Caballero, M.L.; González-Muñoz, M.; Rodríguez-Mahillo, A.I.; Rodríguez-Perez, R.; Silva, A. Isolation of heat-resistant allergen from fish parasite Anisakis simplex. Parasitol. Res. 2005, 96, 285–289. [Google Scholar] [CrossRef]
- Caballero, M.L.; Moneo, I. Several allergens from Anisakis simplex are highly resistant to heat and pepsin treatments. Parasitol. Res. 2004, 93, 248–251. [Google Scholar] [CrossRef]
- Fitzsimmons, C.; Falcone, F.; Dunne, D. Helminth Allergens, Parasite-Specific IgE, and Its Protective Role in Human Immunity. Front. Immunol. 2014, 5, 61. [Google Scholar] [CrossRef] [Green Version]
- Arcos, S.C.; Ciordia, S.; Roberston, L.; Zapico, I.; Jiménez-Ruiz, Y.; Gonzalez-Muñoz, M.; Moneo, I.; Carballeda-Sangiao, N.; Rodriguez-Mahillo, A.; Albar, J.P.; et al. Proteomic profiling and characterization of differential allergens in the nematodes Anisakis simplex sensu stricto and A. pegreffii. Proteomics 2014, 12, 1547–1568. [Google Scholar] [CrossRef]
- Todorova, V.K.; Tankova, C.V.; Dimitrov, T.V. Trichinella spiralis- characterization of circulating and immune-complex-ssociated antigens. Parasitol. Res. 1993, 79, 86–88. [Google Scholar]
- Baeza, M.L.; Rodríguez, A.; Matheu, V.; Rubio, M.; Tornero, P.; de Barrio, M.; Herrero, T.; Santaolalla, M.; Zubeldia, J.M. Characterization of allergens secreted by Anisakis simplex parasite: Clinical relevance in comparison with somatic allergens. Clin. Exp. Allergy 2004, 34, 296–302. [Google Scholar] [CrossRef] [PubMed]
- D’Amelio, S.; Mathiopoulos, K.D.; Santos, C.P.; Pugachev, O.N.; Webb, S.C.; Picanço, M.; Paggi, L. Genetic markers in ribosomal DNA for the identification of members of the genus Anisakis (Nematoda: Ascaridoidea) defined by polymerase-chain-reaction-based restriction fragment length polymorphism. Int. J. Parasitol. 2000, 30, 223–236. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein mesurement with the folin phenol reagent. J. Biol. Chem. 1951, 173, 265–275. [Google Scholar]
- Wray, W.; Boulikas, T.; Wray, V.P.; Hancock, R. Silver staining of proteins in polyacrylamide gels. Anal. Biochem. 1982, 118, 197–203. [Google Scholar] [CrossRef]
- Biron, D.G.; Brun, C.; Levevre, T.; Lebarbenchon, C.; Loxdale, H.D.; Chevenet, F.; Brizard, J.P.; Thomas, F. The pitfalls of proteomics experiments without the correct use of bioinformatics tools. Proteomics 2006, 6, 5577–5596. [Google Scholar] [CrossRef]
- Schevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass spectrometric sequencing of proteins from silver stained polyacrylamide gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450, PubMed ID: 30395289. [Google Scholar] [CrossRef]
- Gorg, A.; Weiss, W.; Dunn, M.J. Current two-dimensional electrophoresis technology for proteomics. Proteomics 2004, 4, 3665–3685. [Google Scholar] [CrossRef]
- Barrett, J.; Brophy, P.M.; Hamilton, J.V. Analysing proteomic data. Int. J. Parasitol. 2005, 35, 543–553. [Google Scholar] [CrossRef] [PubMed]
- Cavallero, S.; Lombardo, F.; Su, X.; Salvemini, M.; Cantacessi, C.; D’Amelio, S. Tissue-specific transcriptomes of Anisakis simplex (sensu stricto) and Anisakis pegreffii reveal potential molecular mechanisms involved in pathogenicity. Parasit. Vectors. 2018, 11, 31. [Google Scholar] [CrossRef] [Green Version]
