
Identification of Immune Markers in Dilated Cardiomyopathies with Heart Failure by Integrated Weighted Gene Coexpression Network Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
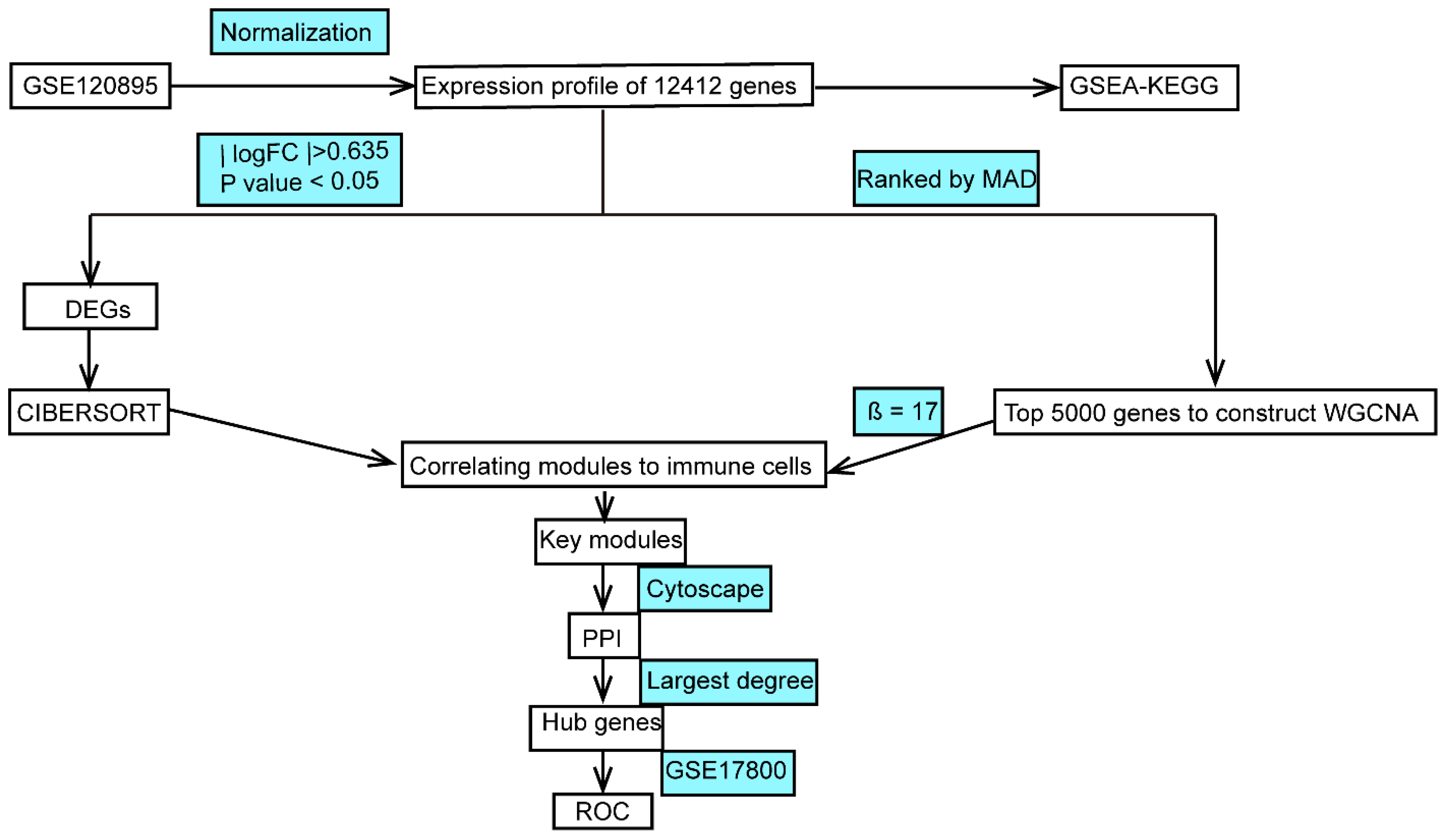
2.1. Data Processing and Analysis
2.2. Functional Enrichment Analysis
2.3. CIBERSORT
2.4. WGCNA
2.5. Protein–Protein Interaction (PPI) Network Analysis and Obtained Hub Genes
2.6. Validation of Hub Genes
3. Results
3.1. Identification of DEGs
3.2. Functional Correlation Analysis
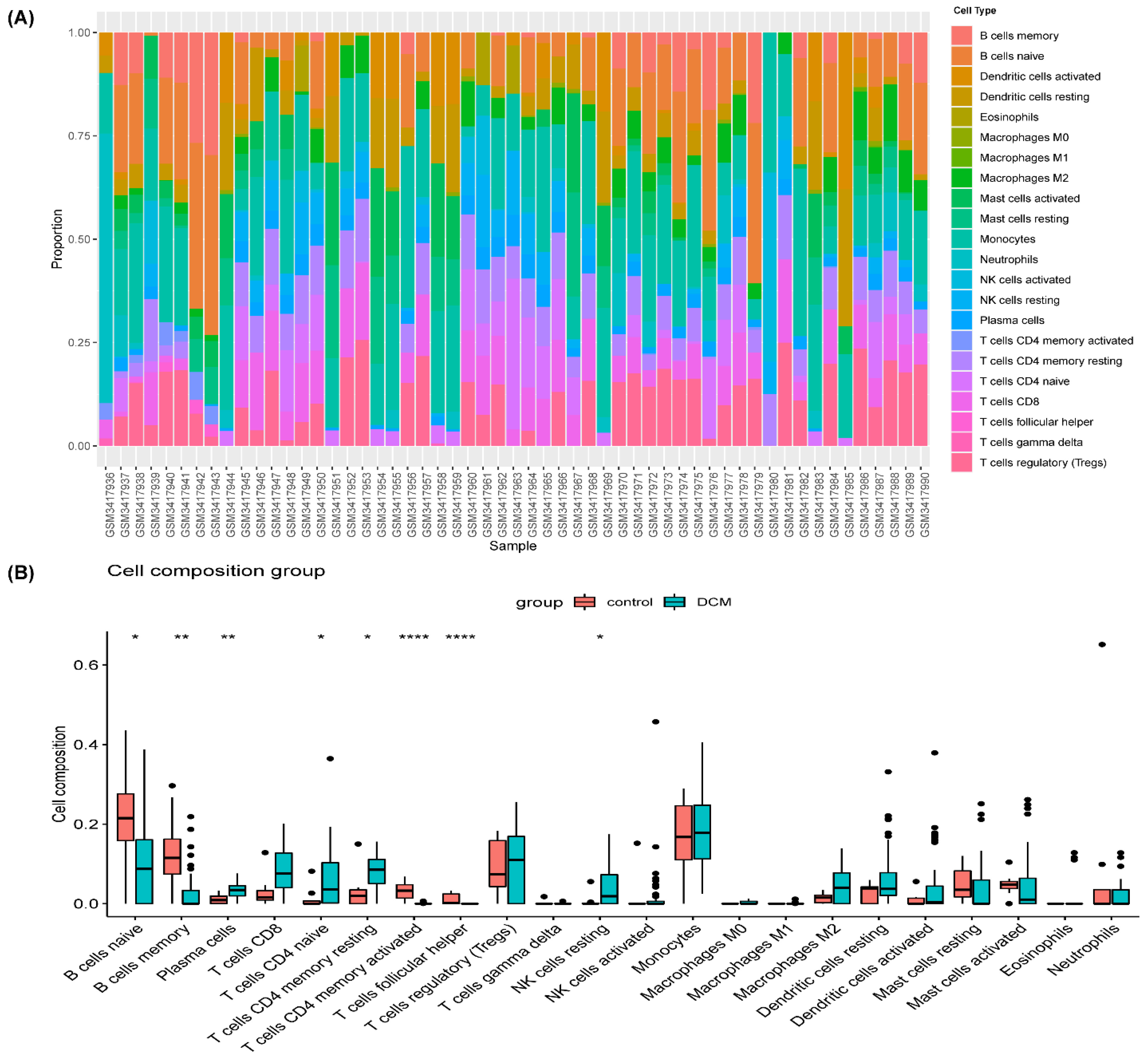
3.3. Immune Cell Infiltration
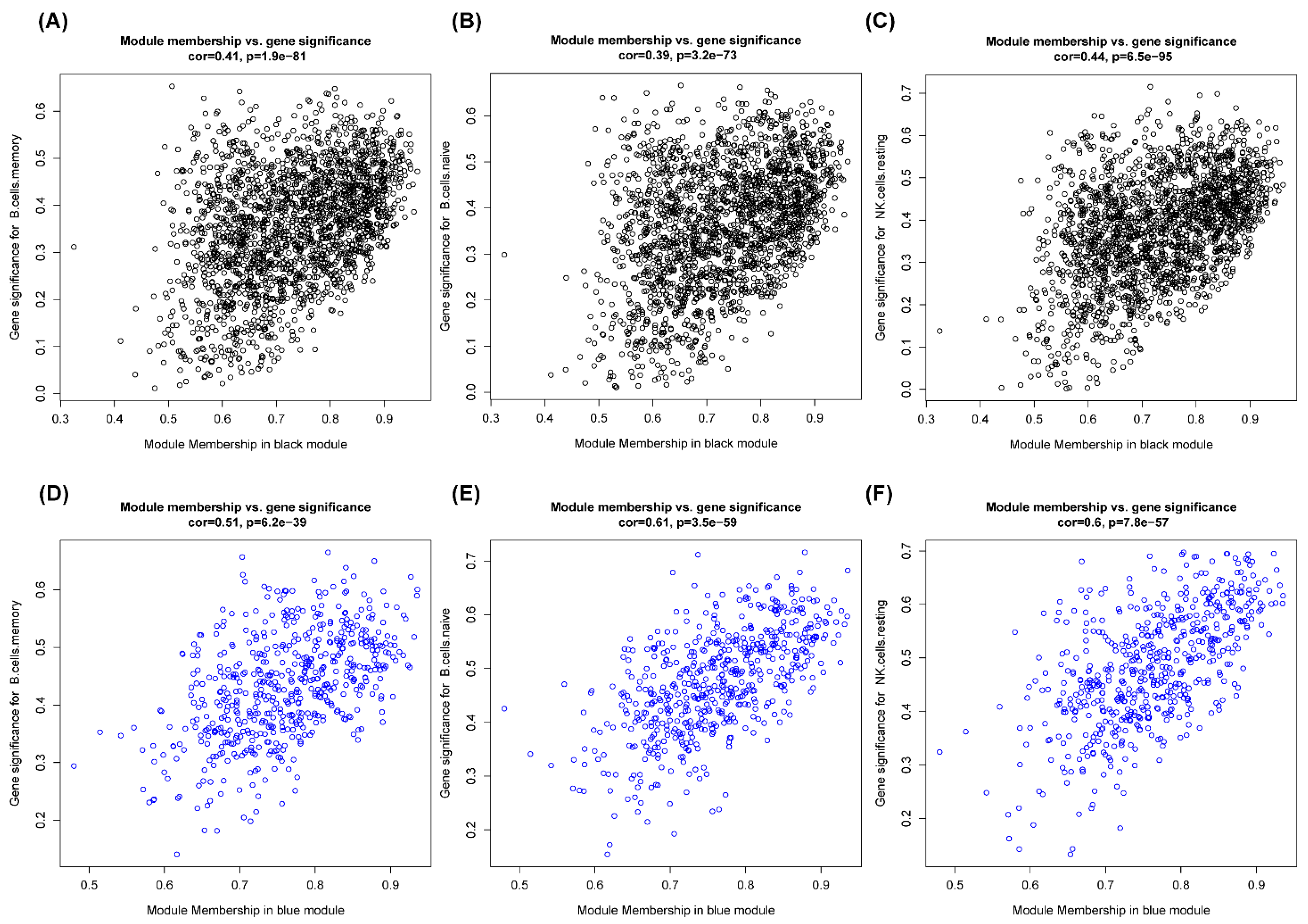
3.4. Construction of Weighted Coexpression Network and Identification of Immune-Corrected Modules
3.5. PPI Network Construction and Identification of Hub Genes
3.6. ROC Curve Analysis of Hub Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ponikowski, P.; Voors, A.A.; Anker, S.D.; Bueno, H.; Cleland, J.; Coats, A.; Falk, V.; Gonzalez-Juanatey, J.R.; Harjola, V.P.; Jankowska, E.A.; et al. 2016 ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure: The Task Force for the diagnosis and treatment of acute and chronic heart failure of the European Society of Cardiology (ESC)Developed with the special contribution of the Heart Failure Association (HFA) of the ESC. Eur. Heart J. 2016, 37, 2129–2200. [Google Scholar] [CrossRef] [PubMed]
- Groenewegen, A.; Rutten, F.H.; Mosterd, A.; Hoes, A.W. Epidemiology of heart failure. Eur. J. Heart Fail. 2020, 22, 1342–1356. [Google Scholar] [CrossRef] [PubMed]
- Roth, G.A.; Forouzanfar, M.H.; Moran, A.E.; Barber, R.; Nguyen, G.; Feigin, V.L.; Naghavi, M.; Mensah, G.A.; Murray, C.J. Demographic and epidemiologic drivers of global cardiovascular mortality. N. Engl. J. Med. 2015, 372, 1333–1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultheiss, H.P.; Fairweather, D.; Caforio, A.; Escher, F.; Hershberger, R.E.; Lipshultz, S.E.; Liu, P.P.; Matsumori, A.; Mazzanti, A.; Mcmurray, J.; et al. Dilated cardiomyopathy. Nat. Rev. Dis. Primers 2019, 5, 32. [Google Scholar] [CrossRef]
