EST–SNP Study of Olea europaea L. Uncovers Functional Polymorphisms between Cultivated and Wild Olives

 ,
, 
 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and EST—SNP Genotyping
2.2. Genetic Differentiation and Population Genetic Structure
2.3. Genetic Diversity
2.4. Identification and Functional Analysis of EST–SNP
3. Results
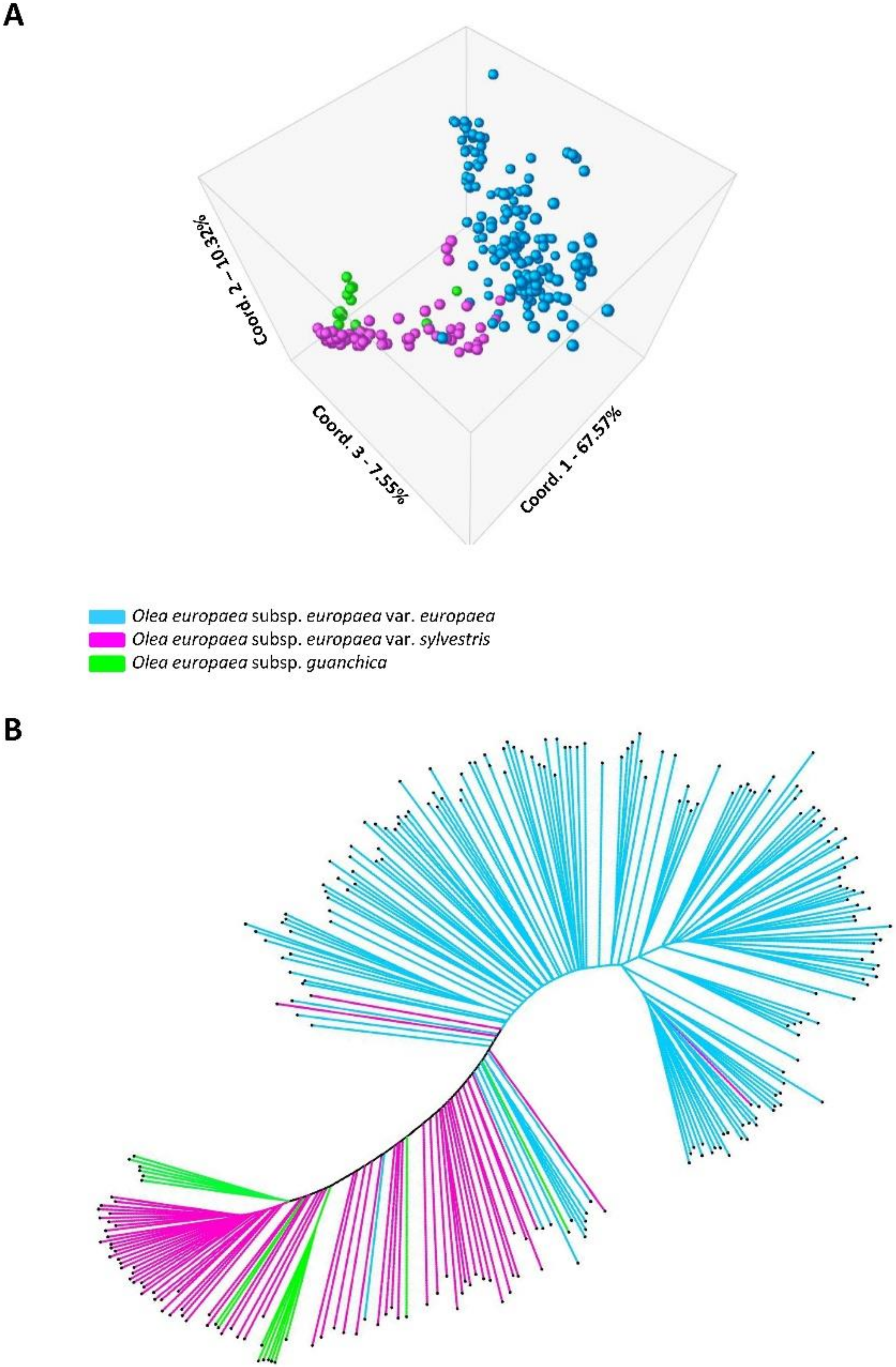
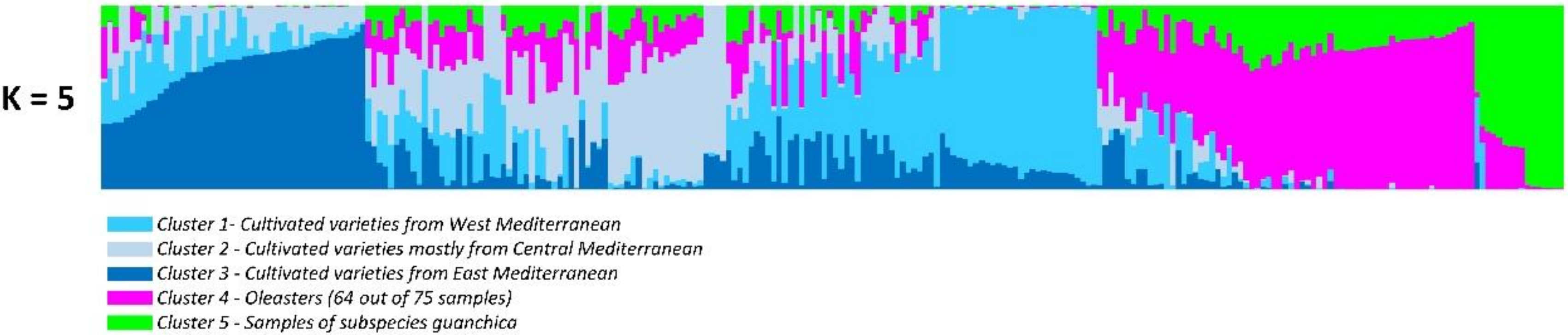
3.1. Genetic Differentiation and Population Structure
3.2. Genetic Diversity of the Cultivated Varieties, Oleasters, and Subsp. Guanchica Samples
3.3. Identification of Genes Carrying Polymorphisms between Cultivated and Wild Olives
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Barazani, O.; Keren-Keiserman, A.; Westberg, E.; Hanin, N.; Dag, A.; Ben-Ari, G.; Fragman-Sapir, O.; Tugendhaft, Y.; Kerem, Z.; Kadereit, J.W. Genetic variation of naturally growing olive trees in Israel: From abandoned groves to feral and wild? BMC Plant Biol. 2016, 16, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Rienzo, V.; Sion, S.; Taranto, F.; D’Agostino, N.; Montemurro, C.; Fanelli, V.; Sabetta, W.; Boucheffa, S.; Tamendjari, A.; Pasqualone, A.; et al. Genetic flow among olive populations within the Mediterranean basin. PeerJ 2018, 6, e5260. [Google Scholar] [CrossRef] [Green Version]
- Kassa, A.; Konrad, H.; Geburek, T. Molecular diversity and gene flow within and among different subspecies of the wild olive (Olea europaea L.): A review. Flora Morphol. Distrib. Funct. Ecol. Plants 2019, 250, 18–26. [Google Scholar] [CrossRef]
- Vargas, P.; Muñoz, G.; Hess, J.; Kadereit, J. Olea europaea subsp. guanchica and subsp. maroccana (Oleaceae), two new names for olive tree relatives. In Anales Del Jardín Botánico de Madrid; Instituto Botánico, Real Jardin de Botánico CSIC: Madrid, Spain, 2000; pp. 360–361. [Google Scholar]
- Green, P.S. A Revision of Olea L. (Oleaceae). Kew Bull. 2002, 57, 91–140. [Google Scholar] [CrossRef]
- Unver, T.; Wu, Z.; Sterck, L.; Turktas, M.; Lohaus, R.; Li, Z.; Yang, M.; He, L.; Deng, T.; Escalante, F.J.; et al. Genome of wild olive and the evolution of oil biosynthesis. Proc. Natl. Acad. Sci. USA 2017, 114, E9413–E9422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lumaret, R.; Ouazzani, N. Ancient wild olives in Mediterranean forests. Nature 2001, 413, 700. [Google Scholar] [CrossRef]
- Hmmam, I.; Mariotti, R.; Ruperti, B.; Cultrera, N.; Baldoni, L.; Barcaccia, G. Venetian olive (Olea europaea) germplasm: Disclosing the genetic identity of locally grown cultivars suited for typical extra virgin oil productions. Genet. Resour. Crop. Evol. 2018, 65, 1733–1750. [Google Scholar] [CrossRef]
- Lazović, B.; Adakalić, M.; Pucci, C.; Perović, T.; Bandelj, D.; Belaj, A.; Mariotti, R.; Baldoni, L. Characterizing ancient and local olive germplasm from Montenegro. Sci. Hortic. 2016, 209, 117–123. [Google Scholar] [CrossRef]
- Mousavi, S.; Mazinani, S.M.H.; Arzani, K.; Ydollahi, A.; Pandolfi, S.; Baldoni, L.; Mariotti, R. Molecular and morphological characterization of Golestan (Iran) olive ecotypes provides evidence for the presence of promising genotypes. Genet. Resour. Crop. Evol. 2014, 61, 775–785. [Google Scholar] [CrossRef]
- Mousavi, S.; Stanzione, V.; Mencuccini, M.; Baldoni, L.; Bufacchi, M.; Mariotti, R. Biochemical and molecular profiling of unknown olive genotypes from central Italy: Determination of major and minor components. Eur. Food Res. Technol. 2019, 245, 83–94. [Google Scholar] [CrossRef]
- Ninot, A.; Howad, W.; Aranzana, M.J.; Senar, R.; Romero, A.; Mariotti, R.; Baldoni, L.; Belaj, A. Survey of over 4, 500 monumental olive trees preserved on-farm in the northeast Iberian Peninsula, their genotyping and characterization. Sci. Hortic. 2018, 231, 253–264. [Google Scholar] [CrossRef] [Green Version]
- Besnard, G.; El Bakkali, A.; Haouane, H.; Baali-Cherif, D.; Moukhli, A.; Khadari, B. Population genetics of Mediterranean and Saharan olives: Geographic patterns of differentiation and evidence for early generations of admixture. Ann. Bot. 2013, 112, 1293–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazinani, S.M.H.; Mariotti, R.; Torkzaban, B.; Sheikh-Hassani, M.; Ataei, S.; Cultrera, N.; Pandolfi, S.; Baldoni, L. High Genetic Diversity Detected in Olives beyond the Boundaries of the Mediterranean Sea. PLoS ONE 2014, 9, e93146. [Google Scholar] [CrossRef] [Green Version]