- Rawlings, N.D.; Barrett, A.J. Evolutionary families of metallopeptidases. Meth. Enzymol. 1995, 248, 183–228. [Google Scholar]
- Henrissat, B.; Callebaut, I.; Fabrega, S.; Lehn, P.; Mornon, J.P.; Davies, G. Conserved catalytic machinery and the prediction of a common fold for several families of glycosyl hydrolases. Proc. Natl. Acad. Sci. USA 1995, 92, 7090–7094. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Egerton, G.; Underwood, A.P.; Sakuda, S.; Bianco, A.E. Expression and Secretion of a Larval-specific Chitinase (Family 18 Glycosyl Hydrolase) by the Infective Stages of the Parasitic Nematode, Onchocerca volvulus. J. Biol. Chem. 2001, 276, 42557–42564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Preston, G.; Bianco, A.E. Chitinase is stored and secreted from the inner body of microfilariae and has a role in ensheathment in the parasitic nematode Brugia malayi. Mol. Biochem. Parasitol. 2008, 161, 55–62. [Google Scholar] [CrossRef]
- Gao, B.; Allen, R.; Maier, T.; McDermott, J.P.; Davis, E.L.; Baum, T.J.; Hussey, R.S. Characterization and developmental expression of a chitinase gene in Heterodera glycines. Int. J. Parasitol. 2002, 32, 1293–1300. [Google Scholar] [CrossRef]
- O’Neil, S.E.; Heinrich, T.K.; Hales, B.J.; Hazell, L.A.; Holt, D.C.; Fischer, K.; Thomas, W.R. The chitinase allergens Der p 15 and Der p 18 from Dermatophagoides pteronyssinus. Clin. Exp. Allergy 2006, 36, 831–839. [Google Scholar] [CrossRef]
- Reyes, J.L.; Terrazas, L.I. The divergent roles of alternatively activated macrophages in helminthic infections. Parasite Immunol. 2007, 29, 609–619. [Google Scholar] [CrossRef]
- Sutherland, T.E.; Maizels, R.M.; Allen, J.E. Chitinases and chitinase-like proteins: Potential therapeutic targets for the treatment of T-helper type 2 allergies. Clin. Exp. Allergy 2009, 39, 943–955. [Google Scholar] [CrossRef]
- Hewitson, J.P.; Harcus, Y.; Murray, J.; van Agtmaal, M.; Filbey, K.J.; Grainger, J.R.; Bridgett, S.; Blaxter, M.L.; Ashton, P.D.; Ashford, D.A.; et al. Proteomic analysis of secretory products from the model gastrointestinal nematode Heligmosomoides polygyrus reveals dominance of Venom Allergen-Like (VAL) proteins. J. Proteomics. 2011, 74, 1573–1594. [Google Scholar] [CrossRef] [Green Version]
- Santiago, H.C.; Bennuru, S.; Ribeiro, J.M.; Nutman, T.B. Structural Differences between Human Proteins and Aero- and Microbial Allergens Define Allergenicity. PLoS ONE 2012, 7, e40552. [Google Scholar] [CrossRef]
- Lindstedt, M.; Schiott, A.; Johnsen, C.R.; Roggen, E.; Johansson-Lindbom, B.; Borrebaeck, C.A. Individuals with occupational allergy to detergent enzymes display a differential transcriptional regulation and cellular immune response. Clin. Exp. Allergy 2005, 35, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Pan, J.; Möhrlen, F.; Schupp, M.O.; Johnsen, R.; Baillie, D.L.; Zapf, R.; Moerman, D.G.; Hutter, H. Characterization of the astacin family of metalloproteases in C. elegans. BMC Dev. Biol. 2010, 10, 14. [Google Scholar] [CrossRef] [Green Version]