- Alimadadi, A.; Munroe, P.B.; Joe, B.; Cheng, X. Meta-analysis of dilated cardiomyopathy using cardiac RNA-seq transcriptomic datasets. Genes 2020, 11, 60. [Google Scholar] [CrossRef] [Green Version]
- Seferovic, P.M.; Polovina, M.; Bauersachs, J.; Arad, M.; Gal, T.B.; Lund, L.H.; Felix, S.B.; Arbustini, E.; Caforio, A.; Farmakis, D.; et al. Heart failure in cardiomyopathies: A position paper from the Heart Failure Association of the European Society of Cardiology. Eur. J. Heart Fail. 2019, 21, 553–576. [Google Scholar] [CrossRef] [Green Version]
- Barth, A.S.; Kuner, R.; Buness, A.; Ruschhaupt, M.; Merk, S.; Zwermann, L.; Kaab, S.; Kreuzer, E.; Steinbeck, G.; Mansmann, U.; et al. Identification of a common gene expression signature in dilated cardiomyopathy across independent microarray studies. J. Am. Coll. Cardiol. 2006, 48, 1610–1617. [Google Scholar] [CrossRef] [Green Version]
- Staudt, A.; Eichler, P.; Trimpert, C.; Felix, S.B.; Greinacher, A. Fc(γ) receptors IIa on cardiomyocytes and their potential functional relevance in dilated cardiomyopathy. J. Am. Coll. Cardiol. 2007, 49, 1684–1692. [Google Scholar] [CrossRef] [Green Version]
- Kadhi, A.; Mohammed, F.; Nemer, G. The genetic pathways underlying immunotherapy in dilated cardiomyopathy. Front. Cardiovasc. Med. 2021, 8, 613295. [Google Scholar] [CrossRef]
- Trimpert, C.; Herda, L.R.; Eckerle, L.G.; Pohle, S.; Muller, C.; Landsberger, M.; Felix, S.B.; Staudt, A. Immunoadsorption in dilated cardiomyopathy: Long-term reduction of cardiodepressant antibodies. Eur. J. Clin. Investig. 2010, 40, 685–691. [Google Scholar] [CrossRef]
- Perez-Iratxeta, C.; Andrade-Navarro, M.A.; Wren, J.D. Evolving research trends in bioinformatics. Brief. Bioinform. 2007, 8, 88–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winnacker, E.L. Interdisciplinary sciences in the 21st century. Curr. Opin. Biotechnol. 2003, 14, 328–331. [Google Scholar] [CrossRef]
- van’t Veer, L.J.; Dai, H.; van de Vijver, M.J.; He, Y.D.; Hart, A.A.M.; Mao, M.; Peterse, H.L.; van der Kooy, K.; Marton, M.J.; Witteveen, A.T.; et al. Gene expression profiling predicts clinical outcome of breast cancer. Nature 2002, 415, 530–536. [Google Scholar] [CrossRef] [Green Version]