- Belaj, A.; Gurbuz Veral, M.; Sikaoui, H.; Moukhli, A.; Khadari, B.; Mariotti, R.; Baldoni, L. Olive Genetic Resources. In The Olive Tree Genome, Compendium of Plant Genomes; Rugini, E., Baldoni, L., Muleo, R., Sebastiani, L., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 27–54. [Google Scholar]
- Mousavi, S.; Mariotti, R.; Bagnoli, F.; Costantini, L.; Cultrera, N.; Arzani, K.; Pandolfi, S.; Vendramin, G.G.; Torkzaban, B.; Mazinani, S.M.H.; et al. The eastern part of the Fertile Crescent concealed an unexpected route of olive (Olea europaea L.) differentiation. Ann. Bot. 2017, 119, 1305–1318. [Google Scholar] [CrossRef]
- Díez, C.M.; Trujillo, I.; Martinez-Urdiroz, N.; Barranco, D.; Rallo, L.; Marfil, P.; Gaut, B.S.; Ruiz, P.F.M. Olive domestication and diversification in the Mediterranean Basin. New Phytol. 2015, 206, 436–447. [Google Scholar] [CrossRef]
- Besnard, G.; De Casas, R.R. Single vs multiple independent olive domestications: The jury is (still) out. New Phytol. 2016, 209, 466–470. [Google Scholar] [CrossRef] [Green Version]
- Dighton, A.; Fairbairn, A.S.; Bourke, S.; Faith, J.T.; Habgood, P. Bronze Age olive domestication in the north Jordan valley: New morphological evidence for regional complexity in early arboricultural practice from Pella in Jordan. Veg. Hist. Archaeobotany 2017, 26, 403–413. [Google Scholar] [CrossRef]
- Julca, I.; Marcet-Houben, M.; Vargas, P.; Gabaldon, T. Phylogenomics of the olive tree (Olea europaea) reveals the relative contribution of ancient allo- and autopolyploidization events. BMC Biol. 2018, 16, 15. [Google Scholar] [CrossRef] [Green Version]
- Langgut, D.; Cheddadi, R.; Carrión, J.; Cavanagh, M.; Colombaroli, D.; Eastwood, W.J.; Greenberg, R.; Litt, T.; Mercuri, A.M.; Miebach, A.; et al. The origin and spread of olive cultivation in the Mediterranean Basin: The fossil pollen evidence. Holocene 2019, 29, 902–922. [Google Scholar] [CrossRef]
- Jiménez-Ruiz, J.; Ramírez-Tejero, J.A.; Fernández-Pozo, N.; Leyva-Pérez, M.d.l.O.; Yan, H.; Rosa, R.d.l.; Belaj, A.; Montes, E.; Rodríguez-Ariza, M.O.; Navarro, F.; et al. Transposon activation is a major driver in the genome evolution of cultivated olive trees ( Olea europaea L.). Plant Genome 2020, 13, e20010. [Google Scholar] [CrossRef] [Green Version]
- Besnard, G.; Terral, J.; Cornille, A. On the origins and domestication of the olive: A review and perspectives. Ann. Bot. 2018, 121, 385–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besnard, G.; Khadari, B.; Navascués, M.; Fernández-Mazuecos, M.; El Bakkali, A.; Arrigo, N.; Baali-Cherif, D.; De Caraffa, V.B.-B.; Santoni, S.; Vargas, P.; et al. The complex history of the olive tree: From Late Quaternary diversification of Mediterranean lineages to primary domestication in the northern Levant. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voropaeva, I.; Stika, H.-P. Plant remains from Roman period town of Obulco (today Porcuna) in Andalusia (Spain)—Distribution and domestication of olive in the west Mediterranean region. Archaeol. Anthr. Sci. 2018, 10, 865–882. [Google Scholar] [CrossRef]
- Naz, S.; Khan, M.R.; Awan, A.A.; Hussain, M.; Ali, S. Diversity and divergence in cultivated and wild olive germplasm collected from northern Pakistan. Int. J. Agric. Biol. 2019, 22, 1109–1115. [Google Scholar]
- Ashraf, U.; Peterson, A.T.; Chaudhry, M.N.; Ashraf, I.; Saqib, Z.; Ahmad, S.R.; Ali, H. Ecological niche model comparison under different climate scenarios: A case study of Oleaspp. in Asia. Ecosphere 2017, 8, e01825. [Google Scholar] [CrossRef]
- Qiu, J.; Zhou, Y.; Mao, L.; Ye, C.; Wang, W.; Zhang, J.; Yu, Y.; Fu, F.; Wang, Y.; Qian, F.; et al. Genomic variation associated with local adaptation of weedy rice during de-domestication. Nat. Commun. 2017, 8, 15323. [Google Scholar] [CrossRef]
- Lu, X.; Fu, X.; Wang, D.; Wang, J.; Chen, X.; Hao, M.; Wang, J.; Gervers, K.A.; Guo, L.; Wang, S.; et al. Resequencing of cv CRI-12 family reveals haplotype block inheritance and recombination of agronomically important genes in artificial selection. Plant Biotechnol. J. 2018, 17, 945–955. [Google Scholar] [CrossRef]
