
Are Bacterio- and Phytoplankton Community Compositions Related in Lakes Differing in Their Cyanobacteria Contribution and Physico-Chemical Properties?


Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Water Samples
2.2. Phytoplankton Analysis and Physicochemical Parameters
2.3. Bacterioplankton Community Composition
2.4. Statistical Analysis
3. Results
3.1. Physico-Chemical Characteristics
3.2. Phytoplankton Composition
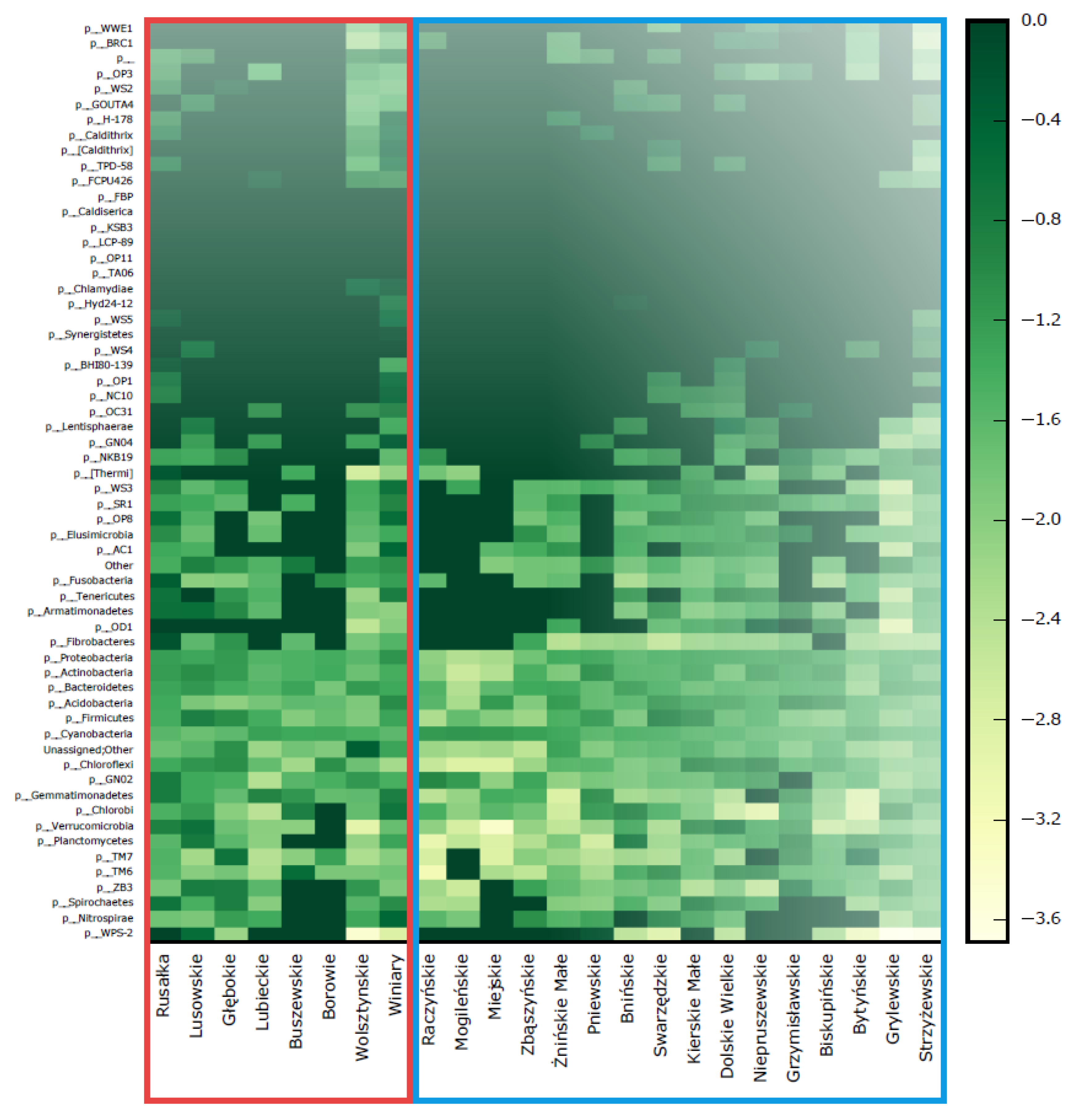
3.3. Bacterioplankton Community Composition
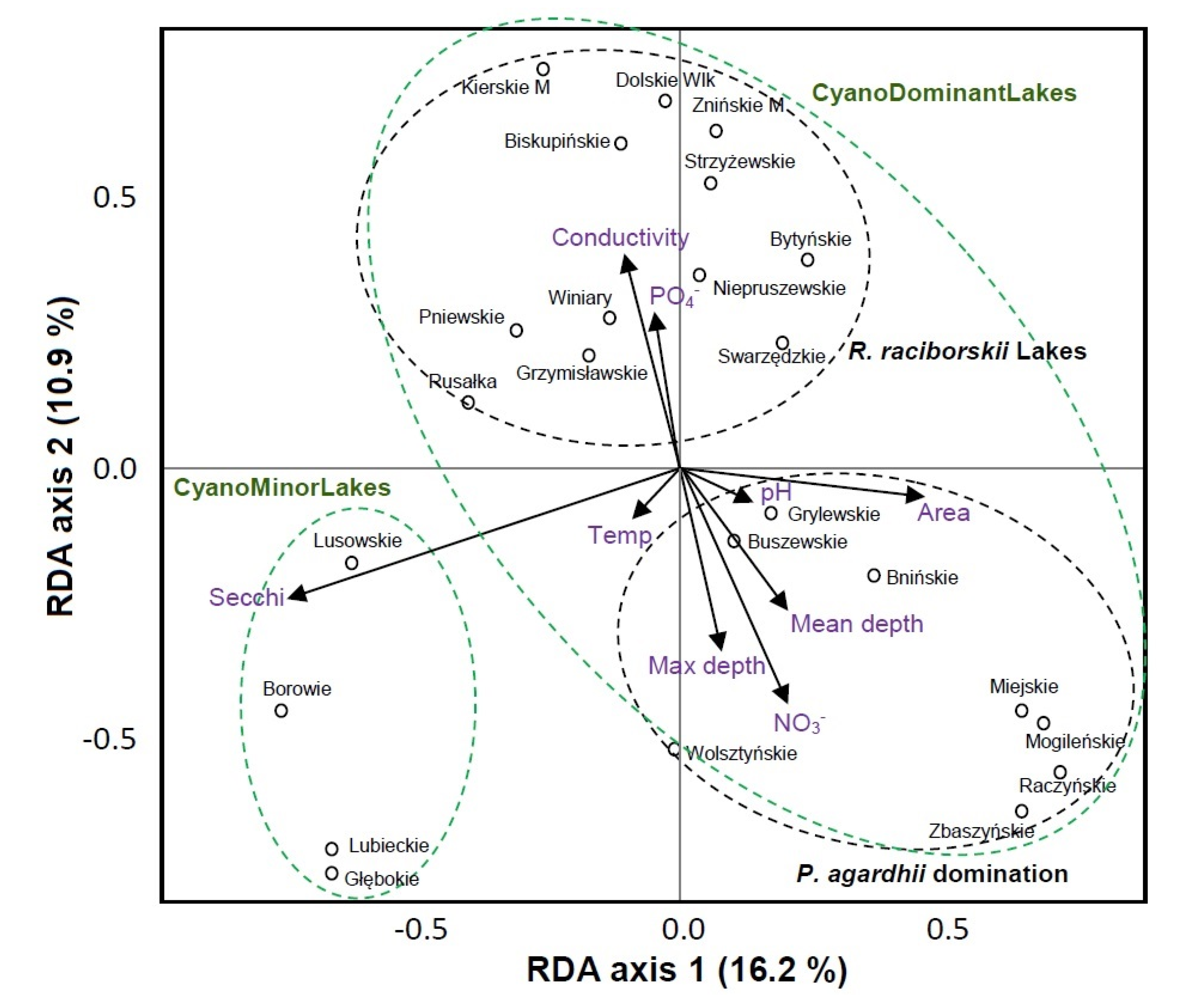
3.4. Community Patterns and Drivers
4. Discussion
4.1. Environmental Factors Driving the Bacteria and Phytoplankton Community Composition
4.2. The Bacteria and Phytoplankton Community Composition across Different Types of Lakes and the Relations within Microbial Communities
4.3. Congruence in the Composition of Microbial Community in the Context of Nitrification and Denitrification
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mur, L.R.; Schreurs, H. Light as a selective factor in the distribution of phytoplankton species. Water Sci. Technol. 1995, 32. [Google Scholar] [CrossRef]
- Guildford, S.J.; Hecky, R.E. Total nitrogen, total phosphorus, and nutrient limitation in lakes and oceans: Is there a common relationship? Limnol. Oceanogr. 2000, 45, 1213–1223. [Google Scholar] [CrossRef] [Green Version]
- Wetzel, R.G. Limnology, 2nd ed.; Saunders College Publishing: Philadelphia, PA, USA, 1983. [Google Scholar]
- Reynolds, C.S. Functional morphology and the adaptive strategies of freshwater phytoplankton. In Growth and Reproductive Strategies of Freshwater Phytoplankton; Sandgren, C.S., Ed.; Cambridge University Press: New York, NY, USA, 1988; pp. 388–433. [Google Scholar]
- Jeppesen, E.; Peder Jensen, J.; SØndergaard, M.; Lauridsen, T.; Landkildehus, F. Trophic structure, species richness and biodiversity in Danish lakes: Changes along a phosphorus gradient. Freshw. Biol. 2000, 45, 201–218. [Google Scholar] [CrossRef]
- Ferguson, H.W.; Claxton, M.J.; Moccia, R.D.; Wilkie, E.J. The quantitative clearance of bacteria from the bloodstream of rainbow trout (Salmo gairdneri). Vet. Pathol. 1982, 19, 687–699. [Google Scholar] [CrossRef] [Green Version]
- Wetzel, R.G. Death, detritus, and energy flow in aquatic ecosystems. Freshw. Biol. 1995, 33, 83–89. [Google Scholar] [CrossRef]