- Newman, A.M.; Liu, C.L.; Green, M.R.; Gentles, A.J.; Feng, W.; Xu, Y.; Hoang, C.D.; Diehn, M.; Alizadeh, A.A. Robust enumeration of cell subsets from tissue expression profiles. Nat. Methods 2015, 12, 453–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witt, E.; Hammer, E.; Dorr, M.; Weitmann, K.; Beug, D.; Lehnert, K.; Nauck, M.; Volker, U.; Felix, S.B.; Ameling, S. Correlation of gene expression and clinical parameters identifies a set of genes reflecting LV systolic dysfunction and morphological alterations. Physiol. Genomics 2019, 51, 356–367. [Google Scholar] [CrossRef]
- Ameling, S.; Herda, L.R.; Hammer, E.; Steil, L.; Teumer, A.; Trimpert, C.; Dorr, M.; Kroemer, H.K.; Klingel, K.; Kandolf, R.; et al. Myocardial gene expression profiles and cardiodepressant autoantibodies predict response of patients with dilated cardiomyopathy to immunoadsorption therapy. Eur. Heart J. 2013, 34, 666–675. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. Omics 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.G.; Yan, G.R.; He, Q.Y. DOSE: An R/Bioconductor package for disease ontology semantic and enrichment analysis. Bioinformatics 2015, 31, 608–609. [Google Scholar] [CrossRef] [Green Version]
- Zhao, E.; Xie, H.; Zhang, Y. Predicting diagnostic gene biomarkers associated with immune infiltration in patients with acute myocardial infarction. Front. Cardiovasc. Med. 2020, 7, 586871. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Zhuang, L.; Shen, X.; Yang, L. Glucotoxicity activation of IL6 and IL11 and subsequent induction of fibrosis may be involved in the pathogenesis of islet dysfunction. Front. Mol. Biosci. 2021, 8, 708127. [Google Scholar] [CrossRef]
- Vella, D.; Marini, S.; Vitali, F.; Di Silvestre, D.; Mauri, G.; Bellazzi, R. MTGO: PPI network analysis via topological and functional module identification. Sci. Rep. 2018, 8, 5499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, C.H.; Chen, S.H.; Wu, H.H.; Ho, C.W.; Ko, M.T.; Lin, C.Y. cytoHubba: Identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8, S11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robin, X.; Turck, N.; Hainard, A.; Tiberti, N.; Lisacek, F.; Sanchez, J.C.; Muller, M. pROC: An open-source package for R and S+ to analyze and compare ROC curves. BMC Bioinform. 2011, 12, 77. [Google Scholar] [CrossRef] [PubMed]