- Ruban, A.V. Plant science: Crops on the fast track for light. Nature 2017, 541, 36. [Google Scholar] [CrossRef]
- Arora, S.; Steuernagel, B.; Gaurav, K.; Chandramohan, S.; Long, Y.; Matny, O.; Johnson, R.; Enk, J.; Periyannan, S.; Singh, N.; et al. Resistance gene cloning from a wild crop relative by sequence capture and association genetics. Nat. Biotechnol. 2019, 37, 139–143. [Google Scholar] [CrossRef]
- Doebley, J.; Gaut, B.S.; Smith, B.D. The Molecular Genetics of Crop Domestication. Cell 2006, 127, 1309–1321. [Google Scholar] [CrossRef] [Green Version]
- Fan, C.; Xing, Y.; Mao, H.; Lu, T.; Han, B.; Xu, C.; Li, X.; Zhang, Q. GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor. Appl. Genet. 2006, 112, 1164–1171. [Google Scholar] [CrossRef] [PubMed]
- Luikart, G.; England, P.R.; Tallmon, D.; Jordan, S.; Taberlet, P. The power and promise of population genomics: From genotyping to genome typing. Nat. Rev. Genet. 2003, 4, 981–994. [Google Scholar] [CrossRef]
- Zhou, Y.; Massonnet, M.; Sanjak, J.S.; Cantu, D.; Gaut, B.S. Evolutionary genomics of grape (Vitis vinifera ssp. vinifera) domestication. Proc. Natl. Acad. Sci. USA 2017, 114, 11715–11720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, A.J.; Gross, B.L. From forest to field: Perennial fruit crop domestication. Am. J. Bot. 2011, 98, 1389–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- This, P.; Lacombe, T.; Cadle-Davidson, M.; Owens, C.L. Wine grape (Vitis vinifera L.) color associates with allelic variation in the domestication gene VvmybA1. Theor. Appl. Genet. 2007, 114, 723–730. [Google Scholar] [CrossRef]
- Gros-Balthazard, M.; Besnard, G.; Sarah, G.; Holtz, Y.; Leclercq, J.; Santoni, S.; Wegmann, D.; Glémain, S.; Khadari, B. Evolutionary transcriptomics reveals the origins of olives and the genomic changes associated with their domestication. Plant J. 2019, 100, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Mace, E.S.; Tai, S.; Cruickshank, A.; Campbell, B.C.; Zhao, X.; Van Oosterom, E.J.; Godwin, I.D.; Botella, M.A.; Jordan, D.R. Whole-Genome Analysis of Candidate genes Associated with Seed Size and Weight in Sorghum bicolor Reveals Signatures of Artificial Selection and Insights into Parallel Domestication in Cereal Crops. Front. Plant Sci. 2017, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Arbeiter, A.B.; Hladnik, M.; Jakše, J.; Bandelj, D. Identification and validation of novel EST-SSR markers in olives. Sci. Agricola 2017, 74, 215–225. [Google Scholar] [CrossRef]
- De La Rosa, R.; Belaj, A.; Muñoz-Mérida, A.; Trelles, O.; Ortíz-Martín, I.; González-Plaza, J.J.; Valpuesta, V.; Beuzón, C.R. Development of EST-derived SSR Markers with Long-core Repeat in Olive and Their Use for Paternity Testing. J. Am. Soc. Hortic. Sci. 2013, 138, 290–296. [Google Scholar] [CrossRef]
- Mariotti, R.; Cultrera, N.G.M.; Mousavi, S.; Baglivo, F.; Rossi, M.; Albertini, E.; Alagna, F.; Carbone, F.; Perrotta, G.; Baldoni, L. Development, evaluation, and validation of new EST-SSR markers in olive (Olea europaea L.). Tree Genet. Genomes 2016, 12, 120. [Google Scholar] [CrossRef]
- Kyriakopoulou, C.I.; Kalogianni, D.P. Genetic Identification of the Wild Form of Olive (Olea europaea var. sylvestris) Using Allele-Specific Real-Time PCR. Foods 2020, 9, 467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Niu, E.; Shi, A.; Mou, B. Genetic Diversity Analysis of Olive Germplasm (Olea europaea L.) With Genotyping-by-Sequencing Technology. Front. Genet. 2019, 10, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biton, I.; Doron-Faigenboim, A.; Jamwal, M.; Mani, Y.; Eshed, R.; Rosen, A.; Sherman, A.; Ophir, R.; Lavee, S.; Avidan, B.; et al. Development of a large set of SNP markers for assessing phylogenetic relationships between the olive cultivars composing the Israeli olive germplasm collection. Mol. Breed. 2015, 35, 107. [Google Scholar] [CrossRef]
- Ipek, A.; Yılmaz, K.; Sıkıcı, P.; Tangu, N.A.; Öz, A.T.; Bayraktar, M.; Ipek, M.; Gulen, H. SNP Discovery by GBS in Olive and the Construction of a High-Density Genetic Linkage Map. Biochem. Genet. 2016, 54, 313–325. [Google Scholar] [CrossRef]