- Nealson, K.H. Sediment bacteria: Who’s there, what are they doing, and what’s new? Annu. Rev. Earth Planet. Sci. 1997, 25, 403–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpenter, S.R.; Pace, M.L. Dystrophy and eutrophy in lake ecosystems: Implications of fluctuating inputs. Oikos 1997, 78, 3. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, C.S. River plankton: The paradigm regained. In The Ecological Basis for River Management; Herper, M., Ferouson, N.J.D., Eds.; Wiley: Chichester, UK; Wiley: New York, NY, USA; Wiley: Brisbane, Australia; Wiley: Toronto, ON, Canada; Wiley: Singapore, 1995; pp. 161–174. [Google Scholar]
- Dziga, D.; Tokodi, N.; Drobac, D.; Kokociński, M.; Antosiak, A.; Puchalski, J.; Strzałka, W.; Madej, M.; Svirčev, Z.; Meriluoto, J. The effect of a combined hydrogen peroxide-MlrA treatment on the phytoplankton community and microcystin concentrations in a mesocosm experiment in lake Ludoš. Toxins 2019, 11, 725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dziga, D.; Maksylewicz, A.; Maroszek, M.; Marek, S. Combined treatment of toxic cyanobacteria Microcystis aeruginosa with hydrogen peroxide and microcystin biodegradation agents results in quick toxins elimination. Acta Biochim. Pol. 2018, 65, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Pal, M.; Yesankar, P.J.; Dwivedi, A.; Qureshi, A. Biotic control of harmful algal blooms (HABs): A brief review. J. Environ. Manag. 2020, 268, 110687. [Google Scholar] [CrossRef]
- Ndlela, L.L.; Oberholster, P.J.; Van Wyk, J.H.; Cheng, P.H. Bacteria as biological control agents of freshwater cyanobacteria: Is it feasible beyond the laboratory? Appl. Microbiol. Biotechnol. 2018, 102, 9911–9923. [Google Scholar] [CrossRef]
- Kent, A.D.; Yannarell, A.C.; Rusak, J.A.; Triplett, E.W.; McMahon, K.D. Synchrony in aquatic microbial community dynamics. ISME J. 2007, 1, 38–47. [Google Scholar] [CrossRef]
- Soininen, J.; Korhonen, J.J.; Karhu, J.; Vetterli, A. Disentangling the spatial patterns in community composition of prokaryotic and eukaryotic lake plankton. Limnol. Oceanogr. 2011, 56, 508–520. [Google Scholar] [CrossRef]
- Xiao, M.; Li, M.; Reynolds, C.S. Colony formation in the cyanobacterium Microcystis. Biol. Rev. 2018, 93, 1399–1420. [Google Scholar] [CrossRef] [Green Version]
