Aspects of Multicellularity in Saccharomyces cerevisiae Yeast: A Review of Evolutionary and Physiological Mechanisms


{kind=link}
Abstract
:1. Introduction
2. Mechanisms and Methods to Study Saccharomyces cerevisiae Adhesion
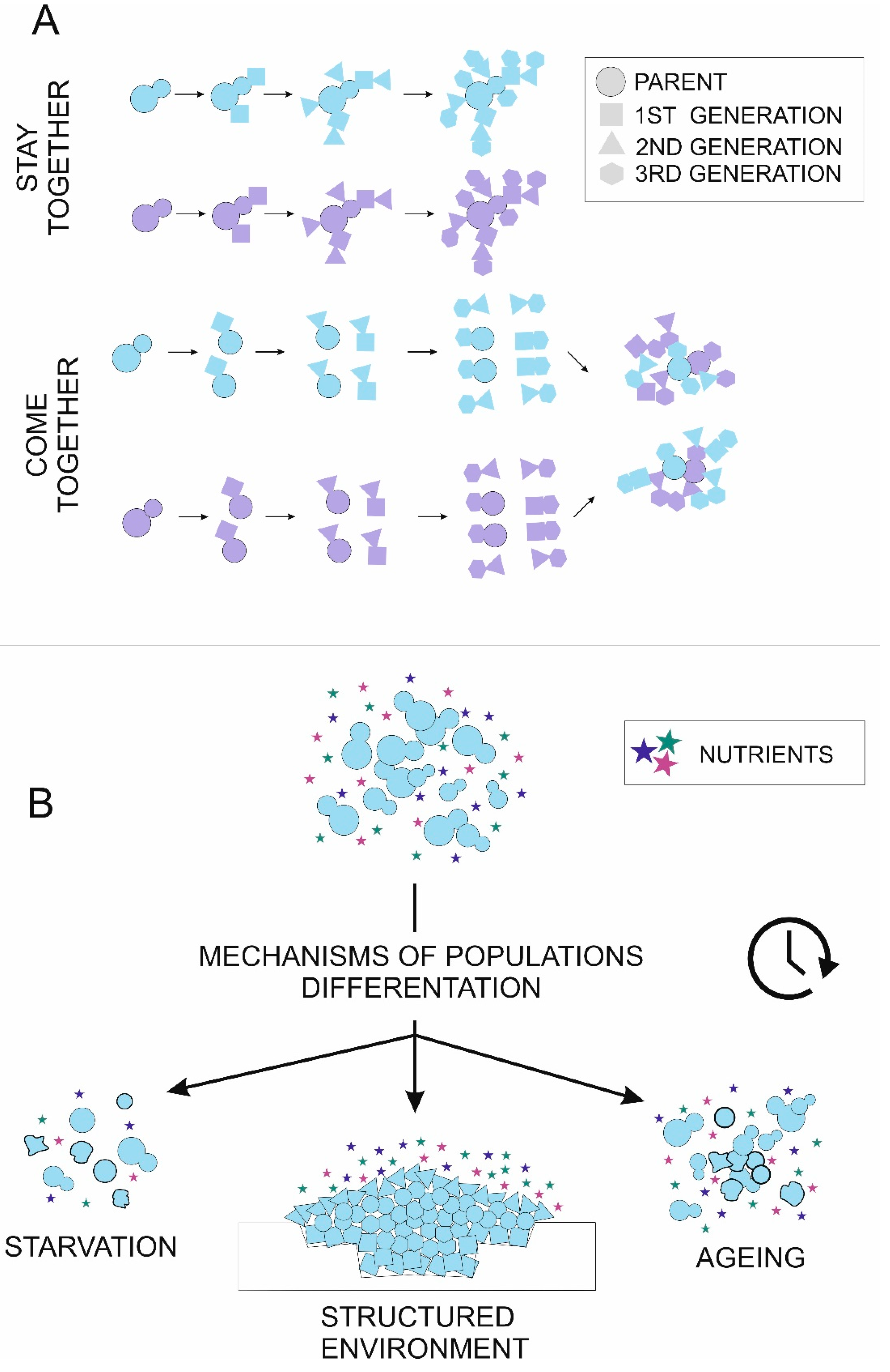
3. Mechanisms and Methods to Study Saccharomyces cerevisiae Cell Differentiation
3.1. Structured Environment
3.2. Starvation
3.3. Aging
4. Evolutionary Advantages of Yeast Multicellularity and Heterogeneity
5. Conclusions and Future Directions
Funding
Conflicts of Interest
Glossary
| Stay together strategy: | One of the possible types of multicellular group formation, where cells reproducing vegetatively by budding form groups by cell-cell adhesion or incomplete cell wall separation, where the daughter cell stays attached to its mother cell. |
| Come together strategy: | One of the possible types of multicellular group formation, where individual cells adhere to each other by cell wall proteins. |
| Colony, non-adhesive colony: | Structures formed on solid substrates exposed to air. Cells that do not produce adhesins are sessile and will develop non-adhesive colonies. This is the typical growth form of many laboratory strains on solid agar media [11]. |
| Biofilm: | Multicellular structure (adhesive colony) on a semisolid medium. Typically, cells in biofilms adhere to each other and to the foreign surface [11]. |
| Mat: | |
| Stalks: | Stalk structures growing vertically from small holes in the agar. Saccharomyces cerevisiae stalks are 5–30-mm long differentiated structures composed of two distinct layers, identified by ultrastructural analysis, of stalk thin sections. The central core, which is composed of yeast cells and spores, is surrounded by surface layers of highly vacuolized cells with thick cell walls. These cells are dying (or already dead) and seem to protect the structure against drying and other environmental effects, forming a skin-like structure [37]. |
| Flocs: | Clumps of cells created by cell-to-cell adhesion that may sediment to the bottom. Cells can stick to each other due to cell wall proteins—flocculins [11]. |
| Flors: | Specific types of interfacial biofilm on air–liquid self-adhesion groups that can float on the liquid surface [11]. |
| Division of labour: | Three conditions are needed to identify a division of labour: (a) phenotypic variation, reflecting different tasks undertaken by individuals within a population; (b) cooperation between individuals performing different tasks and (c) adaptation—that the behaviour maximizes the inclusive fitness of all individuals involved [33]. |
| Public goods: | Resources produced by cooperating organisms, which are available in the environment and can be used by other organisms in the population. |
| Cooperation: | Interaction in which organisms perform a costly task together for the common benefit (for example, production of a public good), which can be used by all population members. |