- Dagenais, G.R.; Leong, D.P.; Rangarajan, S.; Lanas, F.; Lopez-Jaramillo, P.; Gupta, R.; Diaz, R.; Avezum, A.; Oliveira, G.; Wielgosz, A.; et al. Variations in common diseases, hospital admissions, and deaths in middle-aged adults in 21 countries from five continents (PURE): A prospective cohort study. Lancet 2020, 395, 785–794. [Google Scholar] [CrossRef]
- Smith, J.G. Molecular epidemiology of heart failure: Translational challenges and opportunities. JACC Basic Transl. Sci. 2017, 2, 757–769. [Google Scholar] [CrossRef]
- Zhang, Y.; Bauersachs, J.; Langer, H.F. Immune mechanisms in heart failure. Eur. J. Heart Fail. 2017, 19, 1379–1389. [Google Scholar] [CrossRef] [Green Version]
- Carrillo-Salinas, F.J.; Ngwenyama, N.; Anastasiou, M.; Kaur, K.; Alcaide, P. Heart inflammation: Immune cell roles and roads to the heart. Am. J. Pathol. 2019, 189, 1482–1494. [Google Scholar] [CrossRef] [Green Version]
- Seferovic, P.M.; Polovina, M.M.; Coats, A. Heart failure in dilated non-ischaemic cardiomyopathy. Eur. Heart J. Suppl. 2019, 21, M40–M43. [Google Scholar] [CrossRef] [Green Version]
- Pistulli, R.; Konig, S.; Drobnik, S.; Kretzschmar, D.; Rohm, I.; Lichtenauer, M.; Fritzenwanger, M.; Mall, G.; Mall, G.; Figulla, H.R.; et al. Decrease in dendritic cells in endomyocardial biopsies of human dilated cardiomyopathy. Eur. J. Heart Fail. 2013, 15, 974–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, S.H.; Xu, J.D.; Sun, S.N.; Li, Y.; Zhou, Z.; Li, H.; Liu, X.; Deng, J.P.; Huang, Y.S.; Chen, Z.X.; et al. Single-cell transcriptomic analyses of cardiac immune cells reveal that Rel-driven CD72-positive macrophages induce cardiomyocyte injury. Cardiovasc. Res. 2021, cvab193. [Google Scholar] [CrossRef] [PubMed]
