
Disparity of Hepatocellular Carcinoma in Tumor Microenvironment-Related Genes and Infiltrating Immune Cells between Asian and Non-Asian Populations

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Acquisition
2.2. Definition of Differentially Expressed Genes (DEGs)
2.3. Protein–Protein Interaction (PPI) Network
2.4. Immune Cell Profiling
2.5. Survival Analysis
2.6. Gene Ontology and Kyoto Encyclopedia of Genes and Genomes (KEGG) Enrichment Analysis
3. Results
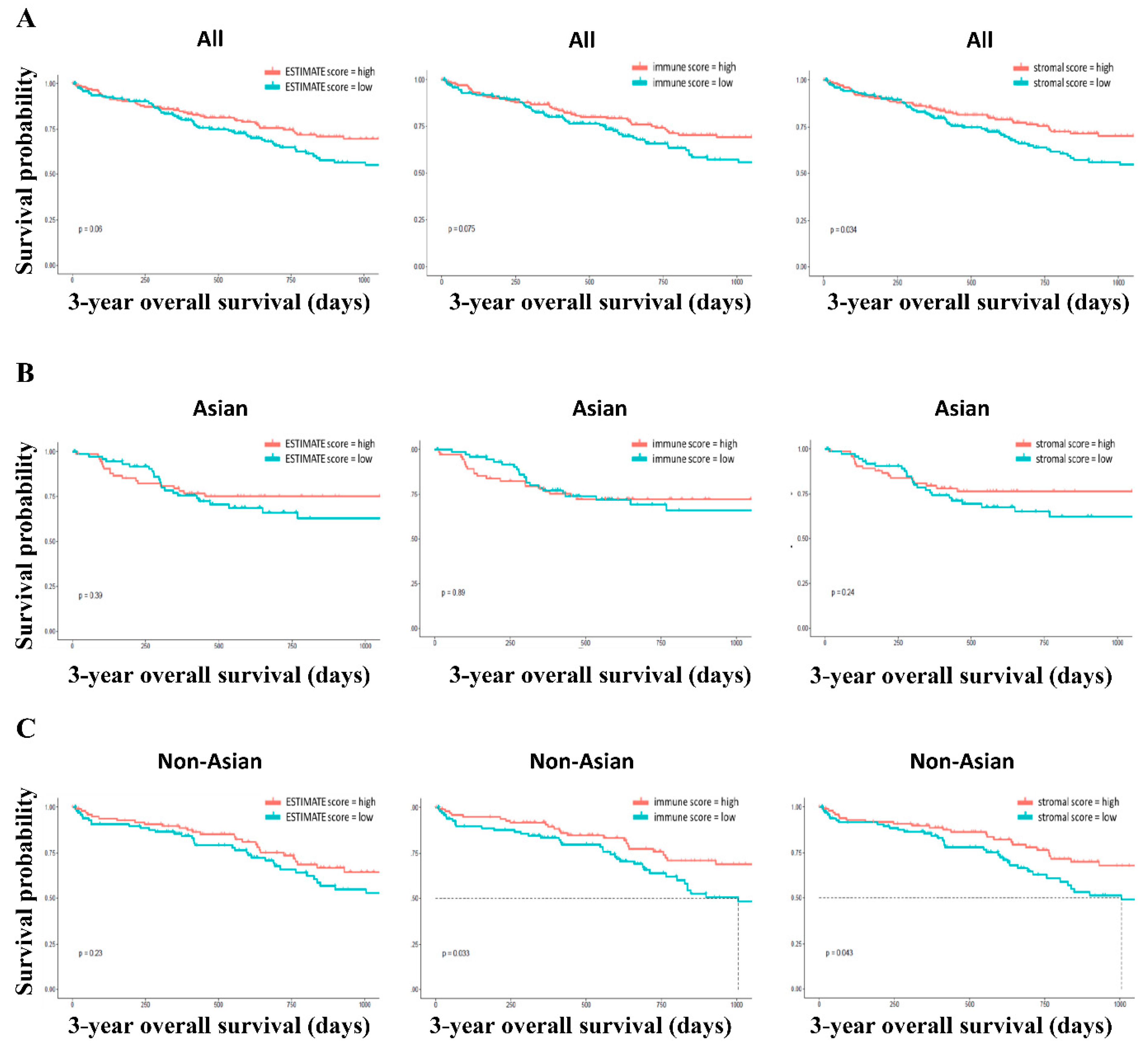
3.1. Study Patients’ Demographics and Kaplan–Meier Survival Analysis
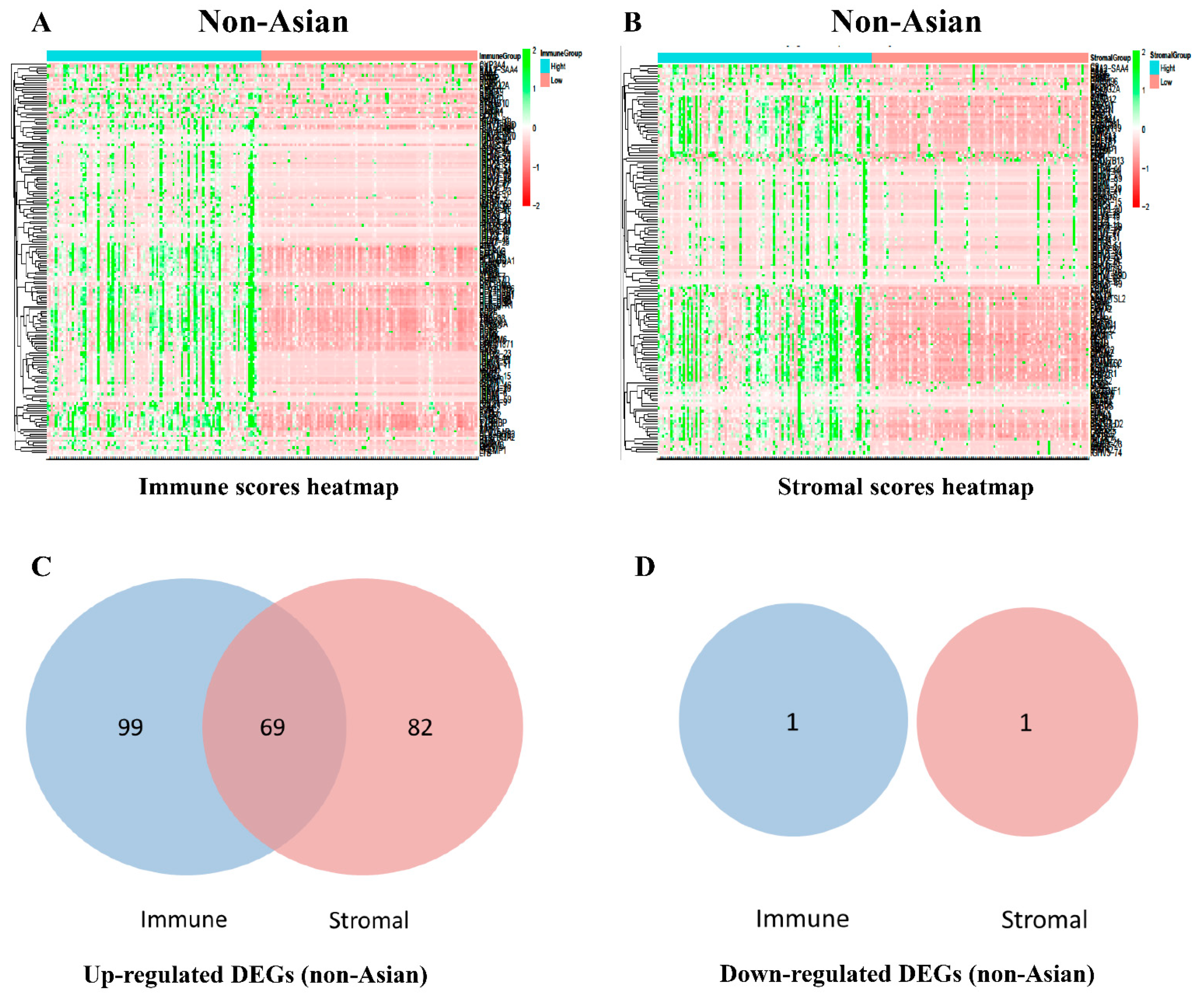
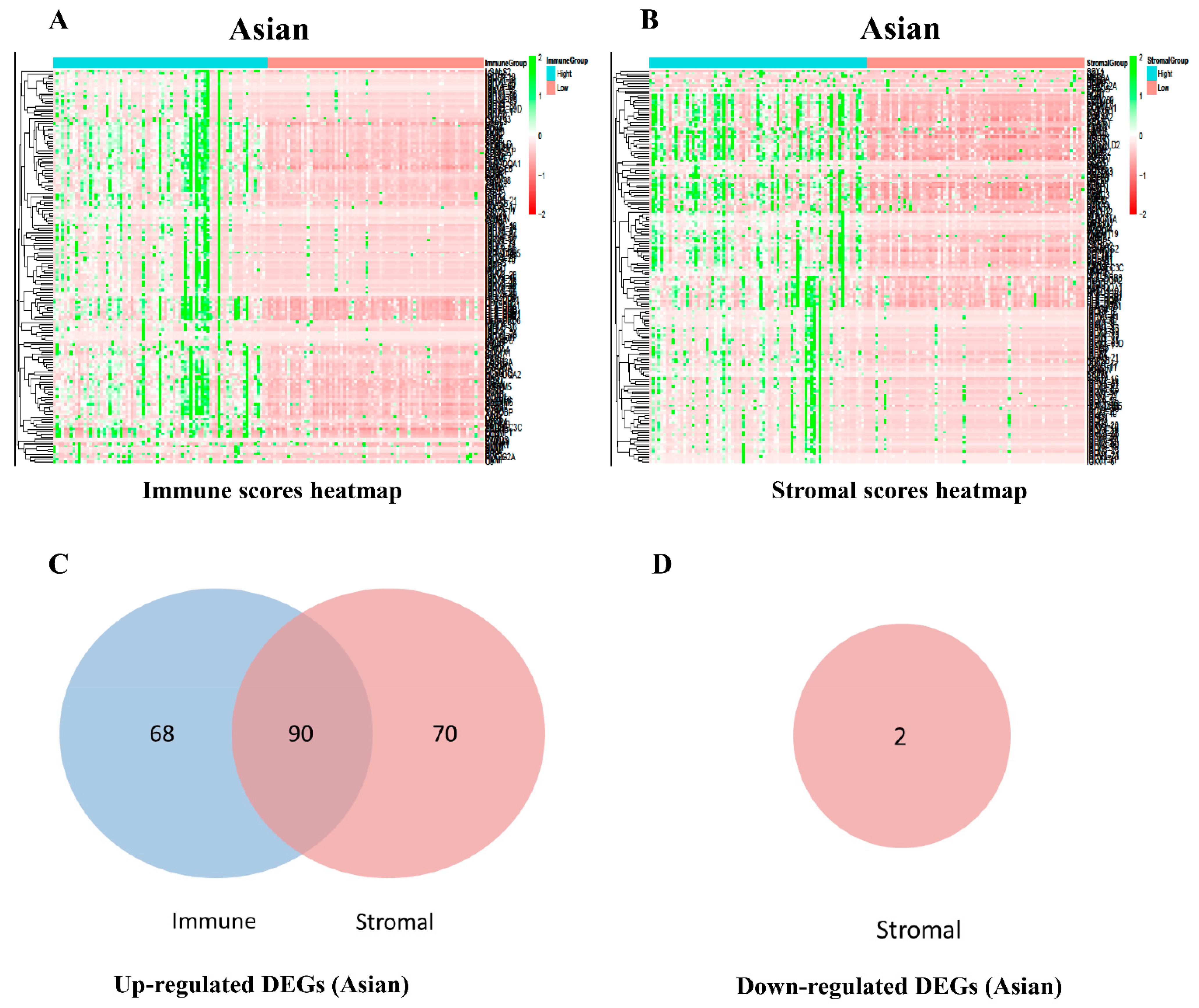
3.2. Screening of DEGs
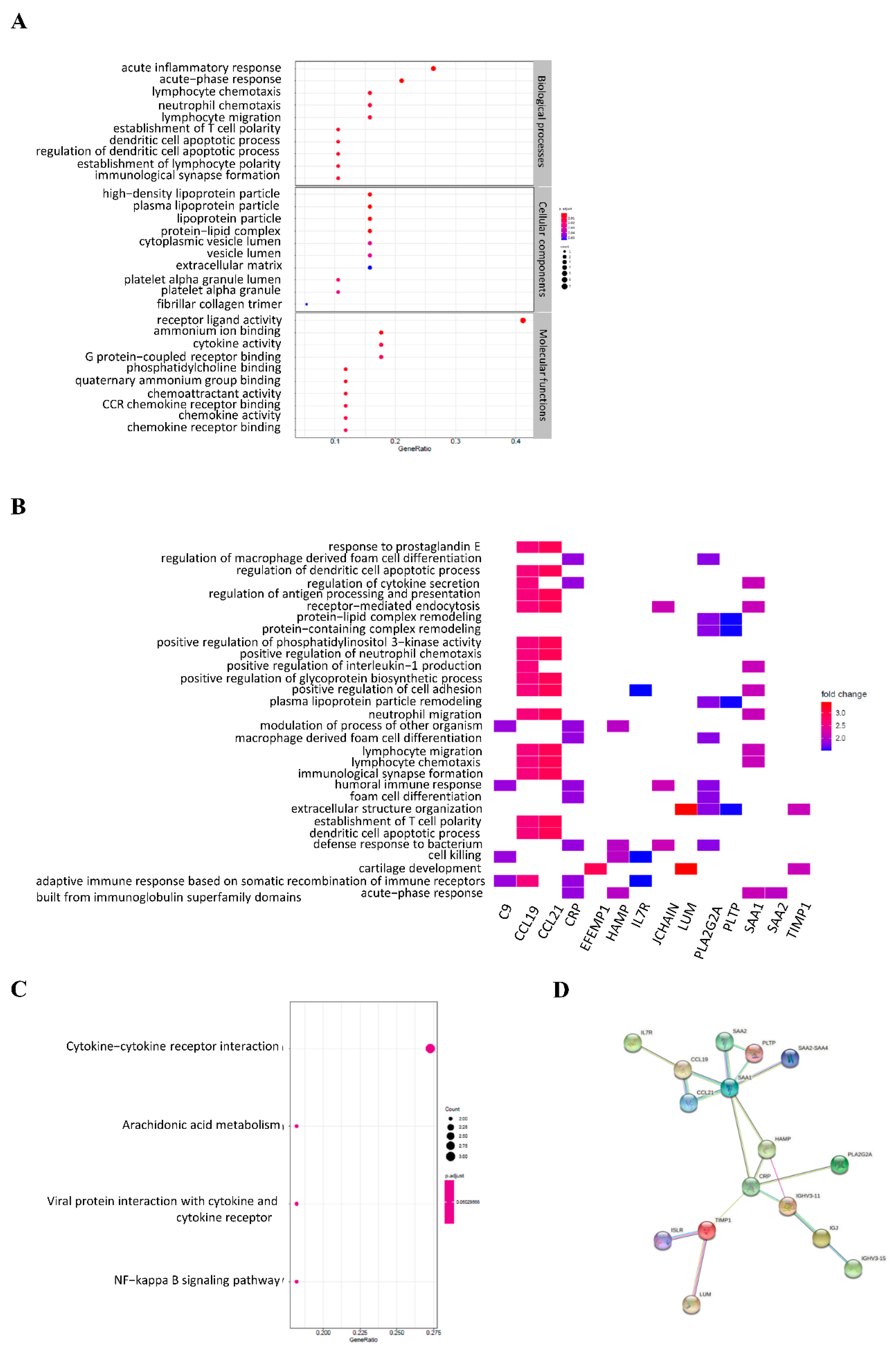
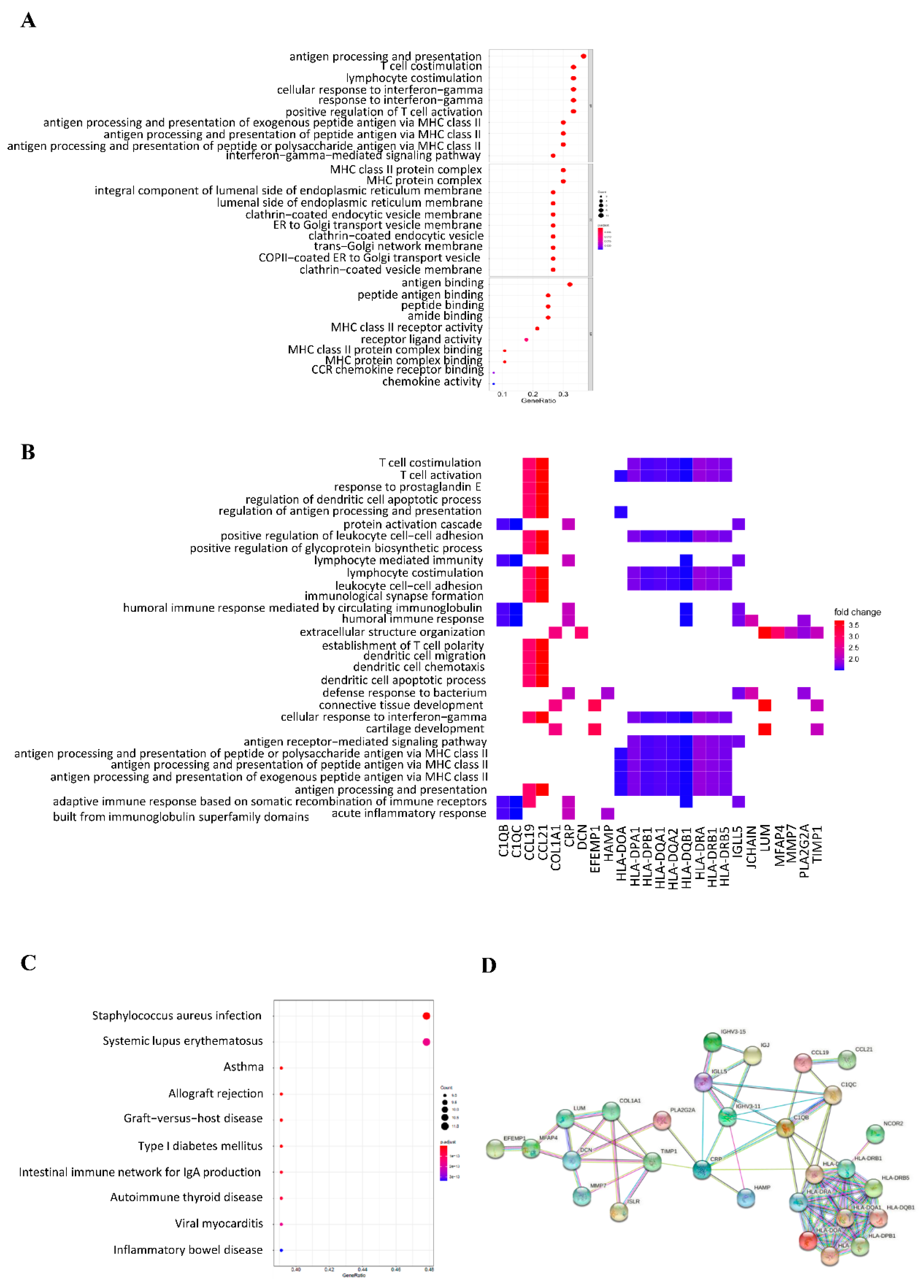
3.3. Gene Ontology and KEGG Enrichment Analysis of DEGs
3.4. PPI Network of DEGs
3.5. Hub Genes in HCC
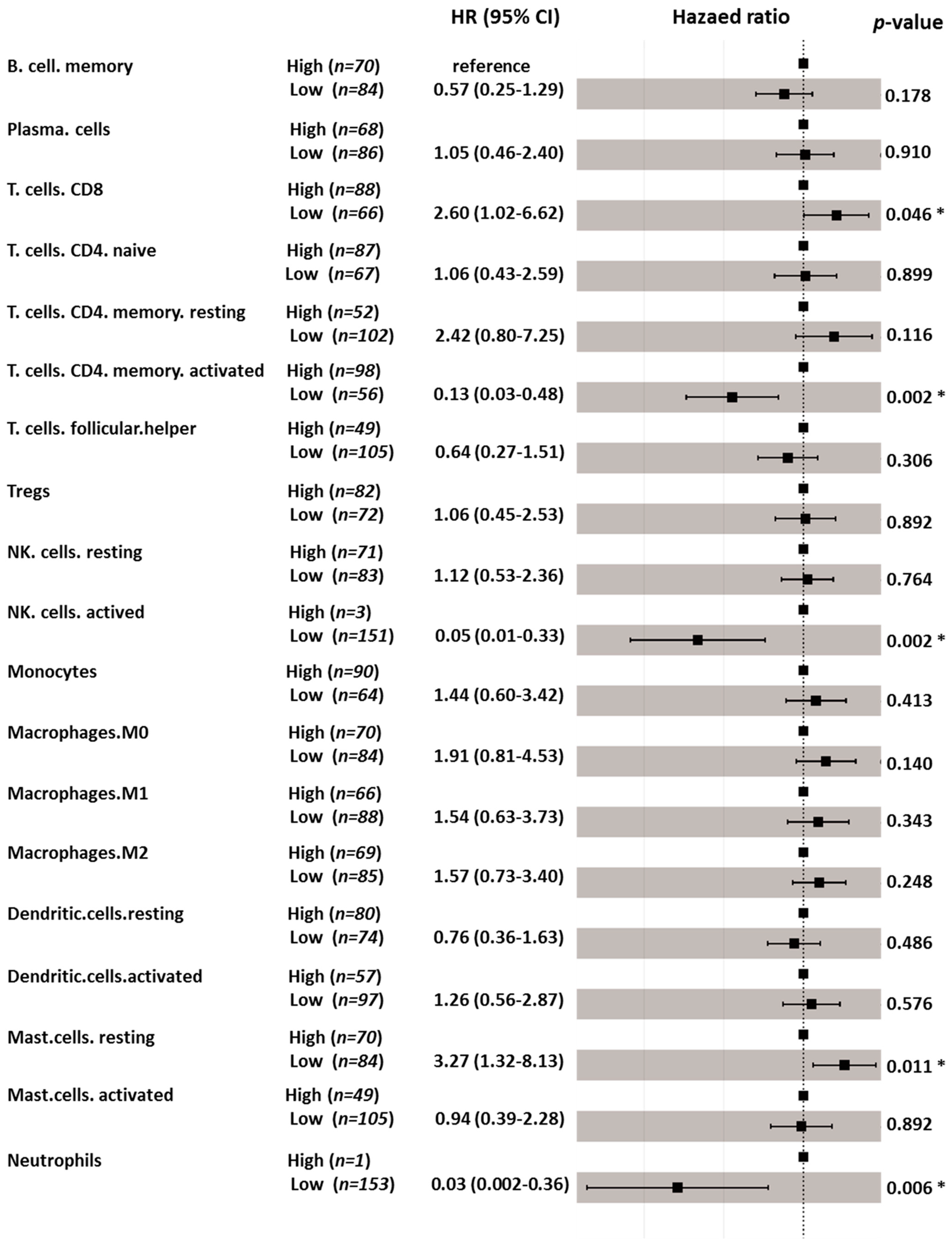
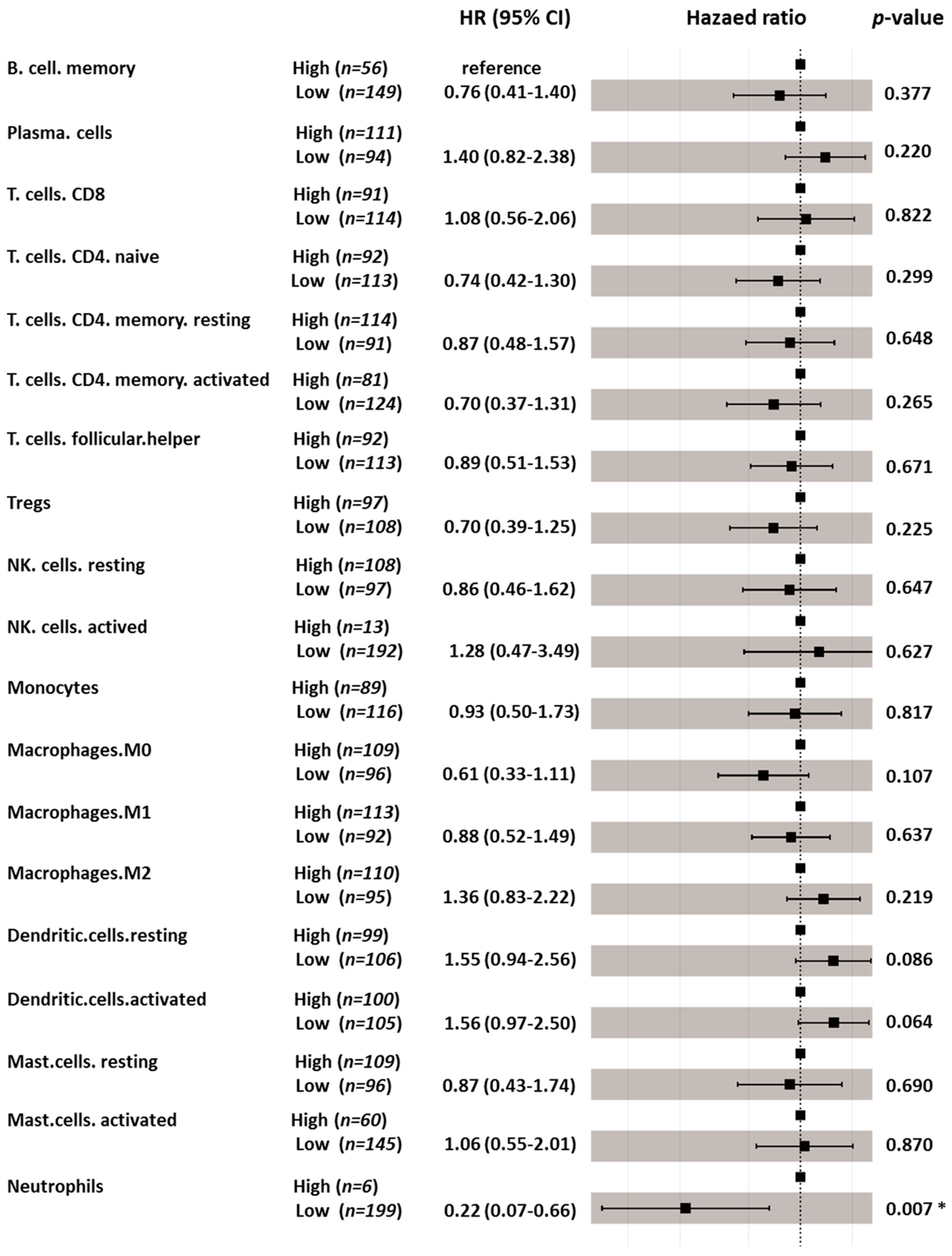
3.6. Immune Cell Profiling and Prognostic Effects
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sia, D.; Villanueva, A.; Friedman, S.L.; Llovet, J.M. Liver Cancer Cell of Origin, Molecular Class, and Effects on Patient Prognosis. Gastroenterology 2017, 152, 745–761. [Google Scholar] [CrossRef] [Green Version]
- Llovet, J.M.; Zucman-Rossi, J.; Pikarsky, E.; Sangro, B.; Schwartz, M.; Sherman, M.; Gores, G. Hepatocellular carcinoma. Nat. Rev. Dis. Primers 2016, 2, 16018. [Google Scholar] [CrossRef]
- Capece, D.; Fischietti, M.; Verzella, D.; Gaggiano, A.; Cicciarelli, G.; Tessitore, A.; Zazzeroni, F.; Alesse, E. The inflammatory microenvironment in hepatocellular carcinoma: A pivotal role for tumor-associated macrophages. BioMed Res. Int. 2013, 2013, 187204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omer, R.E.; Kuijsten, A.; Kadaru, A.M.; Kok, F.J.; Idris, M.O.; El