| Homologous recombination: | Method of genetic modification in which chromosomal genes are swapped with genes on plasmid (which can be designed by the researcher), so that a gene of interest is transferred to the chromosome. |
References
- Szathmary, E.; Maynard-Smith, J.M. The Major Evolutionary Transitions. Nature 1995, 374, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Grosberg, R.K.; Strathmann, R.R. The evolution of multicellularity: A minor major transition? Annu. Rev. Ecol. Evol. Syst. 2007, 38, 621–654. [Google Scholar] [CrossRef] [Green Version]
- Nagy, L.G.; Kovács, G.M.; Krizsán, K. Complex multicellularity in fungi: Evolutionary convergence, single origin, or both? Biol. Rev. 2018, 93, 1778–1794. [Google Scholar] [CrossRef] [PubMed]
- Knoll, A.H. The multiple origins of complex multicellularity. Annu. Rev. Earth Planet Sci. 2011, 39, 217–239. [Google Scholar] [CrossRef] [Green Version]
- Honigberg, S.M. Cell signals, cell contacts, and the organization of yeast communities. Eukaryot Cell 2011, 10, 466–473. [Google Scholar] [CrossRef] [Green Version]
- Ispolatov, I.; Ackermann, M.; Doebeli, M. Division of labour and the evolution of multicellularity. Proc. R. Soc. Lond. B Biol. Sci. 2012, 279, 1768–1776. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, J.A.; Doebeli, M. A simple and general explanation for the evolution of altruism. Soc. Lond. B Biol. Sci. 2009, 276, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Momeni, B.; Waite, A.J.; Shou, W. Spatial self-organization favors heterotypic cooperation over cheating. Elife 2013, 2, e00960. [Google Scholar] [CrossRef]
- Goldsby, H.J.; Dornhaus, A.; Kerr, B.; Ofria, C. Task-switching costs promote the evolution of division of labor and shifts in individuality. Proc. Natl. Acad. Sci. USA 2012, 109, 13686–13691. [Google Scholar] [CrossRef] [Green Version]
- Schiessl, K.T.; Ross-Gillespie, A.; Cornforth, D.M.; Weigert, M.; Bigosch, C.; Brown, S.P.; Ackermann, M.; Kümmerli, R. Individual-versus group-optimality in the production of secreted bacterial compounds. Evolution 2019, 73, 675–688. [Google Scholar] [CrossRef] [Green Version]
- Bruckner, S.; Mosch, H.U. Choosing the right lifestyle: Adhesion and development in Saccharomyces cerevisiae. FEMS Microbiol. Rev. 2012, 36, 25–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yvert, G.; Brem, R.B.; Whittle, J.; Akey, J.M.; Foss, E.; Smith, E.N.; Mackelprang, R.; Kruglyak, L. Trans-acting regulatory variation in Saccharomyces cerevisiae and the role of transcription factors. Nat. Genet. 2003, 35, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Wloch-Salamon, D.M.; Plech, M.; Majewska, J. Generation of stable, non-aggregating Saccharomyces cerevisiae wild isolates. Acta Biochim. Pol. 2013, 60, 657–660. [Google Scholar] [CrossRef] [Green Version]