- Staudt, A.; Staudt, Y.; Dorr, M.; Bohm, M.; Knebel, F.; Hummel, A.; Wunderle, L.; Tiburcy, M.; Wernecke, K.D.; Baumann, G.; et al. Potential role of humoral immunity in cardiac dysfunction of patients suffering from dilated cardiomyopathy. J. Am. Coll. Cardiol. 2004, 44, 829–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horn, S.; Lueking, A.; Murphy, D.; Staudt, A.; Gutjahr, C.; Schulte, K.; Konig, A.; Landsberger, M.; Lehrach, H.; Felix, S.B.; et al. Profiling humoral autoimmune repertoire of dilated cardiomyopathy (DCM) patients and development of a disease-associated protein chip. Proteomics 2006, 6, 605–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afanasyeva, M.; Georgakopoulos, D.; Rose, N.R. Autoimmune myocarditis: Cellular mediators of cardiac dysfunction. Autoimmun. Rev. 2004, 3, 476–486. [Google Scholar] [CrossRef] [PubMed]
- Kanda, T.; Yokoyama, T.; Suzuki, T.; Murata, K. Functional abnormalities of circulating natural killer cell subpopulations in patients with dilated cardiomyopathy. Tohoku J. Exp. Med. 1992, 168, 529–537. [Google Scholar] [CrossRef] [Green Version]
- Ohlow, M.A.; Brunelli, M.; Schreiber, M.; Lauer, B. Therapeutic effect of immunoadsorption and subsequent immunoglobulin substitution in patients with dilated cardiomyopathy: Results from the observational prospective Bad Berka Registry. J. Cardiol. 2017, 69, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Bhardwaj, G.; Dorr, M.; Sappa, P.K.; Ameling, S.; Dhople, V.; Steil, L.; Klingel, K.; Empen, K.; Beug, D.; Volker, U.; et al. Endomyocardial proteomic signature corresponding to the response of patients with dilated cardiomyopathy to immunoadsorption therapy. J. Proteomics 2017, 150, 121–129. [Google Scholar] [CrossRef]
- Grover, R.; Burse, S.A.; Shankrit, S.; Aggarwal, A.; Kirty, K.; Narta, K.; Srivastav, R.; Ray, A.K.; Malik, G.; Vats, A.; et al. Myg1 exonuclease couples the nuclear and mitochondrial translational programs through RNA processing. Nucleic Acids Res. 2019, 47, 5852–5866. [Google Scholar] [CrossRef] [Green Version]