- Decock, H.; Knox, D.P.; Claerebout, E.; Degraff, D.C. Partial characterisation of proteolytic enzymes in different developmental stages of Ostertagia ostertagi. J. Helmin. 1993, 67, 271–278. [Google Scholar] [CrossRef]
- Lawrence, C.E.; Pritchard, D.I. Differential secretion of acetylcholinesterase and proteases during the development of Heligmosomoides polygyrus. Int. J. Parasitol. 1993, 26, 499–508. [Google Scholar] [CrossRef]
- McKeand, J.B.; Knox, D.P.; Duncan, J.L.; Kennedy, M.W. The immunogenicity of the acetylcholinesterases of the cattle lung worm Dictyocaulus viviparus. Int. J. Parasitol. 1994, 24, 501–510. [Google Scholar] [CrossRef]
- Rhoads, M.L.; Fetterer, R.H. Developmentally regulated secretions of a cathepsin-L like protease by Haemonchus contortus. J. Parasitol. 1995, 81, 505–512. [Google Scholar] [CrossRef]
- Robertson, L.; Robertson, W.M.; Jones, J.T. Direct analysis of the secretions of the potato cyst nematode Globodera rostochiensis. Parasitology 1999, 119, 167–176. [Google Scholar] [CrossRef]
- Sakanari, J.A.; McKerrow, J.H. Identification of the secreted neutral proteases from Anisakis simplex. J. Parsitol. 1990, 76, 625–630. [Google Scholar] [CrossRef]
- Kurzik-Dumke, U.; Neubauer, M.; Debes, A. Identification of a novel Drosophila melanogaster heat-shock gene, lethal (2) denticleless [l(2)dtl], coding for an 83-kDa protein. Gene 1996, 171, 163–170. [Google Scholar] [CrossRef]
- Narberhaus, F. Alpha-crystallin-type heat shock proteins: Socializing minichaperones in the context of a multichaperone network. Microbiol. Mol. Biol. Rev. 2002, 66, 64–93. [Google Scholar] [CrossRef] [Green Version]
- Hartman, D.; Cottee, P.A.; Savin, K.W.; Bhave, M.; Presidente, P.J.; Fulton, L.; Walkiewicz, M.; Newton, S.E. Haemonchus contortus: Molecular characterisation of a small heat shock proteín. Exp. Parasitol. 2003, 104, 96–103. [Google Scholar] [CrossRef]
- Tweedie, S.; Grigg, M.E.; Ingram, L.; Selkirk, M.E. The expression of a small heat shock protein homologue is developmentally regulated in Nippostrongylus brasiliensis. Mol. Biochem. Parasitol. 1993, 61, 149–154. [Google Scholar] [CrossRef]
- Raghavan, N.; Ghosh, I.; Eisinger, W.; Pastrana, D.; Scott, A. Development regulated expression of a unique small heat shock protein in Brugia malayi. Mol. Biochem. Parasitol. 1999, 104, 233–246. [Google Scholar] [CrossRef]
- Chen, Y.; Boros, D.L. The Schistosoma mansoni egg-derived r38 peptide-induced Th1 response affects the synchronous pulmonary but not the asynchronous hepatic granuloma growth. Parasite Immunol. 2001, 23, 43–50. [Google Scholar] [CrossRef]
- Hewitson, J.P.; Harcus, Y.M.; Curwen, R.S.; Dowle, A.A.; Atmadja, A.K.; Ashton, P.D.; Wilson, A.; Maizels, R.M. The secretome of the filarial parasite, Brugia malayi: Proteomic profile of adult excretory-secretory products. Mol. Biochem. Parasitol. 2008, 160, 8–21. [Google Scholar] [CrossRef]
- González-Miguel, J.; Morchón, R.; Mellado, I.; Carretón, E.; Montoya-Alonso, J.A.; Simón, F. Excretory/secretory antigens from Dirofilaria immitis adult worms interact with the host fibrinolytic system involving the vascular endothelium. Mol. Biochem. Parasitol. 2012, 181, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Lind, S.E.; Smith, C.J. Actin Is a Noncompetitive Plasmin Inhibitor. J. Biol. Chem. 1991, 266, 5273–5278. [Google Scholar]