Khidir, I.M.; van’t Veer, P. Population-attributable risk of dietary aflatoxins and hepatitis B virus infection with respect to hepatocellular carcinoma. Nutr. Cancer 2004, 48, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, X.; Gong, G.; Ben, Q.; Qiu, W.; Chen, Y.; Li, G.; Wang, L. Increased risk of hepatocellular carcinoma in patients with diabetes mellitus: A systematic review and meta-analysis of cohort studies. Int. J. Cancer 2012, 130, 1639–1648. [Google Scholar] [CrossRef]
- Zucman-Rossi, J.; Villanueva, A.; Nault, J.C.; Llovet, J.M. Genetic Landscape and Biomarkers of Hepatocellular Carcinoma. Gastroenterology 2015, 149, 1226–1239.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.D.; Ma, Y.S.; Fang, Y.; Liu, L.L.; Fu, D.; Shen, X.Z. Role of the microenvironment in hepatocellular carcinoma development and progression. Cancer Treat. Rev. 2012, 38, 218–225. [Google Scholar] [CrossRef]
- Yang, J.D.; Nakamura, I.; Roberts, L.R. The tumor microenvironment in hepatocellular carcinoma: Current status and therapeutic targets. Semin. Cancer Biol. 2011, 21, 35–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, H.R. Roles of Tumor Microenvironment in Hepatocelluar Carcinoma. Curr. Med. Chem. 2015, 11, 82–93. [Google Scholar]
- Shiga, K.; Hara, M.; Nagasaki, T.; Sato, T.; Takahashi, H.; Takeyama, H. Cancer-Associated Fibroblasts: Their Characteristics and Their Roles in Tumor Growth. Cancers 2015, 7, 2443–2458. [Google Scholar] [CrossRef]
- Birgani, M.T.; Carloni, V. Tumor Microenvironment, a Paradigm in Hepatocellular Carcinoma Progression and Therapy. Int. J. Mol. Sci. 2017, 18, 405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalluri, R.; Zeisberg, M. Fibroblasts in cancer. Nat. Rev. Cancer 2006, 6, 392–401. [Google Scholar] [CrossRef]