- Pineda-Mendoza, R.M.; Briones-Roblero, C.I.; Gonzalez-Escobedo, R.; Rivera-Orduña, F.N.; Martínez-Jerónimo, F.; Zúñiga, G. Seasonal changes in the bacterial community structure of three eutrophicated urban lakes in Mexico city, with emphasis on Microcystis spp. Toxicon 2020, 179, 8–20. [Google Scholar] [CrossRef]
- Paver, S.F.; Hayek, K.R.; Gano, K.A.; Fagen, J.R.; Brown, C.T.; Davis-Richardson, A.G.; Crabb, D.B.; Rosario-Passapera, R.; Giongo, A.; Triplett, E.W.; et al. Interactions between specific phytoplankton and bacteria affect lake bacterial community succession. Environ. Microbiol. 2013, 15, 2489–2504. [Google Scholar] [CrossRef]
- Newton, R.J.; Kent, A.D.; Triplett, E.W.; McMahon, K.D. Microbial community dynamics in a humic lake: Differential persistence of common freshwater phylotypes. Environ. Microbiol. 2006, 8, 956–970. [Google Scholar] [CrossRef]
- Jones, S.E.; Cadkin, T.A.; Newton, R.J.; McMahon, K.D. Spatial and temporal scales of aquatic bacterial β diversity. Front. Microbiol. 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Shade, A.; Peter, H.; Allison, S.D.; Baho, D.L.; Berga, M.; Bürgmann, H.; Huber, D.H.; Langenheder, S.; Lennon, J.T.; Martiny, J.B.H.; et al. Fundamentals of microbial community resistance and resilience. Front. Microbiol. 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Shapleigh, J.P. Denitrifying prokaryotes. In The Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2013; pp. 405–425. [Google Scholar]
- Dziga, D.; Kokociński, M.; Barylski, J.; Nowicki, G.; Maksylewicz, A.; Antosiak, A.; Banaś, A.K.; Strzałka, W. Correlation between specific groups of heterotrophic bacteria and microcystin biodegradation in freshwater bodies of central Europe. FEMS Microbiol. Ecol. 2019, 95. [Google Scholar] [CrossRef]
- Jiang, Y.; Xiao, P.; Liu, Y.; Wang, J.; Li, R. Targeted deep sequencing reveals high diversity and variable dominance of bloom-forming cyanobacteria in eutrophic lakes. Harmful Algae 2017, 64, 42–50. [Google Scholar] [CrossRef]