- Philips, M.A.; Vikesa, J.; Luuk, H.; Jonson, L.; Lillevali, K.; Rehfeld, J.F.; Vasar, E.; Koks, S.; Nielsen, F.C. Characterization of MYG1 gene and protein: Subcellular distribution and function. Biol. Cell 2009, 101, 361–373. [Google Scholar] [CrossRef]
- Han, X.; Li, A.; Wang, W.; Du, L.; Wang, C.; Huang, G. MYG1 promotes proliferation and inhibits autophagy in lung adenocarcinoma cells via the AMPK/mTOR complex 1 signaling pathway. Oncol. Lett. 2021, 21, 334. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, A.M.; Minguet, S. Caveolin-1, tetraspanin CD81 and flotillins in lymphocyte cell membrane organization, signaling and immunopathology. Biochem. Soc. Trans. 2020, 48, 2387–2397. [Google Scholar] [CrossRef] [PubMed]
- Slaughter, N.; Laux, I.; Tu, X.; Whitelegge, J.; Zhu, X.; Effros, R.; Bickel, P.; Nel, A. The flotillins are integral membrane proteins in lipid rafts that contain TCR-associated signaling components: Implications for T-cell activation. Clin. Immunol. 2003, 108, 138–151. [Google Scholar] [CrossRef]
- Neumann-Giesen, C.; Fernow, I.; Amaddii, M.; Tikkanen, R. Role of EGF-induced tyrosine phosphorylation of reggie-1/flotillin-2 in cell spreading and signaling to the actin cytoskeleton. J. Cell Sci. 2007, 120, 395–406. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.; Kang, Y.; Yan, C.; Yang, C.; Zhang, T.; Huo, H.; Liu, Y. LUBAC and OTULIN regulate autophagy initiation and maturation by mediating the linear ubiquitination and the stabilization of ATG13. Autophagy 2021, 17, 1684–1699. [Google Scholar] [CrossRef]
- Levine, B.; Mizushima, N.; Virgin, H.W. Autophagy in immunity and inflammation. Nature 2011, 469, 323–335. [Google Scholar] [CrossRef]
- Enomoto, H.; Mittal, N.; Inomata, T.; Arimura, T.; Izumi, T.; Kimura, A.; Fukuda, K.; Makino, S. Dilated cardiomyopathy-linked heat shock protein family D member 1 mutations cause up-regulation of reactive oxygen species and autophagy through mitochondrial dysfunction. Cardiovasc. Res. 2021, 117, 1118–1131. [Google Scholar] [CrossRef]