- D’Agostino, N.; Taranto, F.; Camposeo, S.; Mangini, G.; Fanelli, V.; Gadaleta, S.; Miazzi, M.M.; Pavan, S.; Di Rienzo, V.; Sabetta, W.; et al. GBS-derived SNP catalogue unveiled wide genetic variability and geographical relationships of Italian olive cultivars. Sci. Rep. 2018, 8, 15877. [Google Scholar] [CrossRef] [Green Version]
- Belaj, A.; De La Rosa, R.; Lorite, I.J.; Mariotti, R.; Cultrera, N.G.; Beuzón, C.R.; González-Plaza, J.J.; Muñoz-Merida, A.; Trelles, O.; Baldoni, L. Usefulness of a New Large Set of High Throughput EST-SNP Markers as a Tool for Olive Germplasm Collection Management. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- García-Verdugo, C.; Fay, M.F.; Granado-Yela, C.; Rubio de Casas, R.; Balaguer, L.; Besnard, G.; Vargas, P. Genetic diversity and differentiation processes in the ploidy series of Olea europaea L.: A multiscale approach from subsp. to insular populations. Mol. Ecol. 2009, 18, 454–467. [Google Scholar] [CrossRef]
- Hess, J.; Kadereit, J.W.; Vargas, P. The colonization history of Olea europaea L. in Macaronesia based on internal transcribed spacer 1 (ITS-1) sequences, randomly amplified polymorphic DNAs (RAPD), and intersimple sequence repeats (ISSR). Mol. Ecol. 2000, 9, 857–868. [Google Scholar] [CrossRef] [Green Version]
- Peakall, R.; Smouse, P.E. Genalex 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; c1979–c2000; pp. 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGAX: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Earl, D.A.; Vonholdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2011, 4, 359–361. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; E Ramos-Onsins, S.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Thomas, D.C.; Campbell, M.J.; Kejariwal, A.; Mi, H.; Karlak, B.; Daverman, R.; Diemer, K.; Muruganujan, A.; Narechania, A. PANTHER: A Library of Protein Families and Subfamilies Indexed by Function. Genome Res. 2003, 13, 2129–2141. [Google Scholar] [CrossRef] [Green Version]
- Mastretta-Yanes, A.; Gasman, F.A.; Burgeff, C.; Ramírez, M.C.; Piñero, D.; Sarukhán, J. An Initiative for the Study and Use of Genetic Diversity of Domesticated Plants and Their Wild Relatives. Front. Plant Sci. 2018, 9, 209. [Google Scholar] [CrossRef] [Green Version]
- Mariotti, R.; Fornasiero, A.; Mousavi, S.; Cultrera, N.G.; Brizioli, F.; Pandolfi, S.; Passeri, V.; Rossi, M.; Magris, G.; Scalabrin, S.; et al. Genetic Mapping of the Incompatibility Locus in Olive and Development of a Linked Sequence-Tagged Site Marker. Front. Plant Sci. 2020, 10, 10. [Google Scholar] [CrossRef]
- Hedayati, V.; Mousavi, A.; Razavi, K.; Cultrera, N.; Alagna, F.; Mariotti, R.; Mazinani, S.M.H.; Baldoni, L. Polymorphisms in the AOX2 gene are associated with the rooting ability of olive cuttings. Plant Cell Rep. 2015, 34, 1151–1164. [Google Scholar] [CrossRef]
- Cultrera, N.G.M.; Sarri, V.; Lucentini, L.; Ceccarelli, M.; Alagna, F.; Mariotti, R.; Mousavi, S.; Ruiz, C.G.; Baldoni, L. High Levels of Variation Within Gene Sequences of Olea europaea L. Front. Plant Sci. 2019, 9, 1932. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Wei, C.-L.; Liu, H.-W.; Wu, J.-L.; Li, Z.-G.; Zhang, L.; Jian, J.-B.; Li, Y.-Y.; Tai, Y.-L.; Zhang, J.; et al. Genetic Divergence between Camellia sinensis and Its Wild Relatives Revealed via Genome-Wide SNPs from RAD Sequencing. PLoS ONE 2016, 11, e0151424. [Google Scholar] [CrossRef] [Green Version]
- Baldoni, L.; Tosti, N.; Ricciolini, C.; Belaj, A.; Arcioni, S.; Pannelli, G.; Germanà, M.A.; Mulas, M.; Porceddu, A. Genetic Structure of Wild and Cultivated Olives in the Central Mediterranean Basin. Ann. Bot. 2006, 98, 935–942. [Google Scholar] [CrossRef] [PubMed]
- Kaniewski, D.; Van Campo, E.; Boiy, T.; Terral, J.-F.; Khadari, B.; Besnard, G. Primary domestication and early uses of the emblematic olive tree: Palaeobotanical, historical and molecular evidence from the Middle East. Biol. Rev. 2012, 87, 885–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terral, J.-F.; Alonso, N.; Capdevila, R.B.I.; Chatti, N.; Fabre, L.; Fiorentino, G.; Marinval, P.; Jordà, G.P.; Pradat, B.; Rovira, N.; et al. Historical biogeography of olive domestication (Olea europaea L.) as revealed by geometrical morphometry applied to biological and archaeological material. J. Biogeogr. 2004, 31, 63–77. [Google Scholar] [CrossRef]