- Bagatini, I.L.; Eiler, A.; Bertilsson, S.; Klaveness, D.; Tessarolli, L.P.; Vieira, A.A.H. Host-Specificity and dynamics in bacterial communities associated with bloom-forming freshwater phytoplankton. PLoS ONE 2014, 9, e85950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antosiak, A.; Tokodi, N.; Maziarz, R.; Kokociński, M.; Brzozowska, A.; Strzałka, W.; Banaś, A.K.; Willis, A.; Dziga, D. Different gene expression response of Polish and Australian Raphidiopsis raciborskii strains to the chill/light stress. Appl. Sci. 2020, 10, 5437. [Google Scholar] [CrossRef]
- Vico, P.; Bonilla, S.; Cremella, B.; Aubriot, L.; Iriarte, A.; Piccini, C. Biogeography of the cyanobacterium Raphidiopsis (Cylindrospermopsis) raciborskii: Integrating genomics, phylogenetic and toxicity data. Mol. Phylogenet. Evol. 2020, 148, 106824. [Google Scholar] [CrossRef]
- Hillebrand, H.; Dürselen, C.-D.; Kirschtel, D.; Pollingher, U.; Zohary, T. Biovolume calculation for pelagic and benthic microalgae. J. Phycol. 1999, 35, 403–424. [Google Scholar] [CrossRef]
- Napiórkowska-Krzebietke, A.; Kobos, J. Assessment of the cell biovolume of phytoplankton widespread in coastal and inland water bodies. Water Res. 2016, 104, 532–546. [Google Scholar] [CrossRef]
- Wetzel, R.G.; Likens, G.E. Limnological Analyses, 3rd ed.; Springer: New York, NY, USA, 2000. [Google Scholar]
- Ziesemer, K.A.; Mann, A.E.; Sankaranarayanan, K.; Schroeder, H.; Ozga, A.T.; Brandt, B.W.; Zaura, E.; Waters-Rist, A.; Hoogland, M.; Salazar-García, D.C.; et al. Intrinsic challenges in ancient microbiome reconstruction using 16S rRNA gene amplification. Sci. Rep. 2015, 5, 16498. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematic; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Kokociński, M.; Gągała, I.; Jasser, I.; Karosienė, J.; Kasperovičienė, J.; Kobos, J.; Koreivienė, J.; Soininen, J.; Szczurowska, A.; Woszczyk, M.; et al. Distribution of invasive Cylindrospermopsis raciborskii in the East-Central Europe is driven by climatic and local environmental variables. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef] [Green Version]
- Rücker, J.; Wiedner, C.; Zippel, P. Factors controlling the dominance of Planktothrix agardhii and Limnothrix redekei in eutrophic shallow lakes. Hydrobiologia 1997, 342, 107–115. [Google Scholar] [CrossRef]
- Wejnerowski, Ł.; Rzymski, P.; Kokociński, M.; Meriluoto, J. The structure and toxicity of winter cyanobacterial bloom in a eutrophic lake of the temperate zone. Ecotoxicology 2018, 27, 752–760. [Google Scholar] [CrossRef] [Green Version]