- Benndorf, D.; Muller, A.; Bock, K.; Manuwald, O.; Herbarth, O.; von Bergen, M. Identification of spore allergens from the indoor mould Aspergillus versicolor. Allergy 2008, 63, 454–460. [Google Scholar] [CrossRef]
- Jirapongsananuruk, O.; Jirarattanasopa, N.; Pongpruksa, S.; Vichyanond, P.; Piboonpocanun, S. Glyceraldehyde-3-phosphate dehydrogenase as a major allergen in rambutan-induced anaphylaxis. Ann. Allerg. Asthma Im. 2001, 106, 545–547. [Google Scholar] [CrossRef]
- Chalmers, I.W.; McArdle, A.J.; Coulson, R.M.; Wagner, M.A.; Schmid, R.; Hirai, H.; Hoffmann, K.F. Developmentally regulated expression, alternative splicing and distinct subgroupings in members of the Schistosoma mansoni venom allergen-like (SmVAL) gene family. BMC Genom. 2008, 9, 89. [Google Scholar] [CrossRef] [Green Version]
- Zhan, B.; Liu, Y.; Badamchian, M.; Williamson, A.; Feng, J.; Loukas, A.; Hawdon, J.M.; Hotez, P.J. Molecular characterisation of the Ancylostoma-secreted protein family from the adult stage of Ancylostoma caninum. Int. J. Parasitol. 2003, 33, 897–907. [Google Scholar] [CrossRef]
- Murray, J.; Gregory, W.F.; Gomez-Escobar, N.; Atmadja, A.K.; Maizels, R.M. Expression and immune recognition of Brugia malayi VAL-1, a homologue of vespid venom allergens and Ancylostoma secreted proteins. Mol. Biochem. Parasitol. 2001, 118, 89–96. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, A.J.; Tawe, W.; Leon, O.; Cao, L.; Liu, J.; Oksov, Y. Ov-ASP-1, the Onchocerca volvulus homologue of the activation associated secreted protein family is immunostimulatory and can induce protective anti-larval immunity. Parasite Immunol. 2004, 26, 53–62. [Google Scholar] [CrossRef]
- Klein, R.D.; Winterrowd, C.A.; Hatzenbuhler, N.T.; Shea, M.H.; Favreau, M.A.; Nulf, S.C.; Geary, T.G. Cloning of a cDNA encoding phosphoenolpyruvate carboxykinase from Haemonchus contortus. Mol. Biochem. Parasitol. 1992, 50, 285–294. [Google Scholar] [CrossRef]
- Geary, T.G.; Winterrowd, C.A.; Alexander-Bowman, S.J.; Favreau, M.A.; Nulf, S.C.; Klein, R.D. Ascaris suum: Cloning of a cDNA Encoding Phosphoenolpyruvate Carboxykinase. Exp. Parasitol. 1993, 77, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Seidler, N.W. GAPDH, as a virulence factor. Adv. Exp. Med. Biol. 2013, 985, 149–178. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.P.; Mao, X.Y.; Ren, F.Z.; Che, H.L. Attenuating effect of casein glycomacropeptide on proliferation, differentiation, and lipid accumulation of in vitro Sprague-Dawley rat preadipocytes. J. Dairy Sci. 2011, 676–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, L.; Bedding, R.A. Characteristics of protectant synthesis of infective juveniles of Steinernema carpocapsae and importance of glycerol as a protectant for survival of the nematodes during osmotic dehydration. Comp. Biochem. Physiol. 2002, 131, 757–765. [Google Scholar] [CrossRef]
- Rothstein, M. Nematode bio chemistry. X. Excretion of glycerol by free living nematodes. Comp. Biochem. Physiol. 1969, 30, 641–648. [Google Scholar] [CrossRef]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef]