- Saumitou-Laprade, P.; Vernet, P.; Vekemans, X.; Billiard, S.; Gallina, S.; Essalouh, L.; Mhaïs, A.; Moukhli, A.; El Bakkali, A.; Barcaccia, G.; et al. Elucidation of the genetic architecture of self-incompatibility in olive: Evolutionary consequences and perspectives for orchard management. Evol. Appl. 2017, 10, 867–880. [Google Scholar] [CrossRef]
- Beghè, D.; Piotti, A.; Satovic, Z.; de la Rosa, R.; Belaj, A. Pollen-mediated gene flow and fine-scale spatial genetic structure in Olea europaea subsp. europaea var. sylvestris. Ann. Bot. 2017, 119, 671–679. [Google Scholar]
- Wang, J.; Chu, S.; Zhang, H.; Zhu, Y.; Cheng, H.; Yu, D. Development and application of a novel genome-wide SNP array reveals domestication history in soybean. Sci. Rep. 2016, 6, 20728. [Google Scholar] [CrossRef]
- Bettaieb, I.; Bouktila, D. Genome-wide analysis of NBS-encoding resistance genes in the Mediterranean olive tree (Olea europaea subsp. europaea var. europaea): Insights into their molecular diversity, evolution and function. Tree Genet. Genomes 2020, 16, 23. [Google Scholar] [CrossRef]
- Nelson, M.R.; Marnellos, G.; Kämmerer, S.; Hoyal, C.R.; Shi, M.M.; Cantor, C.R.; Braun, A. Large-Scale Validation of Single Nucleotide Polymorphisms in Gene Regions. Genome Res. 2004, 14, 1664–1668. [Google Scholar] [CrossRef] [Green Version]
- Bartoszewski, R.A.; Jablonsky, M.; Bartoszewska, S.; Stevenson, L.; Dai, Q.; Kappes, J.; Collawn, J.F.; Bebok, Z. A Synonymous Single Nucleotide Polymorphism in ΔF508CFTRAlters the Secondary Structure of the mRNA and the Expression of the Mutant Protein. J. Biol. Chem. 2010, 285, 28741–28748. [Google Scholar] [CrossRef] [Green Version]
- Sauna, Z.E.; Kimchi-Sarfaty, C. Understanding the contribution of synonymous mutations to human disease. Nat. Rev. Genet. 2011, 12, 683–691. [Google Scholar] [CrossRef]
- She, R.; Jarosz, D.F. Mapping Causal Variants with Single Nucleotide Resolution Reveals the Biochemical Drivers of Phenotypic Change. Cell 2018, 172, 478–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, X.; Li, M.-W.; Xie, M.; Liu, X.; Ni, M.; Shao, G.; Song, C.; Yim, A.K.-Y.; Tao, Y.; Wong, F.-L.; et al. Identification of a novel salt tolerance gene in wild soybean by whole-genome sequencing. Nat. Commun. 2014, 5, 4340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brogna, S.; McLeod, T.; Petric, M. The Meaning of NMD: Translate or Perish. Trends Genet. 2016, 32, 395–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mateyak, M.; Sasikumar, A.N.; Dunaway, S.; Kinzy, T.G. The Unique Evolutionary Distribution of Eukaryotic Elongation Factor 3. In Evolution of the Protein Synthesis Machinery and Its Regulation; Springer Science and Business Media LLC: Berlin, Germany, 2016; pp. 313–326. [Google Scholar]
- Elakhdar, A.; Ushijima, T.; Fukuda, M.; Yamashiro, N.; Kawagoe, Y.; Kumamaru, T. Eukaryotic peptide chain release factor 1 participates in translation termination of specific cysteine-poor prolamines in rice endosperm. Plant Sci. 2018, 281, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.-D.; Wang, J.; Wang, D.; Kong, Z.-Q.; Zhou, L.; Zhang, G.-Y.; Gui, Y.-J.; Li, J.-J.; Huang, J.-Q.; Wang, B.-L.; et al. Population genomics demystifies the defoliation phenotype in the plant pathogen Verticillium dahliae. New Phytol. 2019, 222, 1012–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerra, D.; Lamontanara, A.; Bagnaresi, P.; Orrù, L.; Rizza, F.; Zelasco, S.; Beghè, D.; Ganino, T.; Pagani, D.; Cattivelli, L.; et al. Transcriptome changes associated with cold acclimation in leaves of olive tree (Olea europaea L.). Tree Genet. Genomes 2015, 11, 1–24. [Google Scholar] [CrossRef]
- Van Dingenen, J.; Vermeersch, M.; De Milde, L.; Hulsmans, S.; De Winne, N.; Van Leene, J.; Gonzalez, N.; Dhondt, S.; De Jaeger, G.; Rolland, F.A.; et al. The role of HEXOKINASE1 in Arabidopsis leaf growth. Plant Mol. Biol. 2018, 99, 79–93. [Google Scholar] [CrossRef]
- Moyroud, E.; Kusters, E.; Monniaux, M.; Koes, R.; Parcy, F. LEAFY blossoms. Trends Plant Sci. 2010, 15, 346–352. [Google Scholar] [CrossRef]