- Mankiewicz-Boczek, J. Application of molecular tools in ecohydrology. Ecohydrol. Hydrobiol. 2012, 12, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Te, S.H.; Tan, B.F.; Thompson, J.R.; Gin, K.Y.-H. Relationship of Microbiota and Cyanobacterial Secondary Metabolites in Planktothricoides -Dominated Bloom. Environ. Sci. Technol. 2017, 51, 4199–4209. [Google Scholar] [CrossRef]
- Parveen, B.; Ravet, V.; Djediat, C.; Mary, I.; Quiblier, C.; Debroas, D.; Humbert, J.-F. Bacterial communities associated with Microcystis colonies differ from free-living communities living in the same ecosystem. Environ. Microbiol. Rep. 2013, 5, 716–724. [Google Scholar] [CrossRef]
- Zheng, X.; Xiao, L.; Ren, J.; Yang, L. The effect of a microcystis aeruginosa bloom on the bacterioplankton community composition of lake Xuanwa. J. Freshw. Ecol. 2008, 23, 297–304. [Google Scholar] [CrossRef] [Green Version]
- Koo, H.; Mojib, N.; Hakim, J.A.; Hawes, I.; Tanabe, Y.; Andersen, D.T.; Bej, A.K. Microbial communities and their predicted metabolic functions in growth laminae of a unique large conical mat from lake Untersee, East Antarctica. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, H.; Xu, J.; Lavoie, M.; Fan, X.; Liu, G.; Sun, L.; Fu, Z.; Qian, H. Biological and chemical factors driving the temporal distribution of cyanobacteria and heterotrophic bacteria in a eutrophic lake (West Lake, China). Appl. Microbiol. Biotechnol. 2017, 101, 1685–1696. [Google Scholar] [CrossRef] [PubMed]
- Bouvy, M.; Pagano, M.; Troussellier, M. Effects of a cyanobacterial bloom (Cylindrospermopsis raciborskii) on bacteria and zooplankton communities in Ingazeira reservoir (northeast Brazil). Aquat. Microb. Ecol. 2001, 25, 215–227. [Google Scholar] [CrossRef] [Green Version]
- Woodhouse, J.N.; Ziegler, J.; Grossart, H.-P.; Neilan, B.A. Cyanobacterial community composition and bacteria–bacteria interactions promote the stable occurrence of particle-associated bacteria. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, C.; Thomas, T.; Lewis, M.; Steinberg, P.; Kjelleberg, S. Composition, uniqueness and variability of the epiphytic bacterial community of the green alga Ulva australis. ISME J. 2011, 5, 590–600. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Cai, Y.; Kong, F.; Yu, Y. Changes in abundance and community structure of bacteria associated with buoyant Microcystis colonies during the decline of cyanobacterial bloom (autumn–winter transition). Ann. Limnol. Int. J. Limnol. 2011, 47, 355–362. [Google Scholar] [CrossRef] [Green Version]