- Carr, P.; Ollis, D. α/β Hydrolase Fold: An Update. Protein Peptide Lett. 2009, 16, 1137–1148. [Google Scholar] [CrossRef]
- Reis Montella, I.; Schama, R.; Valle, D. The classification of sterases: An important gene family involved in insecticide resistance. A review. Mem. Inst. Oswaldo Cruz. 2012, 107. [Google Scholar] [CrossRef] [Green Version]
- Aranda, J.; Cerqueira, N.M.F.S.A.; Fernandes, P.A.; Roca, M.; Tuñon, I.; Ramos, M.J. The Catalytic Mechanism of Carboxylesterases. A Computational Study. Biochemistry 2014, 53, 5820–5829. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.L. Why do some nematode parasites of the alimentary tract secrete acetylcholinesterase. Int. J. Parasitol. 1996, 26, 499–508. [Google Scholar] [CrossRef]
- Podolska, M.; Nadolna, K.; Wąs, A.; Gosz, E.; Szostakowska, B. Acetylcholinesterase secretion by third-stage larvae of Anisakis simplex (Nematoda: Anisakidae) from Baltic herring. Bull. Eur. Ass. Fish Pathol. 2012, 32, 225–232. [Google Scholar]
- Podolska, M.; Nadolna, K. Acetylcholinesterase secreted by Anisakis simplex larvae (Nematoda: Anisakidae) parasitizing herring, Clupea harengus: An inverse relationship of enzyme activity in the host-parasite system. Parasitol. Res. 2014, 13, 2231–2238. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, E.; Romaris, F.; Lorenzo, S.; Moreno, J.; Bonay, P.; Ubeira, F.M.; Garate, T.A. A recombinant enolase from Anisakis simplex is differentially recognized in natural human and mouse experimental infections. Med. Microbiol Immunol. 2006, 195, 1–10. [Google Scholar] [CrossRef]
- Liu, J.K.; Shih, N.Y. The role of enolase in tissue invasion and metastasis of pathogens and tumor cells. J. Cancer Mol. 2007, 3, 45–48. [Google Scholar]
- Liu, H.; Zeng, H.; Yao, Q.; Yuan, J.; Zhang, Y.; Qiu, D.; Yang, X.; Yang, H.; Liu, Z. Steinernema glaseri surface enolase: Molecular cloning, biological characterization, and role in host immune suppression. Mol. Biochem. Parasitol. 2012, 185, 89–98. [Google Scholar] [CrossRef]
- Llorens, C.; Arcos, S.C.; Robertson, L.; Ramos, R.; Futami, R.; Soriano, B.; Ciordia, S.; Careche, M.; González-Muñoz, M.; Jiménez-Ruiz, Y.; et al. Functional insights into the infective larval stage of Anisakis simplex s.s., Anisakis pegreffii and their hybrids based on gene expression patterns. BMC Genom. 2018. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Protein | UniProt Accession | Taxonomy | Matched Peptides | (MS/MS) | Score | Gene | |
|---|---|---|---|---|---|---|---|
| 1-D bands (Figure 1 and Figure 2) | |||||||
| A | Peptidase_M13_N domain-containing protein | A0A0M3KHN4 | Anisakis simplex | 10 | R.ILTNYIVWR.Y R.YDDILQDFLR.V K.WAQNYWFR.K | 133/175 | ASIM_LOCUS19882 |
| B | Uncharacterized protein | A0A0M3IZF7 | Anisakis simplex | 21/22 | R.PFMGYNFGR.Y R.MGAAVLHELTR.T R.RPEGVPLVFESR.D R.LYAINPEAGFFGVAPGTSHK.T | 205/238 | ASIM_LOCUS540 |
| C | unidentified | 4 | R.SFLGVPFAEPPIGENR.F | 51/56 | N/A | ||
| D | SCP domain-containing protein predicted | A0A0M3JRE8 | Anisakis simplex | 12/13 | R.RAQEYAER.C R.NMYYLDYDCDLER.R R.STNLCIAPGYTAPEPNPR.P | 193/255 | ASIM_LOCUS10032 |
| E | Uncharacterized protein | A0A0M3K6E2 | Anisakis simplex | 20 | R.SKEEADDCFYR.G K.EFVALNEDNQKR.C K.TADGFYTDCYFGR.G K.CWPYSYEAFVIAAR.Y R.FGTSSPNSVYTPEENTR.R R.FGTSSPNSVYTPEENTRR.D R.DVAAFFAHAIQETGANDASVYTGR.S | 469/530 | ASIM_LOCUS15940 |