- Fernández-Ocaña, A.M.; García-López, M.C.; Jiménez-Ruiz, J.; Saniger, L.; Macías, D.; Navarro, F.; Oya, R.; Belaj, A.; De La Rosa, R.; Corpas, F.J.; et al. Identification of a gene involved in the juvenile-to-adult transition (JAT) in cultivated olive trees. Tree Genet. Genomes 2010, 6, 891–903. [Google Scholar] [CrossRef]
- Liu, C.; Thong, Z.; Yu, H. Coming into bloom: The specification of floral meristems. Development 2009, 136, 3379–3391. [Google Scholar] [CrossRef] [Green Version]
- Ding, F.; Zhang, S.; Chen, H.; Peng, H.; Lu, J.; He, X.; Pan, J. Functional analysis of a homologue of the FLORICAULA/LEAFY gene in litchi (Litchi chinensis Sonn.) revealing its significance in early flowering process. Genes Genom. 2018, 40, 1259–1267. [Google Scholar] [CrossRef] [PubMed]
- Peña, L.; Martín-Trillo, M.; Juárez, J.; Pina, J.A.; Navarro, L.; Martinez-Zapater, J.M. Constitutive expression of Arabidopsis LEAFY or APETALA1 genes in citrus reduces their generation time. Nat. Biotechnol. 2001, 19, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Rhee, J.Y.; Lee, S.H.; Chung, G.C.; Park, S.J.; Segami, S.; Maeshima, M.; Choi, G. Functionally redundant LNG3 and LNG4 genes regulate turgor-driven polar cell elongation through activation of XTH17 and XTH24. Plant Mol. Biol. 2018, 97, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Iurlaro, A.; De Caroli, M.; Sabella, E.; De Pascali, M.; Rampino, P.; De Bellis, L.; Perrotta, C.; Dalessandro, G.; Piro, G.; Fry, S.C.; et al. Drought and Heat Differentially Affect XTH Expression and XET Activity and Action in 3-Day-Old Seedlings of Durum Wheat Cultivars with Different Stress Susceptibility. Front. Plant Sci. 2016, 7, 1686. [Google Scholar] [CrossRef]
- Abe, M.; Kaya, H.; Watanabe-Taneda, A.; Shibuta, M.; Yamaguchi, A.; Sakamoto, T.; Kurata, T.; Ausin, I.; Araki, T.; Alonso-Blanco, C. FE, a phloem-specific Myb-related protein, promotes flowering through transcriptional activation ofFLOWERING LOCUS TandFLOWERING LOCUS T INTERACTING PROTEIN 1. Plant J. 2015, 83, 1059–1068. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Rao, P.; Yang, X.; Su, X.; Zhao, T.; Gao, K.; Yang, X.; An, X. A Global View of Transcriptome Dynamics during Male Floral Bud Development in Populus tomentosa. Sci. Rep. 2018, 8, 722. [Google Scholar] [CrossRef] [Green Version]
- Haberman, A.; Bakhshian, O.; Cerezo-Medina, S.; Paltiel, J.; Adler, C.; Ben-Ari, G.; Mercado, J.A.; Pliego-Alfaro, F.; Lavee, S.; Samach, A. A possible role for flowering locus T-encoding genes in interpreting environmental and internal cues affecting olive (Olea europaea L.) flower induction. Plant Cell Environ. 2017, 40, 1263–1280. [Google Scholar] [CrossRef]
- Koo, B.-H.; Yoo, S.-C.; Park, J.-W.; Kwon, C.-T.; Lee, B.-D.; An, G.; Zhang, Z.; Li, J.; Li, Z.; Paek, N.-C. Natural Variation in OsPRR37 Regulates Heading Date and Contributes to Rice Cultivation at a Wide Range of Latitudes. Mol. Plant 2013, 6, 1877–1888. [Google Scholar] [CrossRef] [Green Version]
- Song, S.; Li, L.; Li, Y.; Wang, T.; Fu, X. Overexpression of gene OsSUI1 affects floral organ development in rice (Oryza sativa L.). Mol. Breed. 2018, 38, 4. [Google Scholar] [CrossRef]
- Bates, P.D. Understanding the control of acyl flux through the lipid metabolic network of plant oil biosynthesis. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2016, 1861, 1214–1225. [Google Scholar] [CrossRef]
- Liu, M.; Li, X.; Liu, Y.; Shi, Y.; Ma, X. Analysis of differentially expressed genes under UV-B radiation in the desert plant Reaumuria soongorica. Gene 2015, 574, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Grienenberger, E.; Kim, S.S.; Lallemand, B.; Geoffroy, P.; Heintz, D.; Souza, C.d.A.; Heitz, T.; Douglas, C.J.; Legrand, M. Analysis of TETRAKETIDE α-PYRONE REDUCTASE Function in Arabidopsis thaliana Reveals a Previously Unknown, but Conserved, Biochemical Pathway in Sporopollenin Monomer Biosynthesis. Plant Cell 2010, 22, 4067–4083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hölscher, C.; Lutterbey, M.-C.; Lansing, H.; Meyer, T.; Fischer, K.; Von Schaewen, A. Defects in Peroxisomal 6-Phosphogluconate Dehydrogenase Isoform PGD2 Prevent Gametophytic Interaction in Arabidopsis thaliana. Plant Physiol. 2016, 171, 192–205. [Google Scholar] [CrossRef] [Green Version]