- Koschwanez, J.H.; Foster, K.R.; Murray, A.W. Sucrose utilization in budding yeast as a model for the origin of undifferentiated multicellularity. PLoS Biol. 2011, 9, e1001122. [Google Scholar] [CrossRef]
- Sandberg, T.E.; Salazar, M.J.; Weng, L.L.; Palsson, B.O.; Feist, A.M. The emergence of adaptive laboratory evolution as an efficient tool for biological discovery and industrial biotechnology. Metab. Eng. 2019. [Google Scholar] [CrossRef]
- Kawecki, T.J.; Lenski, R.E.; Ebert, D.; Hollis, B.; Olivieri, I.; Whitlock, M.C. Experimental evolution. Trends Ecol. Evol. 2012, 27, 547–560. [Google Scholar] [CrossRef] [Green Version]
- Ratcliff, W.C.; Denison, R.F.; Borrello, M.; Travisano, M. Experimental evolution of multicellularity. Proc. Natl. Acad. Sci. USA 2012, 109, 1595–1600. [Google Scholar] [CrossRef] [Green Version]
- Ratcliff, W.C.; Fankhauser, J.D.; Rogers, D.W.; Greig, D.; Travisano, M. Origins of multicellular evolvability in snowflake yeast. Nat. Commun. 2015, 6, 6102. [Google Scholar] [CrossRef] [Green Version]
- Oud, B.; Guadalupe-Medina, V.; Nijkamp, J.F.; de Ridder, D.; Pronk, J.T.; van Maris, A.J.A.; Daran, J.M. Genome duplication and mutations in ACE2 cause multicellular, fast-sedimenting phenotypes in evolved Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2013, 110, E4223–E4231. [Google Scholar] [CrossRef] [Green Version]
- Kayacan, Y.; Van Mieghem, T.; Delvaux, F.; Delvaux, F.R.; Willaert, R. Adaptive Evolution of Industrial Brewer’s Yeast Strains towards a Snowflake Phenotype. Fermentation 2020, 6, 20. [Google Scholar] [CrossRef] [Green Version]
- Kuzdzal-Fick, J.; Chen, L.; Balazsi, G. Evolutionary trade-offs between unicellularity and multicellularity in budding yeast. BioRxiv 2018, 347609. [Google Scholar]
- Teunissen, A.W.R.H.; Steensma, H.Y. The dominant flocculation genes of Saccharomyces cerevisiae constitute a new subtelomeric gene family. Yeast 1995, 11, 1001–1013. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Styles, C.A.; Feng, Q.; Fink, G.R. A Saccharomyces gene family involved in invasive growth, cell–cell adhesion, and mating. Proc. Natl. Acad. Sci. USA 2000, 97, 12158–12163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, R.M.; Regenberg, B. Multicellular group formation in Saccharomyces cerevisiae. Proc R Soc. Lond. B 2019, 286, 20191098. [Google Scholar] [CrossRef] [Green Version]
- Brückner, S.; Schubert, R.; Kraushaar, T.; Hartmann, R.; Hoffmann, D.; Jelli, E.; Drescher, K.; Müller, D.J.; Essen, L.O.; Mösch, H.-U. Kin discrimination in social yeast is mediated by cell surface receptors of the Flo11 adhesin family. Elife 2020, 9, e55587. [Google Scholar] [CrossRef]
- Vachova, L.; Stovicek, V.; Hlavacek, O.; Chernyavskiy, O.; Stepanek, L.; Kubinova, L.; Palkova, Z. Flo11p, drug efflux pumps, and the extracellular matrix cooperate to form biofilm yeast colonies. J. Cell Biol. 2011, 194, 679–687. [Google Scholar] [CrossRef] [Green Version]