- Wynn, T.A. Cellular and molecular mechanisms of fibrosis. J. Pathol. 2008, 214, 199–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, U.E.; Friedman, S.L. Mechanisms of hepatic fibrogenesis. Best Pract. Res. Clin. Gastroenterol. 2011, 25, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Zhang, L.; Xu, Y.; Zhang, Z.; Ren, G.; Tang, K.; Kuang, P.; Zhao, B.; Yin, Z.; Wang, X. Hepatic stellate cells promote tumor progression by enhancement of immunosuppressive cells in an orthotopic liver tumor mouse model. Lab. Investig. J. Tech. Methods Pathol. 2014, 94, 182–191. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Zhao, W.; Xu, J.; Li, J.; Hong, Z.; Yin, Z.; Wang, X. Activated hepatic stellate cells promote liver cancer by induction of myeloid-derived suppressor cells through cyclooxygenase-2. Oncotarget 2016, 7, 8866–8878. [Google Scholar] [CrossRef] [Green Version]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, inflammation, and cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [Green Version]
- Solinas, G.; Germano, G.; Mantovani, A.; Allavena, P. Tumor-associated macrophages (TAM) as major players of the cancer-related inflammation. J. Leukoc. Biol. 2009, 86, 1065–1073. [Google Scholar] [CrossRef] [Green Version]
- Lewis, C.E.; Pollard, J.W. Distinct role of macrophages in different tumor microenvironments. Cancer Res. 2006, 66, 605–612. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Xu, D.; Liu, Z.; Shi, M.; Zhao, P.; Fu, B.; Zhang, Z.; Yang, H.; Zhang, H.; Zhou, C.; et al. Increased regulatory T cells correlate with CD8 T-cell impairment and poor survival in hepatocellular carcinoma patients. Gastroenterology 2007, 132, 2328–2339. [Google Scholar] [CrossRef]