| F | Lipase_3 domain-containing protein | A0A0M3K4F3 | Anisakis simplex | 10 | K.HTELVPYSFR.V K.IIADHASYFDR.V K.NVLPSDEKWEVVER.I R.GTTTSSQLFLQGAGAISGR.A | 120/134 | ASIM_LOCUS15251 |
| Actin 3 | A0A0B4SVN7 | Anisakis simplex | 9/10 | K.QEYDESGPSIVHR.K K.SYELPDGQVITIGNER.F | 113/119 | N/A | |
| G | Lipase_3 domain-containing protein | A0A0M3K4F3 | Anisakis simplex | 7/6 | K.HTELVPYSFR.V | 71/68 | ASIM_LOCUS15251 |
| G | Uncharacterized protein | A0A0M3K6E2 | Anisakis simplex | 11 | K.TADGFYTDCYFGR.G | 80 | ASIM_LOCUS15940 |
| H | Glyceraldehyde-3-phosphate dehydrogenase | A0A097IYH5 | Anisakis simplex | 4 | R.VPTPDVSVVDLTCR.L K.LISWYDNEFGYSCR.V | 96 | GPDH |
| H | Hemoglobin | A0A1W7HP35 | Anisakis simplex | 10 | --- | 58 | N/A |
| H | Uncharacterized protein | A0A0M3JQQ1 | Anisakis simplex | 2 | R.VLTDAIYLISHIDGTTR.M | 147 | ASIM_LOCUS9737 (very sort seq) |
| H | Metalloendopeptidase | A0A0M3K299 | Anisakis simplex | 9/7 | R.VLFENINPPMR.C K.SSNYFLTDEDFER.A | 142/94 | ASIM_LOCUS14439 |
| I | Metalloendopeptidase | A0A0M3K299 | Anisakis simplex | 10/9 | R.VLFENINPPMR.C K.SSNYFLTDEDFER.A | 180/156 | ASIM_LOCUS14439 |
| I | Metalloendopeptidase | A0A3G5BC99 | Anisakis simplex | 10/9 | R.TGFSLNDVR.K | 102/94 | nas-13 |
| J | Metalloendopeptidase | A0A0M3K299 | Anisakis simplex | 8 | R.VLFENINPPMR.C K.SSNYFLTDEDFER.A | 92/134 | ASIM_LOCUS14439 |
| K | Metalloendopeptidase | A0A0M3K299 | Anisakis simplex | 9 | R.DPNALWPK.G R.VLFENINPPMR.C K.SSNYFLTDEDFER.A | 139/126 | ASIM_LOCUS14439 |
| L | Metalloendopeptidase | A0A0M3K299 | Anisakis simplex | 10 | R.DPNALWPK.G R.ADRDDYVR.V R.VLFENINPPMR.C | 138 | ASIM_LOCUS14439 |
| M | Protein lethal(2) essential for life (inferred by orthology to a D. melanogaster protein) | A0A0M3JX08 | Anisakis simplex | 8 | R.SIPIQAAPR.Q K.QNQQLPAAR.S R.YAIVPPTFER.A | 96 | ASIM_LOCUS12322 |
| M | SCP domain-containing protein | A0A0M3K1U4 | Anisakis simplex | 5 | K.QVNVVCEYR.N | 71 | ASIM_LOCUS14262 |
| 2-D spots (Figure 3) | |||||||
| 16 | Peptidase_M13_N domain-containing protein | A0A0M3JI80 | Anisakis simplex | 7 | K.YDLTNLLIHTSLTR.A | 78 | ASIM_LOCUS7120 |
| 16 | Peptidase_M13_N domain-containing protein | A0A0M3KHN4 | Anisakis simplex | 4 | R.ILTNYIVWR.Y | 70 | ASIM_LOCUS19882 |
| 29 | Uncharacterized protein | A0A0M3K8A1 | Anisakis simplex | 21 | K.SASIVNEQR.I R.FVCSLGVDPGIK.L K.LAAEIGALQHDER.Y | 214 | ASIM_LOCUS16599 |
| 31 | CO esterase domain-containing protein | A0A0M3JYK8 | Anisakis simplex | 6 | R.ADVFLGVPYAQPPVGALR.F | 101 | ASIM_LOCUS12965 |
| 35 | Enolase | Q8MU59 | Anisakis simplex | 8 | R.AAVPSGASTGVHEALELR.D K.STLNIQLVGDDLTVTNR.E | 101 | eno |
| 36 | Enolase | Q8MU59 | Anisakis cimplex | 16 | R.AAVPSGASTGVHEALELR.D K.STLNIQLVGDDLTVTNR.E R.YGLDATAVGDEGGFAPNIQDNR.E | 276 | eno |
| 37 | Enolase | Q8MU59 | Anisakis simplex | 7 | R.AAVPSGASTGVHEALELR.D | 66 | eno |
| 46 | SCP domain-containing protein | A0A0M3JRE8 | Anisakis simplex | 6 | R.RAQEYAER.C R.STNLCIAPGYTAPEPNPR.P | 59 | ASIM_LOCUS10032 |
| 50 | SCP domain-containing protein | A0A0M3JRE8 | Anisakis simplex | 7 | R.RAQEYAER.C R.STNLCIAPGYTAPEPNPR.P | 84 | ASIM_LOCUS10032 |