- Torbensen, R.; Moller, H.D.; Gresham, D.; Alizadeh, S.; Ochmann, D.; Boles, E.; Regenberg, B. Amino Acid Transporter Genes Are Essential for FLO11-Dependent and FLO11-Independent Biofilm Formation and Invasive Growth in Saccharomyces cerevisiae. PLoS ONE 2012, 7, e41272. [Google Scholar] [CrossRef] [Green Version]
- Kraushaar, T.; Brückner, S.; Veelders, M.; Rhinow, D.; Schreiner, F.; Birke, R.; Pagenstecher, A.; Mösch, H.U.; Essen, L.O. Interactions by the fungal Flo11 adhesin depend on a fibronectin type III-like adhesin domain girdled by aromatic bands. Structure 2015, 23, 1005–1017. [Google Scholar] [CrossRef] [Green Version]
- Rossouw, D.; Meiring, S.P.; Bauer, F.F. Modifying Saccharomyces cerevisiae adhesion properties regulates yeast ecosystem dynamics. mSphere 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Hope, E.A.; Amorosi, C.J.; Miller, A.W.; Dang, K.; Heil, C.S.; Dunham, M.J. Experimental evolution reveals favored adaptive routes to cell aggregation in yeast. Genetics 2017, 206, 1153–1167. [Google Scholar] [CrossRef] [Green Version]
- Smukalla, S.; Caldara, M.; Pochet, N.; Beauvais, A.; Guadagnini, S.; Yan, C.; Vinces, M.D.; Jansen, A.; Prevost, M.C.; Latge, J.P.; et al. FLO1 is a variable green beard gene that drives biofilm-like cooperation in budding yeast. Cell 2008, 135, 726–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piccirillo, S.; Kapros, T.; Honigberg, S.M. Phenotypic plasticity within yeast colonies: Differential partitioning of cell fates. Curr. Genet. 2016, 62, 467–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wloch-Salamon, D.M.; Fisher, R.M.; Regenberg, B. Division of labour in the yeast, Saccharomyces cerevisiae. Yeast 2017. [Google Scholar] [CrossRef] [Green Version]
- Takhaveev, V.; Heinemann, M. Metabolic heterogeneity in clonal microbial populations. Curr. Opin. Microbiol. 2018, 45, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M.; Stecher, B.; Freed, N.E.; Songhet, P.; Hardt, W.-D.; Doebeli, M. Self-destructive cooperation mediated by phenotypic noise. Nature 2008, 454, 987–990. [Google Scholar] [CrossRef]
- Schreiber, F.; Ackermann, M. Environmental drivers of metabolic heterogeneity in clonal microbial populations. Curr. Opin. Biotechnol. 2020, 62, 202–211. [Google Scholar] [CrossRef]
- Váchová, L.; Palková, Z. How structured yeast multicellular communities live, age and die? FEMS Yeast Res. 2018, 18, foy033. [Google Scholar] [CrossRef]
- Nagarajan, S.; Kruckeberg, A.L.; Schmidt, K.H.; Kroll, E.; Hamilton, M.; McInnerney, K.; Summers, R.; Taylor, T.; Rosenzweig, F. Uncoupling reproduction from metabolism extends chronological lifespan in yeast. Proc. Natl. Acad. Sci. USA 2014, 111, E1538–E1547. [Google Scholar] [CrossRef] [Green Version]
- Marinkovic, Z.S.; Vulin, C.; Acman, M.; Song, X.; Di Meglio, J.M.; Lindner, A.B.; Hersen, P. A microfluidic device for inferring metabolic landscapes in yeast monolayer colonies. Elife 2019, 8. [Google Scholar] [CrossRef]