- Yoshihara, K.; Shahmoradgoli, M.; Martinez, E.; Vegesna, R.; Kim, H.; Torres-Garcia, W.; Trevino, V.; Shen, H.; Laird, P.W.; Levine, D.A.; et al. Inferring tumour purity and stromal and immune cell admixture from expression data. Nat. Commun. 2013, 4, 2612. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhao, K.; Kang, H.; Wang, M.; Wu, A. Exploring immune-related genes with prognostic value in microenvironment of breast cancer from TCGA database. Medicine 2020, 99, e19561. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.H.; Xu, Y.; Wang, J.; Wan, F.N.; Wang, H.K.; Cao, D.L.; Shi, G.H.; Qu, Y.Y.; Zhang, H.L.; Ye, D.W. Prognostic value and immune infiltration of novel signatures in clear cell renal cell carcinoma microenvironment. Aging 2019, 11, 6999–7020. [Google Scholar] [CrossRef] [PubMed]
- Mao, M.; Yu, Q.; Huang, R.; Lu, Y.; Wang, Z.; Liao, L. Stromal score as a prognostic factor in primary gastric cancer and close association with tumor immune microenvironment. Cancer Med. 2020, 9, 4980–4990. [Google Scholar] [CrossRef]
- Qi, X.; Qi, C.; Qin, B.; Kang, X.; Hu, Y.; Han, W. Immune-Stromal Score Signature: Novel Prognostic Tool of the Tumor Microenvironment in Lung Adenocarcinoma. Front. Oncol. 2020, 10, 541330. [Google Scholar] [CrossRef] [PubMed]
- Ge, P.L.; Li, S.F.; Wang, W.W.; Li, C.B.; Fu, Y.B.; Feng, Z.K.; Li, L.; Zhang, G.; Gao, Z.Q.; Dang, X.W.; et al. Prognostic values of immune scores and immune microenvironment-related genes for hepatocellular carcinoma. Aging 2020, 12, 5479–5499. [Google Scholar] [CrossRef]
- Ferlay, J.; Shin, H.R.; Bray, F.; Forman, D.; Mathers, C.; Parkin, D.M. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int. J. Cancer 2010, 127, 2893–2917. [Google Scholar] [CrossRef]
- McGlynn, K.A.; Petrick, J.L.; El-Serag, H.B. Epidemiology of Hepatocellular Carcinoma. Hepatology 2021, 73 (Suppl. 1), 4–13. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, A. Hepatocellular Carcinoma. N. Engl. J. Med. 2019, 380, 1450–1462. [Google Scholar] [CrossRef] [Green Version]