- Adam, B.; Klawonn, I.; Svedén, J.B.; Bergkvist, J.; Nahar, N.; Walve, J.; Littmann, S.; Whitehouse, M.J.; Lavik, G.; Kuypers, M.M.M.; et al. N2-Fixation, ammonium release and N-transfer to the microbial and classical food web within a plankton community. ISME J. 2016, 10, 450–459. [Google Scholar] [CrossRef] [Green Version]
- Briand, E.; Humbert, J.; Tambosco, K.; Bormans, M.; Gerwick, W.H. Role of bacteria in the production and degradation of Microcystis cyanopeptides. Microbiologyopen 2016, 5, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Woodhouse, J.N.; Kinsela, A.S.; Collins, R.N.; Bowling, L.C.; Honeyman, G.L.; Holliday, J.K.; Neilan, B.A. Microbial communities reflect temporal changes in cyanobacterial composition in a shallow ephemeral freshwater lake. ISME J. 2016, 10, 1337–1351. [Google Scholar] [CrossRef] [PubMed]
- Guedes, I.A.; Rachid, C.T.C.C.; Rangel, L.M.; Silva, L.H.S.; Bisch, P.M.; Azevedo, S.M.F.O.; Pacheco, A.B.F. Close Link between harmful cyanobacterial dominance and associated bacterioplankton in a tropical eutrophic reservoir. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Dziallas, C.; Grossart, H.-P. Temperature and biotic factors influence bacterial communities associated with the cyanobacterium Microcystis sp. Environ. Microbiol. 2011, 13, 1632–1641. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Lu, T.; Song, H.; Lavoie, M.; Xu, J.; Fan, X.; Pan, X. Spatial variability of cyanobacteria and heterotrophic bacteria in lake Taihu (China). Bull. Environ. Contam. Toxicol. 2017, 99, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Jin, L.; Ko, S.-R.; Chun, S.-J.; Oh, H.-S.; Lee, C.S.; Srivastava, A.; Oh, H.-M.; Ahn, C.-Y. Periphyton effects on bacterial assemblages and harmful cyanobacterial blooms in a eutrophic freshwater lake: A mesocosm study. Sci. Rep. 2017, 7, 7827. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lake | Total Phytoplankton Biomass [mg L−1] | Cyanobacteria Biomass [mg L−1] | Cyanobacteria Biomass [%] | Planktothrix agardhii Biomass [%] | Raphidiopsis raciborskii Biomass [%] |
|---|---|---|---|---|---|