- Pfeiffer, T.; Morley, A. An evolutionary perspective on the Crabtree effect. Front. Mol. Biosci. 2014, 1, 17. [Google Scholar] [CrossRef]
- Hagman, A.; Piškur, J. A study on the fundamental mechanism and the evolutionary driving forces behind aerobic fermentation in yeast. PLoS ONE 2015, 10, e0116942. [Google Scholar] [CrossRef] [PubMed]
- Regenberg, B.; Hanghøj, K.E.; Andersen, K.S.; Boomsma, J.J. Clonal yeast biofilms can reap competitive advantages through cell differentiation without being obligatorily multicellular. Proc. R. Soc. B 2016, 283, 20161303. [Google Scholar] [CrossRef] [PubMed]
- White, M.G.; Piccirillo, S.; Dusevich, V.; Law, D.J.; Kapros, T.; Honigberg, S.M. Flo11p adhesin required for meiotic differentiation in Saccharomyces cerevisiae minicolonies grown on plastic surfaces. FEMS Yeast Res. 2011, 11, 223–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faria-Oliveira, F.; Carvalho, J.; Belmiro, C.L.R.; Martinez-Gomariz, M.; Hernaez, M.L.; Pavão, M.; Gil, C.; Lucas, C.; Ferreira, C. Methodologies to generate, extract, purify and fractionate yeast ECM for analytical use in proteomics and glycomics. BMC Microbiol. 2014, 14, 244. [Google Scholar] [CrossRef] [Green Version]
- Stovicek, V.; Vachova, L.; Palkova, Z. Yeast biofilm colony as an orchestrated multicellular organism. Commun Integr. Biol. 2012, 5, 203–205. [Google Scholar] [CrossRef] [PubMed]
- Čáp, M.; Štěpánek, L.; Harant, K.; Váchová, L.; Palková, Z. Cell differentiation within a yeast colony: Metabolic and regulatory parallels with a tumor-affected organism. Mol. Cell 2012, 46, 436–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paczia, N.; Nilgen, A.; Lehmann, T.; Gätgens, J.; Wiechert, W.; Noack, S. Extensive exometabolome analysis reveals extended overflow metabolism in various microorganisms. Microb. Cell Fac. 2012, 11, 122. [Google Scholar] [CrossRef] [Green Version]
- Jouhten, P.; Ponomarova, O.; Gonzalez, R.; Patil, K.R. Saccharomyces cerevisiae metabolism in ecological context. FEMS Yeast Res. 2016, 16, fow080. [Google Scholar] [CrossRef] [Green Version]
- Ponomarova, O.; Patil, K.R. Metabolic interactions in microbial communities: Untangling the Gordian knot. Curr. Opin. Microbiol. 2015, 27, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Campbell, K.; Vowinckel, J.; Ralser, M. Cell-to-cell heterogeneity emerges as consequence of metabolic cooperation in a synthetic yeast community. Biotech. J. 2016, 11, 1169–1178. [Google Scholar] [CrossRef] [Green Version]
- Campbell, K.; Vowinckel, J.; Mülleder, M.; Malmsheimer, S.; Lawrence, N.; Calvani, E.; Miller-Fleming, L.; Alam, M.T.; Christen, S.; Keller, M.A. Self-establishing communities enable cooperative metabolite exchange in a eukaryote. Elife 2015, 4, e09943. [Google Scholar] [CrossRef]