- Choo, S.P.; Tan, W.L.; Goh, B.K.P.; Tai, W.M.; Zhu, A.X. Comparison of hepatocellular carcinoma in Eastern versus Western populations. Cancer 2016, 122, 3430–3446. [Google Scholar] [CrossRef]
- Balogh, J.; Victor, D., 3rd; Asham, E.H.; Burroughs, S.G.; Boktour, M.; Saharia, A.; Li, X.; Ghobrial, R.M.; Monsour, H.P., Jr. Hepatocellular carcinoma: A review. J. Hepatocell. Carcinoma 2016, 3, 41–53. [Google Scholar] [CrossRef] [Green Version]
- El-Serag, H.B.; Kanwal, F. Epidemiology of hepatocellular carcinoma in the United States: Where are we? Where do we go? Hepatology 2014, 60, 1767–1775. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; El-Serag, H.B. The Epidemiology of Hepatocellular Carcinoma in the USA. Curr. Gastroenterol. Rep. 2019, 21, 17. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.H.; Wang, J.B.; Jiang, Y.; Xiang, W.; Liang, H.; Wei, W.Q.; Qiao, Y.L.; Boffetta, P. Attributable causes of liver cancer mortality and incidence in china. Asian Pac. J. Cancer Prev. 2013, 14, 7251–7256. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Kumar, R.; Hissar, S.S.; Saraswat, M.K.; Sharma, B.C.; Sakhuja, P.; Sarin, S.K. Risk factors analysis for hepatocellular carcinoma in patients with and without cirrhosis: A case-control study of 213 hepatocellular carcinoma patients from India. J. Gastroenterol. Hepatol. 2007, 22, 1104–1111. [Google Scholar] [CrossRef]
- Acharya, S.K. Epidemiology of hepatocellular carcinoma in India. J. Clin. Exp. Hepatol. 2014, 4, S27–S33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Niu, X.; Qiu, Z. A Five-Gene Signature Based on Stromal/Immune Scores in the Tumor Microenvironment and Its Clinical Implications for Liver Cancer. DNA Cell Biol. 2020, 39, 1621–1638. [Google Scholar] [CrossRef]