| CyanoDominant Lakes | |||||
| Kierskie Małe | 11.4 | 6.5 | 56.9 | 0 | 8.3 |
| Zbąszyńskie | 16.9 | 13.0 | 76.8 | 73.9 | 0 |
| Niepruszewskie | 33.7 | 25.9 | 76.8 | 1.3 | 1.5 |
| Bnińskie | 11.4 | 6.7 | 58.8 | 46.6 | 2.2 |
| Raczyńskie | 41.8 | 39.2 | 93.7 | 87.5 | 0.07 |
| Grzymisławskie | 2.3 | 1.1 | 51.1 | 0 | 5.0 |
| Dolskie Wielkie | 36.0 | 33.7 | 93.3 | 0 | 30.1 |
| Żnińskie Małe | 30.4 | 26.7 | 87.7 | 0.4 | 0.6 |
| Mogileńskie | 15.2 | 14.3 | 94.3 | 82.2 | 0 |
| Miejskie | 33.6 | 29.9 | 88.9 | 71.9 | 4.3 |
| Pniewskie | 7.1 | 4.3 | 61.1 | 1.7 | 5.0 |
| Bytyńskie | 39.5 | 37.0 | 93.5 | 7.1 | 0 |
| Swarzędzkie | 14.5 | 9.3 | 64.4 | 7.8 | 0.2 |
| Strzyżewskie | 16.0 | 12.9 | 78.5 | 3.0 | 0 |
| Biskupińskie | 55.3 | 50.5 | 91.4 | 1.2 | 0.05 |
| Grylewskie | 2.9 | 2.2 | 75.4 | 11.7 | 0.6 |
| CyanoMinor Lakes | |||||
| Buszewskie | 3.2 | 1.3 | 40.6 | 8.6 | 0.8 |
| Lubieckie | 1.3 | 0.2 | 16.7 | 0 | 0 |
| Borówie | 3.0 | 1.2 | 40.7 | 0 | 0 |
| Głębokie | 2.2 | 0.07 | 3.4 | 0 | 0 |
| Winiary | 8.5 | 3.7 | 41.9 | 0 | 0.3 |
| Rusałka | 5.5 | 0.8 | 15.2 | 0 | 0 |
| Lusowskie | 0.17 | 0.04 | 23.6 | 0 | 0 |
| Wolsztyńskie | 11.9 | 3.1 | 26.7 | 15.2 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kokociński, M.; Dziga, D.; Antosiak, A.; Soininen, J. Are Bacterio- and Phytoplankton Community Compositions Related in Lakes Differing in Their Cyanobacteria Contribution and Physico-Chemical Properties? Genes 2021, 12, 855. https://doi.org/10.3390/genes12060855
Kokociński M, Dziga D, Antosiak A, Soininen J. Are Bacterio- and Phytoplankton Community Compositions Related in Lakes Differing in Their Cyanobacteria Contribution and Physico-Chemical Properties? Genes. 2021; 12(6):855. https://doi.org/10.3390/genes12060855
Chicago/Turabian StyleKokociński, Mikołaj, Dariusz Dziga, Adam Antosiak, and Janne Soininen. 2021. "Are Bacterio- and Phytoplankton Community Compositions Related in Lakes Differing in Their Cyanobacteria Contribution and Physico-Chemical Properties?" Genes 12, no. 6: 855. https://doi.org/10.3390/genes12060855
APA StyleKokociński, M., Dziga, D., Antosiak, A., & Soininen, J. (2021). Are Bacterio- and Phytoplankton Community Compositions Related in Lakes Differing in Their Cyanobacteria Contribution and Physico-Chemical Properties? Genes, 12(6), 855. https://doi.org/10.3390/genes12060855