| 51 | SCP domain-containing protein | A0A0M3JRE8 | Anisakis simplex | 10 | R.RAQEYAER.C R.NMYYLDYDCDLER.R R.STNLCIAPGYTAPEPNPR.P | 159 | ASIM_LOCUS10032 |
| 52 | SCP domain-containing protein | A0A0M3JRE8 | Anisakis simplex | 9 | R.RAQEYAER.C R.NMYYLDYDCDLER.R R.STNLCIAPGYTAPEPNPR.P | 148 | ASIM_LOCUS10032 |
| 54 | SCP domain-containing protein | A0A0M3JRE8 | Anisakis simplex | 9 | R.RAQEYAER.C R.NMYYLDYDCDLER.R R.STNLCIAPGYTAPEPNPR.P | 136 | ASIM_LOCUS10032 |
| 57 | Metalloendopeptidase | A0A3G5BC99 | Anisakis simplex | 9 | R.TGFSLNDVR.K | 97 | nas-13 |
| 58 | Metalloendopeptidase | A0A3G5BC99 | Anisakis simplex | 6 | R.TGFSLNDVR.K | 75 | nas-13 |
| 59 | Metalloendopeptidase | A0A3G5BC99 | Anisakis simplex | 15 | R.TGFSLNDVR.K R.TGFSLNDVRK.I K.GVVIHELMHALGIQHEQSR.T | 190 | nas-13 |
| 60 | Metalloendopeptidase | A0A3G5BC99 | Anisakis simplex | 10 | R.TGFSLNDVR.K R.TGFSLNDVRK.I K.NGKPTIVALEPNR.N | 103 | nas-13 |
| 61 | Metalloendopeptidase | A0A3G5BC99 | Anisakis simplex | 7 | R.TGFSLNDVR.K | 70 | nas-13 |
| 63 | Glycerol-3-phosphate dehydrogenase | A0A0M3K2U5 | Anisakis simplex | 10 | MVSLRNAIVGFTR.A | 47 | ASIM_LOCUS14675 |
| 64 | Metalloendopeptidase | A0A0M3K299 | Anisakis simplex | 8 | R.ADRDDYVR.V R.VLFENINPPMR.C R.VLFENINPPMR.C K.SSNYFLTDEDFER.A | 96 | ASIM_LOCUS14439 |
| 67 | Metalloendopeptidase | A0A0M3K299 | Anisakis simplex | 8 | R.VLFENINPPMR.C R.VLFENINPPMR.C K.SSNYFLTDEDFER.A | 139 | ASIM_LOCUS14439 |
| 68 | Metalloendopeptidase | A0A0M3K299 | Anisakis simplex | 10 | R.VLFENINPPMR.C R.VLFENINPPMR.C K.SSNYFLTDEDFER.A K.SSNYFLTDEDFERAR.S | 169 | ASIM_LOCUS14439 |
| 69 | Metalloendopeptidase | A0A0M3K299 | Anisakis simplex | 7 | R.ADRDDYVR.V K.SSNYFLTDEDFER.A | 82 | ASIM_LOCUS14439 |
| 70 | Metalloendopeptidase | A0A0M3K299 | Anisakis simplex | 8 | R.ADRDDYVR.V R.VLFENINPPMR.C R.VLFENINPPMR.C | 90 | ASIM_LOCUS14439 |
| 73 | Metalloendopeptidase | A0A0M3K299 | Anisakis simplex | 4 | R.ADRDDYVR.V K.SSNYFLTDEDFER.A | 84 | ASIM_LOCUS14439 |
| 79 | SCP domain-containing protein | A0A0M3K1U4 | Anisakis simplex | 9 | K.QVNVVCEYR.N K.YGVTASEAWWSELKR.V R.HAGENIFASSTTGSLGDLGK.Y | 247 | ASIM_LOCUS14262 |
| 80 | SCP domain-containing protein | A0A0M3K1U4 | Anisakis simplex | 6 | K.QVNVVCEYR.N | 75 | ASIM_LOCUS14262 |
| 81 | SCP domain-containing protein | A0A0M3K1U4 | Anisakis simplex | 4 | K.QVNVVCEYR.N | 53 | ASIM_LOCUS14262 |
| 82 | Protein lethal(2)essential for life | A0A0M3JX08 | Anisakis simplex | 9 | R.SIPIQAAPR.Q R.YAIVPPTFER.A | 88 | ASIM_LOCUS12322 |
| Gene | Protein | UniProt Accession | 1-D band | 2-D spot | Molecular function | Biological Process |
|---|---|---|---|---|---|---|
| ASIM_LOCUS19882 | Peptidase_M13_N domain-containing protein | A0A0M3KHN4 | band A | 16 | Catalysis of the hydrolysis of internal, alpha-peptide bonds in a polypeptide chain | Metalloendopeptidase activity |
| ASIM_LOCUS7120 | Peptidase_M13_N domain-containing protein | A0A0M3JI80 | 16 | |||
| ASIM_LOCUS540 | Uncharacterized protein | A0A0M3IZF7 | band B | phosphoenolpyruvate carboxykinase (GTP) activity | gluconeogenesis | |