- Hu, J.; Gao, D.Z. Distinction immune genes of hepatitis-induced heptatocellular carcinoma. Bioinformatics 2012, 28, 3191–3194. [Google Scholar] [CrossRef] [Green Version]
- Nieters, A.; Yuan, J.M.; Sun, C.L.; Zhang, Z.Q.; Stoehlmacher, J.; Govindarajan, S.; Yu, M.C. Effect of cytokine genotypes on the hepatitis B virus-hepatocellular carcinoma association. Cancer 2005, 103, 740–748. [Google Scholar] [CrossRef]
- Ognjanovic, S.; Yuan, J.M.; Chaptman, A.K.; Fan, Y.; Yu, M.C. Genetic polymorphisms in the cytokine genes and risk of hepatocellular carcinoma in low-risk non-Asians of USA. Carcinogenesis 2009, 30, 758–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.X.; Wang, J.L.; Guo, X.J.; He, C.C.; Ying, X.; Ma, J.L.; Zhang, Y.Y.; Zhao, Q.; Zhu, Q. Serum SALL4 is a novel prognosis biomarker with tumor recurrence and poor survival of patients in hepatocellular carcinoma. J. Immunol. Res. 2014, 2014, 262385. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.C.; Vachharajani, N.; Chapman, W.C.; Brunt, E.M. SALL4 immunoreactivity predicts prognosis in Western hepatocellular carcinoma patients but is a rare event: A study of 236 cases. Am. J. Surg. Pathol. 2014, 38, 966–972. [Google Scholar] [CrossRef] [PubMed]
- Binnewies, M.; Roberts, E.W.; Kersten, K.; Chan, V.; Fearon, D.F.; Merad, M.; Coussens, L.M.; Gabrilovich, D.I.; Ostrand-Rosenberg, S.; Hedrick, C.C.; et al. Understanding the tumor immune microenvironment (TIME) for effective therapy. Nat. Med. 2018, 24, 541–550. [Google Scholar] [CrossRef]
- Estes, C.; Anstee, Q.M.; Arias-Loste, M.T.; Bantel, H.; Bellentani, S.; Caballeria, J.; Colombo, M.; Craxi, A.; Crespo, J.; Day, C.P.; et al. Modeling NAFLD disease burden in China, France, Germany, Italy, Japan, Spain, United Kingdom, and United States for the period 2016–2030. J. Hepatol. 2018, 69, 896–904. [Google Scholar] [CrossRef]