- Klosinska, M.M.; Crutchfield, C.A.; Bradley, P.H.; Rabinowitz, J.D.; Broach, J.R. Yeast cells can access distinct quiescent states. Genes Dev. 2011, 25, 336–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aragon, A.D.; Rodriguez, A.L.; Meirelles, O.; Roy, S.; Davidson, G.S.; Tapia, P.H.; Allen, C.; Joe, R.; Benn, D.; Werner-Washburne, M. Characterization of differentiated quiescent and nonquiescent cells in yeast stationary-phase cultures. Mol. Biol. Cell 2008, 19, 1271–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honigberg, S.M. Similar environments but diverse fates: Responses of budding yeast to nutrient deprivation. Microb. Cell 2016, 3, 302. [Google Scholar] [CrossRef]
- Werner-Washburne, M.; Roy, S.; Davidson, G.S. Aging and the survival of quiescent and non-quiescent cells in yeast stationary-phase cultures. Subcell. Biochem. 2012, 57, 123–143. [Google Scholar] [PubMed]
- Allen, C.P.; Anna-A, S.S.; Jaetao, J.E.; Werner-Washburne, M.C. Separation and characterization of quiescent and senescent cells from S-cerevisiae stationary-phase cultures. Mol. Biol. Cell 2004, 15, 367a–368a. [Google Scholar]
- Lee, H.Y.; Cheng, K.Y.; Chao, J.C.; Leu, J.Y. Differentiated cytoplasmic granule formation in quiescent and non-quiescent cells upon chronological aging. Microb. Cell 2016, 3, 109. [Google Scholar] [CrossRef] [Green Version]
- Wloch-Salamon, D.M.; Tomala, K.; Aggeli, D.; Dunn, B. Adaptive roles of SSY1 and SIR3 during cycles of growth and starvation in Saccharomyces cerevisiae populations enriched for quiescent or non-quiescent cells. G3 Genes Genomes Genet. 2017, 7, 1899–1911. [Google Scholar] [CrossRef] [Green Version]
- Krishna, S.; Laxman, S. A minimal “push–pull” bistability model explains oscillations between quiescent and proliferative cell states. Mol. Biol. Cell 2018, 29, 2243–2258. [Google Scholar] [CrossRef]
- Neiman, A.M. Sporulation in the Budding Yeast Saccharomyces cerevisiae. Genetics 2011, 189, 737–765. [Google Scholar] [CrossRef] [Green Version]
- Varahan, S.; Walvekar, A.; Sinha, V.; Krishna, S.; Laxman, S. Metabolic constraints drive self-organization of specialized cell groups. Elife 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Mortimer, R.K.; Johnston, J.R. Life span of individual yeast cells. Nature 1959, 183, 1751–1752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaeberlein, M. Lessons on longevity from budding yeast. Nature 2010, 464, 513–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, S.M.; Hanzén, S.; Nyström, T. Restricted access: Spatial sequestration of damaged proteins during stress and aging. EMBO Rep. 2017, 18, 377–391. [Google Scholar] [CrossRef]
- Levy, S.F.; Ziv, N.; Siegal, M.L. Bet hedging in yeast by heterogeneous, age-correlated expression of a stress protectant. PLoS Biol. 2012, 10, e1001325. [Google Scholar] [CrossRef] [Green Version]
- Wloch-Salamon, D.M.; Bem, A.E. Types of cell death and methods of their detection in yeast Saccharomyces cerevisiae. J. Appl. Microbiol. 2012, 114, 287–298. [Google Scholar] [CrossRef]