- Cao-Pham, A.H.; Urano, D.; Ross-Elliott, T.J.; Jones, A.M. Nudge-nudge, WNK-WNK (kinases), say no more? New Phytol. 2018, 220, 35–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.-M.; Zhao, W.-Q.; Yang, W.-X.; Liu, F.; Chen, J.-P.; Goyer, C.; Liu, D.-Q. Characterization of a Hypersensitive Response-Induced Gene TaHIR3 from Wheat Leaves Infected with Leaf Rust. Plant Mol. Biol. Rep. 2012, 31, 314–322. [Google Scholar] [CrossRef]
- Zhou, L.; Cheung, M.-Y.; Li, M.-W.; Fu, Y.; Sun, Z.; Sun, S.S.; Lam, H.-M. Rice Hypersensitive Induced Reaction Protein 1 (OsHIR1) associates with plasma membrane and triggers hypersensitive cell death. BMC Plant Biol. 2010, 10, 290. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Sun, Y.; Pei, Y.; Zhang, X.; Wang, P.; Li, X.; Li, F.; Hou, Y. A Pectin Methylesterase Inhibitor Enhances Resistance to Verticillium Wilt. Plant Physiol. 2018, 176, 2202–2220. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-N.; Lee, K.-H.; Kim, C.S. Abscisic acid receptor PYRABACTIN RESISTANCE-LIKE 8, PYL8, is involved in glucose response and dark-induced leaf senescence in Arabidopsis. Biochem. Biophys. Res. Commun. 2015, 463, 24–28. [Google Scholar] [CrossRef]
- Yang, H.; Zhao, L.; Zhao, S.; Wang, J.; Shi, H. Biochemical and transcriptomic analyses of drought stress responses of LY1306 tobacco strain. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Chen, S.; Xie, L.; Lu, Z.; Liu, M.; Han, X.; Qiao, G.; Jiang, J.; Zhuo, R.; Qiu, W.; et al. Overexpression of cysteine protease gene from Salix matsudana enhances salt tolerance in transgenic Arabidopsis. Environ. Exp. Bot. 2018, 147, 53–62. [Google Scholar] [CrossRef]
- Park, K.Y.; Kim, W.T.; Kim, E.Y. The proper localization of RESPONSIVE TO DESICCATION 20 in lipid droplets depends on their biogenesis induced by STRESS-RELATED PROTEINS in vegetative tissues. Biochem. Biophys. Res. Commun. 2018, 495, 1885–1889. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Bai, X.; Zhao, W.; Xiang, D.; Wan, Y.; Yan, J.; Zou, L.; Zhao, G. De Novo Assembly and Analysis of Tartary Buckwheat (Fagopyrum tataricum Garetn.) Transcriptome Discloses Key Regulators Involved in Salt-Stress Response. Genes 2017, 8, 255. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.-J.D.; Liebrand, T.W.; Yadeta, K.A.; Coaker, G. PBL13 Is a Serine/Threonine Protein Kinase That Negatively Regulates Arabidopsis Immune Responses. Plant Physiol. 2015, 169, 2950–2962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Movahedi, A.; Sun, W.; Zhang, J.; Wu, X.; Mousavi, M.; Mohammadi, K.; Yin, T.; Zhuge, Q. RNA-directed DNA methylation in plants. Plant Cell Rep. 2015, 34, 1857–1862. [Google Scholar] [CrossRef]
- Dufoo-Hurtado, M.D.; Huerta-Ocampo, J.Ã.; Barrera-Pacheco, A.; Rosa, A.P.B.d.l.; Mercado-Silva, E.M. Low temperature conditioning of garlic (Allium sativum L.) “seed” cloves induces alterations in sprouts proteome. Front. Plant. Sci. 2015, 6, 332. [Google Scholar] [CrossRef] [Green Version]
- Xin, S.; Yu, G.; Sun, L.; Qiang, X.; Xu, N.; Cheng, X. Expression of tomato SlTIP2;2 enhances the tolerance to salt stress in the transgenic Arabidopsis and interacts with target proteins. J. Plant. Res. 2014, 127, 695–708. [Google Scholar] [CrossRef]
- Chen, X.; Snyder, C.L.; Truksa, M.; Shah, S.; Weselake, R.J. sn-Glycerol-3-phosphate acyltransferases in plants. Plant. Signal. Behav. 2011, 6, 1695–1699. [Google Scholar] [CrossRef] [Green Version]
- Bontpart, T.; Cheynier, V.; Ageorges, A.; Terrier, N. BAHD or SCPL acyltransferase? What a dilemma for acylation in the world of plant phenolic compounds. New Phytol. 2015, 208, 695–707. [Google Scholar] [CrossRef]
- Lijuan, C.; Huiming, G.; Yi, L.; Hongmei, C. Chalcone synthase EaCHS1 from Eupatorium adenophorum functions in salt stress tolerance in tobacco. Plant. Cell Rep. 2015, 34, 885–894. [Google Scholar] [CrossRef]
- Zhang, X.; Abrahan, C.; Colquhoun, T.A.; Liu, C.-J. A Proteolytic Regulator Controlling Chalcone Synthase Stability and Flavonoid Biosynthesis in Arabidopsis. Plant. Cell 2017, 29, 1157–1174. [Google Scholar] [CrossRef] [Green Version]