- Suresh, D.; Srinivas, A.N.; Kumar, D.P. Etiology of Hepatocellular Carcinoma: Special Focus on Fatty Liver Disease. Front. Oncol. 2020, 10, 601710. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Asian | Non-Asian | p-Value | |||
|---|---|---|---|---|---|
| (n = 205) | (n = 154) | ||||
| Age (years) | 55.42 ± 11.27 | 62.92 ± 13.65 | <0.001 | ||
| Body weight (kg) | 62.59 ± 10.56 | 81.52 ± 20.92 | <0.001 | ||
| Gender | <0.001 | ||||
| Male | 121 | 78.60% | 120 | 58.50% | |
| Female | 33 | 21.40% | 85 | 41.50% | |
| Stage | 0.498 | ||||
| Stage I | 79 | 51.30% | 88 | 42.90% | |
| Stage II | 34 | 22.10% | 50 | 24.40% | |
| Stage III | 38 | 24.70% | 44 | 21.50% | |
| Stage IV | 1 | 0.60% | 4 | 2.00% | |
| NA | 2 | 1.30% | 19 | 9.30% | |
| Survival status | 0.008 | ||||
| Alive | 119 | 58.00% | 111 | 72.10% | |
| Dead | 86 | 42.00% | 43 | 27.90% | |
| Survival Time (months) | 26.00 ± 22.76 | 27.43 ± 24.84 | |||
| p-Value | |||
|---|---|---|---|
| Gene | All (n = 359) | Asian (n = 205) | Non-Asian (n = 154) |
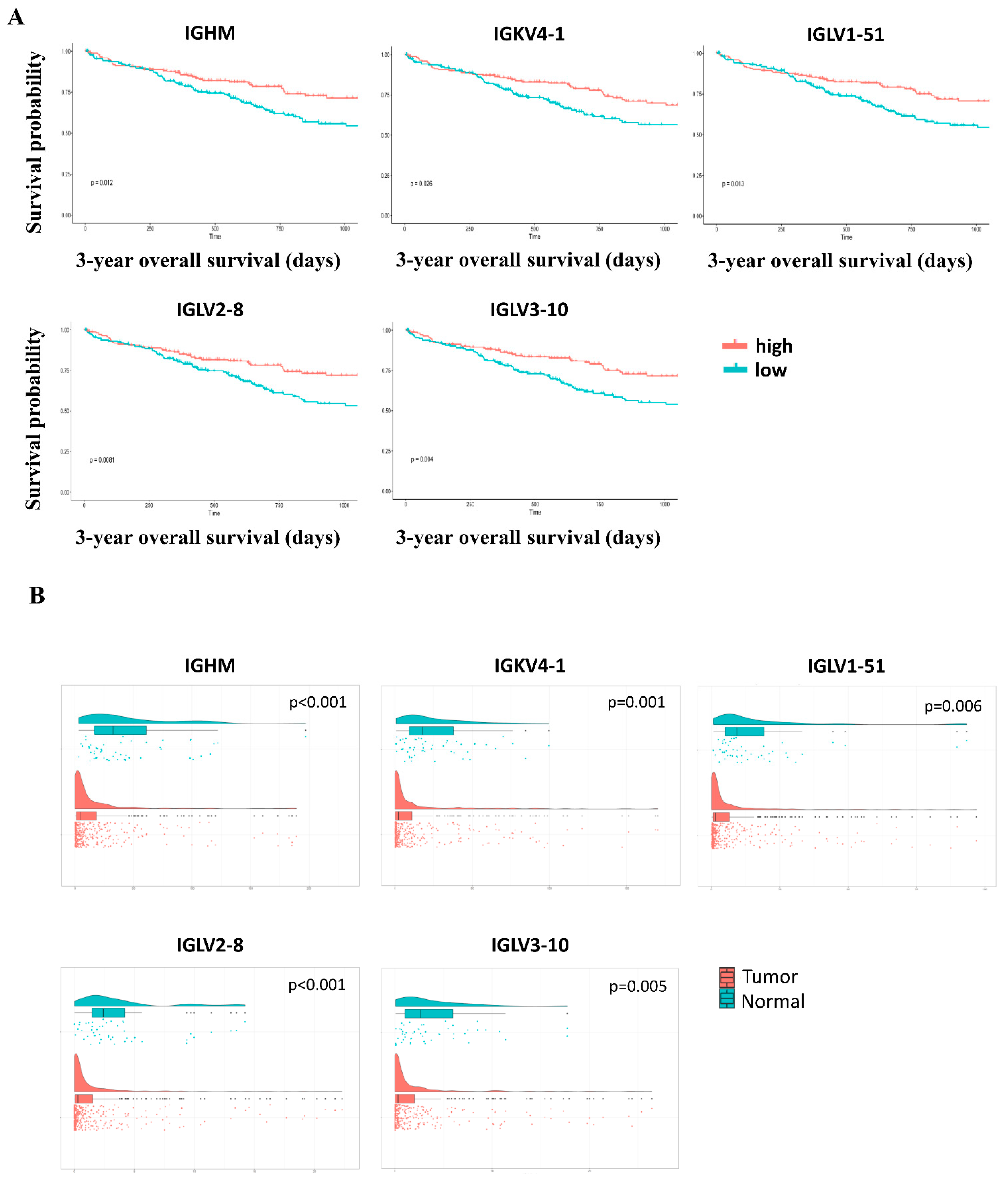
| IGHM | 0.012 | 0.263 | 0.015 |
| IGKV4-1 | 0.026 | 0.186 | 0.068 |
| IGLV1-51 | 0.013 | 0.440 | 0.020 |
| IGLV2-8 | 0.008 | 0.655 | 0.002 |
| IGLV3-10 | 0.004 | 0.127 | 0.018 |
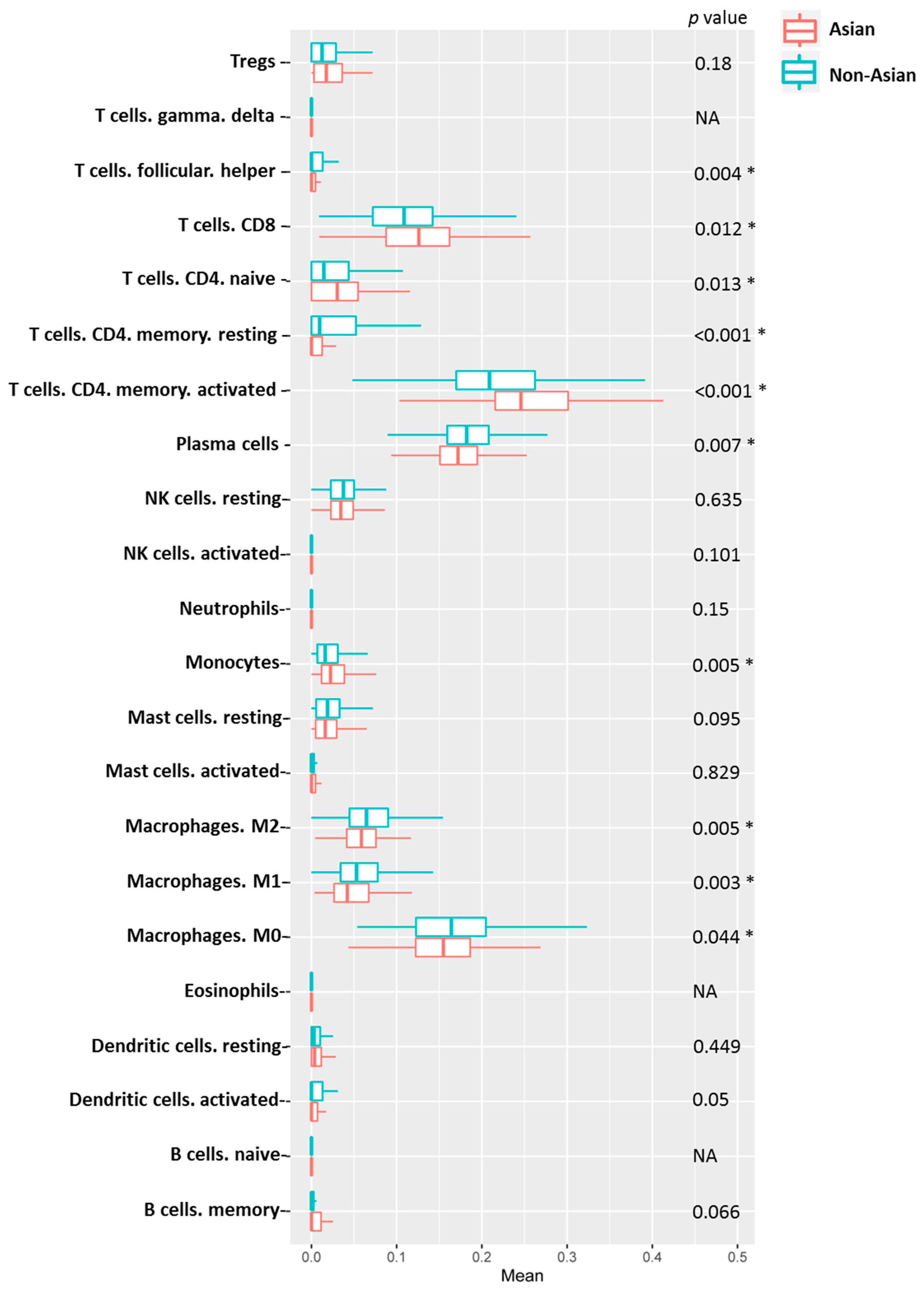
| p-Value | ||
|---|---|---|
| Asian | Non-Asian | |
| Naïve B cells | NA | NA |
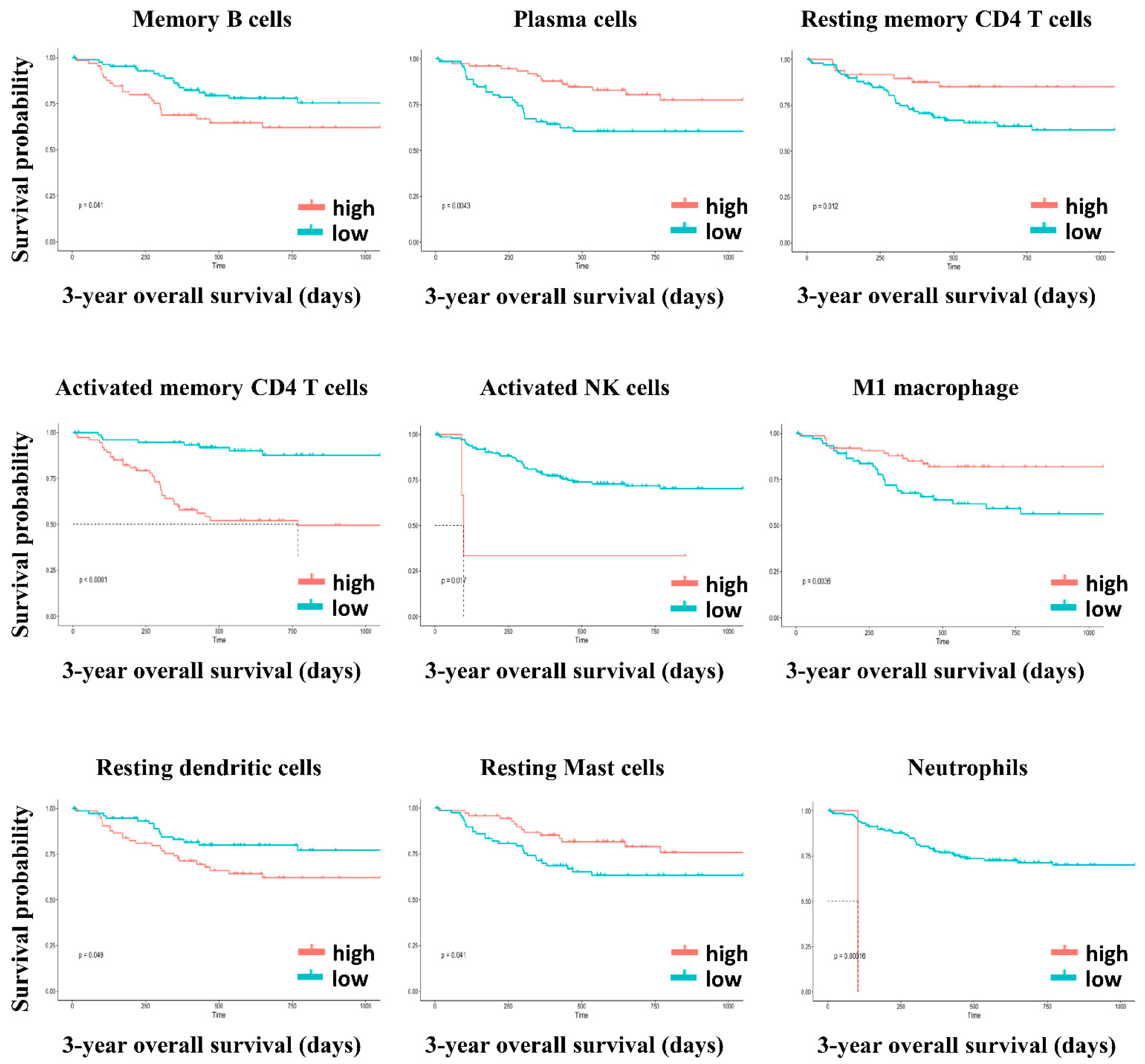
| Memory B cells | 0.041 | 0.574 |
| Plasma cells | 0.004 | 0.186 |
| CD8 T cells | 0.082 | 0.131 |
| CD4 naïve T cells | 0.816 | 0.136 |
| Resting memory CD4 T cells | 0.012 | 0.465 |
| Activated memory CD4 T cells | <0.001 | 0.115 |
| Follicular helper T cells | 0.940 | 0.066 |
| Tregs | 0.882 | 0.566 |