- Zadrag-Tecza, R.; Kwolek-Mirek, M.; Bartosz, G.; Bilinski, T. Cell volume as a factor limiting the replicative lifespan of the yeast Saccharomyces cerevisiae. Biogerontology 2009, 10, 481–488. [Google Scholar] [CrossRef]
- Kamei, Y.; Tamada, Y.; Nakayama, Y.; Fukusaki, E.; Mukai, Y. Changes in transcription and metabolism during the early stage of replicative cellular senescence in budding yeast. J. Biol. Chem. 2014, 289, 32081–32093. [Google Scholar] [CrossRef] [Green Version]
- Janssens, G.E.; Meinema, A.C.; Gonzalez, J.; Wolters, J.C.; Schmidt, A.; Guryev, V.; Bischoff, R.; Wit, E.C.; Veenhoff, L.M.; Heinemann, M. Protein biogenesis machinery is a driver of replicative aging in yeast. Elife 2015, 4, e08527. [Google Scholar] [CrossRef]
- Allen, C.; Buttner, S.; Aragon, A.D.; Thomas, J.A.; Meirelles, O.; Jaetao, J.E.; Benn, D.; Ruby, S.W.; Veenhuis, M.; Madeo, F.; et al. Isolation of quiescent and nonquiescent cells from yeast stationary-phase cultures. J. Cell Biol. 2006, 174, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Palkova, Z.; Janderová, B.; Gabriel, J.; Zikánová, B.; Pospíŝek, M.; Forstová, J. Ammonia mediates communication between yeast colonies. Nature 1997, 390, 532–536. [Google Scholar] [CrossRef] [PubMed]
- Bojsen, R.; Regenberg, B.; Folkesson, A. Saccharomyces cerevisiae biofilm tolerance towards systemic antifungals depends on growth phase. BMC Microbiol. 2014, 14, 305. [Google Scholar] [CrossRef] [Green Version]
- Bojsen, R.; Regenberg, B.; Folkesson, A. Persistence and drug tolerance in pathogenic yeast. Curr. Genet. 2017, 63, 19–22. [Google Scholar] [CrossRef] [PubMed]
- Pentz, J.T.; Taylor, B.P.; Ratcliff, W.C. Apoptosis in snowflake yeast: Novel trait, or side effect of toxic waste? J. R. Soc. Interface 2016, 13, 20160121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peter, J.; De Chiara, M.; Friedrich, A.; Yue, J.X.; Pflieger, D.; Bergström, A.; Sigwalt, A.; Barre, B.; Freel, K.; Llored, A.; et al. Genome evolution across 1,011 Saccharomyces cerevisiae isolates. Nature 2018, 556, 339. [Google Scholar] [CrossRef] [Green Version]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Opalek, M.; Wloch-Salamon, D. Aspects of Multicellularity in Saccharomyces cerevisiae Yeast: A Review of Evolutionary and Physiological Mechanisms. Genes 2020, 11, 690. https://doi.org/10.3390/genes11060690
Opalek M, Wloch-Salamon D. Aspects of Multicellularity in Saccharomyces cerevisiae Yeast: A Review of Evolutionary and Physiological Mechanisms. Genes. 2020; 11(6):690. https://doi.org/10.3390/genes11060690
Chicago/Turabian StyleOpalek, Monika, and Dominika Wloch-Salamon. 2020. "Aspects of Multicellularity in Saccharomyces cerevisiae Yeast: A Review of Evolutionary and Physiological Mechanisms" Genes 11, no. 6: 690. https://doi.org/10.3390/genes11060690
APA StyleOpalek, M., & Wloch-Salamon, D. (2020). Aspects of Multicellularity in Saccharomyces cerevisiae Yeast: A Review of Evolutionary and Physiological Mechanisms. Genes, 11(6), 690. https://doi.org/10.3390/genes11060690