- Reale, S.; Doveri, S.; Díaz, A.; Angiolillo, A.; Lucentini, L.; Pilla, F.; Martín, A.; Donini, P.; Lee, D. SNP-based markers for discriminating olive (Olea europaea L.) cultivars. Genome 2006, 49, 1193–1205. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Kumar, S.R.; Dwivedi, V.; Rai, A.; Shukla, A.K.; Shanker, K.; Nagegowda, D.A. Precursor feeding studies and molecular characterization of geraniol synthase establish the limiting role of geraniol in monoterpene indole alkaloid biosynthesis in Catharanthus roseus leaves. Plant. Sci. 2015, 239, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Alagna, F.; Geu-Flores, F.; Kries, H.; Panara, F.; Baldoni, L.; O’Connor, S.E.; Osbourn, A. Identification and Characterization of the Iridoid Synthase Involved in Oleuropein Biosynthesis in Olive (Olea europaea) Fruits. J. Biol. Chem. 2016, 291, 5542–5554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Servili, M.; Sordini, B.; Esposto, S.; Taticchi, A.; Urbani, S.; Sebastiani, L. Metabolomics of Olive Fruit: A Focus on the Secondary Metabolites. In The Olive Tree Genome, Compendium of Plant Genomes; Springer Science and Business Media LLC: Cham, Switzerland, 2016; pp. 123–139. [Google Scholar]
- Wong, D.C.J.; Pichersky, E.; Peakall, R. The Biosynthesis of Unusual Floral Volatiles and Blends Involved in Orchid Pollination by Deception: Current Progress and Future Prospects. Front. Plant. Sci. 2017, 8, 1955. [Google Scholar] [CrossRef] [Green Version]
- Keereetaweep, J.; Liu, H.; Zhai, Z.; Shanklin, J. Biotin Attachment Domain-Containing Proteins Irreversibly Inhibit Acetyl CoA Carboxylase. Plant. Physiol. 2018, 177, 208–215. [Google Scholar] [CrossRef] [Green Version]
- Liao, B.; Hao, Y.; Lu, J.-X.; Bai, H.; Guan, L.; Zhang, T. Transcriptomic analysis of Perilla frutescens seed to insight into the biosynthesis and metabolic of unsaturated fatty acids. BMC Genom. 2018, 19, 213. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Group 1 | Group 2 | Ks | Kxy | Gst | DeltaSt | GammaSt | Nst | Fst | Dxy | Da |
|---|---|---|---|---|---|---|---|---|---|---|
| Cultivars | Wild olives | 19.837 *** | 25.770 *** | 0.000 | 0.043 | 0.123 | 0.330 | 0.253 | 0.405 | 0.102 |
| Cultivars | subsp. guanchica | 20.128 *** | 25.492 *** | 0.006 | 0.020 | 0.062 | 0.402 | 0.323 | 0.400 | 0.129 |
| Wild olives | subsp. guanchica | 17.052 *** | 17.386 *** | 0.004 | 0.008 | 0.032 | 0.107 | 0.092 | 0.273 | 0.025 |
| Group | N | Na | Ne | Ho | He | UHe | F | |
|---|---|---|---|---|---|---|---|---|
| Cultivar | Mean | 170.43 | 2.013 | 1.536 | 0.369 | 0.324 | 0.325 | −0.120 |
| SE | 0.031 | 0.004 | 0.009 | 0.005 | 0.004 | 0.004 | 0.005 | |
| var. sylvestris | Mean | 72.92 | 1.976 | 1.448 | 0.295 | 0.275 | 0.276 | −0.055 |
| SE | 0.011 | 0.005 | 0.010 | 0.006 | 0.005 | 0.005 | 0.006 | |
| subsp. guanchica | Mean | 15.92 | 1.738 | 1.345 | 0.214 | 0.206 | 0.212 | −0.027 |
| SE | 0.012 | 0.014 | 0.011 | 0.007 | 0.006 | 0.006 | 0.009 | |
| Total | Mean | 86.43 | 1.909 | 1.443 | 0.293 | 0.268 | 0.271 | −0.071 |
| SE | 1.142 | 0.005 | 0.006 | 0.004 | 0.003 | 0.003 | 0.004 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mariotti, R.; Belaj, A.; De La Rosa, R.; Leòn, L.; Brizioli, F.; Baldoni, L.; Mousavi, S. EST–SNP Study of Olea europaea L. Uncovers Functional Polymorphisms between Cultivated and Wild Olives. Genes 2020, 11, 916. https://doi.org/10.3390/genes11080916
Mariotti R, Belaj A, De La Rosa R, Leòn L, Brizioli F, Baldoni L, Mousavi S. EST–SNP Study of Olea europaea L. Uncovers Functional Polymorphisms between Cultivated and Wild Olives. Genes. 2020; 11(8):916. https://doi.org/10.3390/genes11080916
Chicago/Turabian StyleMariotti, Roberto, Angjelina Belaj, Raul De La Rosa, Lorenzo Leòn, Federico Brizioli, Luciana Baldoni, and Soraya Mousavi. 2020. "EST–SNP Study of Olea europaea L. Uncovers Functional Polymorphisms between Cultivated and Wild Olives" Genes 11, no. 8: 916. https://doi.org/10.3390/genes11080916
APA StyleMariotti, R., Belaj, A., De La Rosa, R., Leòn, L., Brizioli, F., Baldoni, L., & Mousavi, S. (2020). EST–SNP Study of Olea europaea L. Uncovers Functional Polymorphisms between Cultivated and Wild Olives. Genes, 11(8), 916. https://doi.org/10.3390/genes11080916