| ASIM_LOCUS10032 | SCP domain-containing protein (*) | A0A0M3JRE8 | band D | 46, 50, 51, 52, 54 | Unclear function. Venom allergens of some insects and Scolopendra | Belongs to the CRISP family (cysteine-rich secretory proteins) |
| ASIM_LOCUS14262 | SCP domain-containing protein (*) | A0A0M3K1U4 | band M | 79, 80, 81 | ||
| ASIM_LOCUS15940 | Uncharacterized protein (*) | A0A0M3K6E2 | bands E, G | chitinase activity | cell wall macromolecule catabolic process | |
| ASIM_LOCUS15251 | Lipase_3 domain-containing protein (*) | A0A0M3K4F3 | bands F, G | hydrolase activity | lipid metabolic process | |
| N/A | Actin 3 | A0A0B4SVN7 | band F | ATP binding | Cytoskeleton | |
| ASIM_LOCUS14439 | Metalloendopeptidase | A0A0M3K299 | bands H, I, J, K, L | 64, 67,68, 69, 70, 73 | Metalloprotease | Metalloendopeptidase activity |
| nas-13 | Metalloendopeptidase | A0A3G5BC99 | band I | 57, 58, 59, 60, 61 | ||
| GPDH | Glyceraldehyde-3-phosphate dehydrogenase (*) | A0A097IYH5 | band H | Catalytic activity (Oxidoreductase) | glucose metabolic process (Glycolisis) | |
| ASIM_LOCUS14675 | Glycerol-3-phosphate dehydrogenase | A0A0M3K2U5 | 63 | Catalytic activity (calcium ion binding) | Oxydation-reduction process | |
| ASIM_LOCUS12322 | Protein lethal(2)essential for life | A0A0M3JX08 | band M | 82 | prevent the precipitation of denatured proteins and to increase cellular tolerance to stress | Alpha crystallin/Small heat shock protein, animal type |
| ASIM_LOCUS16599 | Uncharacterized protein | A0A0M3K8A1 | 29 | Unknown function | From a family of proteins over 500 amino acids in Caenorhabditis elegans and several bacteria | |
| ASIM_LOCUS12965 | CO esterase domain-containing protein | A0A0M3JYK8 | 31 | Unknown function | ||
| eno | Enolase (allergen Ani s Enolase) (*) | Q8MU59 | 35, 36, 37 | magnesium ion binding Phosphopyruvate hydratase activity | Glycolytic process |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robertson, L.; C. Arcos, S.; Ciordia, S.; Carballeda-Sanguiao, N.; Mena, M.d.C.; Sánchez-Alonso, I.; Gonzalez-Muñoz, M.; Careche, M.; Navas, A. Immunoreactive Proteins in the Esophageal Gland Cells of Anisakis Simplex Sensu Stricto Detected by MALDI-TOF/TOF Analysis. Genes 2020, 11, 683. https://doi.org/10.3390/genes11060683
Robertson L, C. Arcos S, Ciordia S, Carballeda-Sanguiao N, Mena MdC, Sánchez-Alonso I, Gonzalez-Muñoz M, Careche M, Navas A. Immunoreactive Proteins in the Esophageal Gland Cells of Anisakis Simplex Sensu Stricto Detected by MALDI-TOF/TOF Analysis. Genes. 2020; 11(6):683. https://doi.org/10.3390/genes11060683
Chicago/Turabian StyleRobertson, Lee, Susana C. Arcos, Sergio Ciordia, Noelia Carballeda-Sanguiao, María del Carmen Mena, Isabel Sánchez-Alonso, Miguel Gonzalez-Muñoz, Mercedes Careche, and Alfonso Navas. 2020. "Immunoreactive Proteins in the Esophageal Gland Cells of Anisakis Simplex Sensu Stricto Detected by MALDI-TOF/TOF Analysis" Genes 11, no. 6: 683. https://doi.org/10.3390/genes11060683
APA StyleRobertson, L., C. Arcos, S., Ciordia, S., Carballeda-Sanguiao, N., Mena, M. d. C., Sánchez-Alonso, I., Gonzalez-Muñoz, M., Careche, M., & Navas, A. (2020). Immunoreactive Proteins in the Esophageal Gland Cells of Anisakis Simplex Sensu Stricto Detected by MALDI-TOF/TOF Analysis. Genes, 11(6), 683. https://doi.org/10.3390/genes11060683