- Sweet, M.E.; Cocciolo, A.; Slavov, D.; Jones, K.L.; Sweet, J.R.; Graw, S.L.; Reece, T.B.; Ambardekar, A.V.; Bristow, M.R.; Mestroni, L.; et al. Transcriptome analysis of human heart failure reveals dysregulated cell adhesion in dilated cardiomyopathy and activated immune pathways in ischemic heart failure. BMC Genomics 2018, 19, 812. [Google Scholar] [CrossRef] [Green Version]
- Qiao, A.; Zhao, Z.; Zhang, H.; Sun, Z.; Cui, X. Gene expression profiling reveals genes and transcription factors associated with dilated and ischemic cardiomyopathies. Pathol. Res. Pract. 2017, 213, 548–557. [Google Scholar] [CrossRef]
- Chen, L.; Yang, F.; Chen, X.; Rao, M.; Zhang, N.N.; Chen, K.; Deng, H.; Song, J.P.; Hu, S.S. Comprehensive myocardial proteogenomics profiling reveals C/EBPalpha as the key factor in the lipid storage of ARVC. J. Proteome Res. 2017, 16, 2863–2876. [Google Scholar] [CrossRef]
- Gaertner, A.; Schwientek, P.; Ellinghaus, P.; Summer, H.; Golz, S.; Kassner, A.; Schulz, U.; Gummert, J.; Milting, H. Myocardial transcriptome analysis of human arrhythmogenic right ventricular cardiomyopathy. Physiol. Genom. 2012, 44, 99–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Guan, H.; Liu, W.; Li, H.; Ding, J.; Feng, Y.; Chen, Z. Identification of Immune Markers in Dilated Cardiomyopathies with Heart Failure by Integrated Weighted Gene Coexpression Network Analysis. Genes 2022, 13, 393. https://doi.org/10.3390/genes13030393
Wang X, Guan H, Liu W, Li H, Ding J, Feng Y, Chen Z. Identification of Immune Markers in Dilated Cardiomyopathies with Heart Failure by Integrated Weighted Gene Coexpression Network Analysis. Genes. 2022; 13(3):393. https://doi.org/10.3390/genes13030393
Chicago/Turabian StyleWang, Xuehua, Hongquan Guan, Wei Liu, Huili Li, Jiaxing Ding, Yu Feng, and Zhijian Chen. 2022. "Identification of Immune Markers in Dilated Cardiomyopathies with Heart Failure by Integrated Weighted Gene Coexpression Network Analysis" Genes 13, no. 3: 393. https://doi.org/10.3390/genes13030393
APA StyleWang, X., Guan, H., Liu, W., Li, H., Ding, J., Feng, Y., & Chen, Z. (2022). Identification of Immune Markers in Dilated Cardiomyopathies with Heart Failure by Integrated Weighted Gene Coexpression Network Analysis. Genes, 13(3), 393. https://doi.org/10.3390/genes13030393