| Γ delta T cells | NA | NA |
| Resting NK cells | 0.296 | 0.370 |
| Activated NK cells | 0.017 | 0.107 |
| Monocytes | 0.515 | 0.135 |
| M0 Macrophages | 0.072 | 0.082 |
| M1 Macrophages | 0.004 | 0.564 |
| M2 Macrophages | 0.084 | 0.387 |
| Resting dendritic cells | 0.049 | 0.766 |
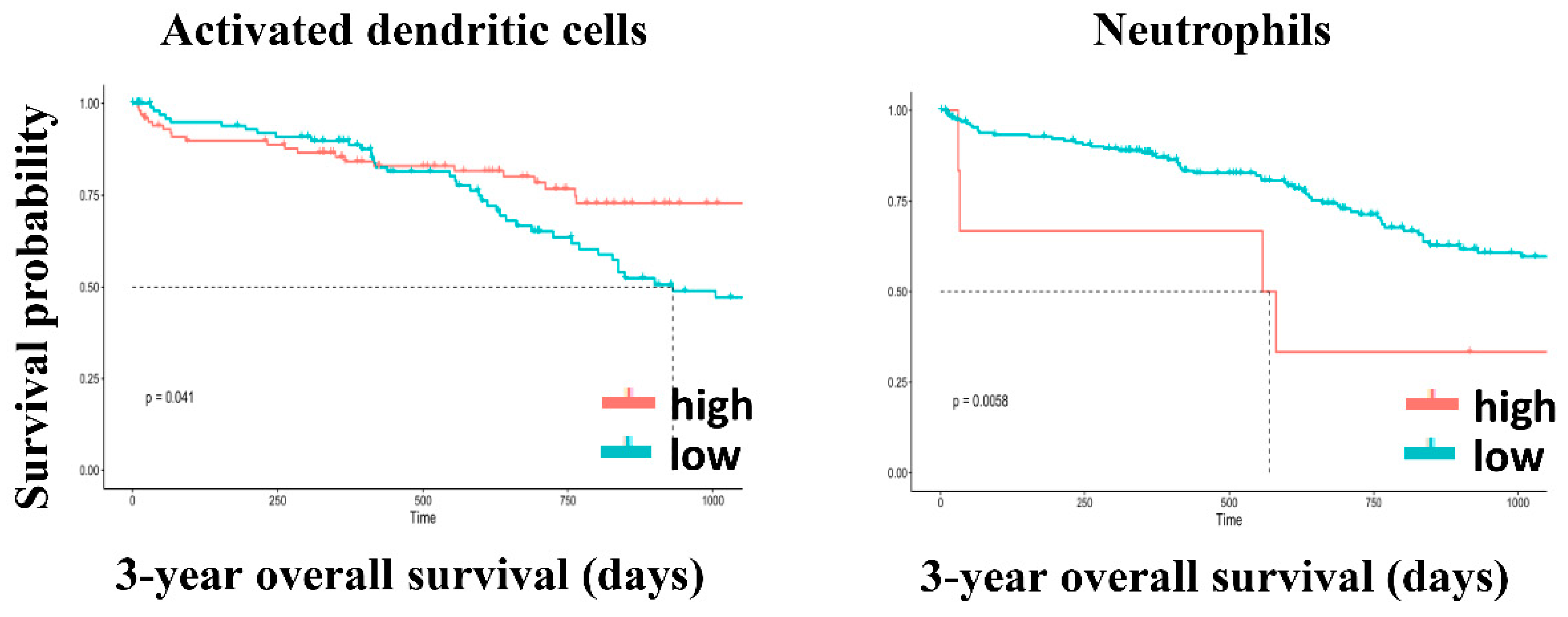
| Activated dendritic cells | 0.892 | 0.041 |
| Resting mast cells | 0.041 | 0.766 |
| Activated mast cells | 0.380 | 0.578 |
| Eosinophils | NA | NA |
| Neutrophils | <0.001 | 0.006 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, L.-H.; Hsieh, T.-M.; Huang, C.-Y.; Liu, Y.-W.; Wu, S.-C.; Chien, P.-C.; Hsieh, C.-H. Disparity of Hepatocellular Carcinoma in Tumor Microenvironment-Related Genes and Infiltrating Immune Cells between Asian and Non-Asian Populations. Genes 2021, 12, 1274. https://doi.org/10.3390/genes12081274
Huang L-H, Hsieh T-M, Huang C-Y, Liu Y-W, Wu S-C, Chien P-C, Hsieh C-H. Disparity of Hepatocellular Carcinoma in Tumor Microenvironment-Related Genes and Infiltrating Immune Cells between Asian and Non-Asian Populations. Genes. 2021; 12(8):1274. https://doi.org/10.3390/genes12081274
Chicago/Turabian StyleHuang, Lien-Hung, Ting-Min Hsieh, Chun-Ying Huang, Yueh-Wei Liu, Shao-Chun Wu, Peng-Chen Chien, and Ching-Hua Hsieh. 2021. "Disparity of Hepatocellular Carcinoma in Tumor Microenvironment-Related Genes and Infiltrating Immune Cells between Asian and Non-Asian Populations" Genes 12, no. 8: 1274. https://doi.org/10.3390/genes12081274
APA StyleHuang, L. -H., Hsieh, T. -M., Huang, C. -Y., Liu, Y. -W., Wu, S. -C., Chien, P. -C., & Hsieh, C. -H. (2021). Disparity of Hepatocellular Carcinoma in Tumor Microenvironment-Related Genes and Infiltrating Immune Cells between Asian and Non-Asian Populations. Genes, 12(8), 1274. https://doi.org/10.3390/genes12081274