
Schizophrenia and Bipolar Polygenic Risk Scores in Relation to Intracranial Volume

 ,
, 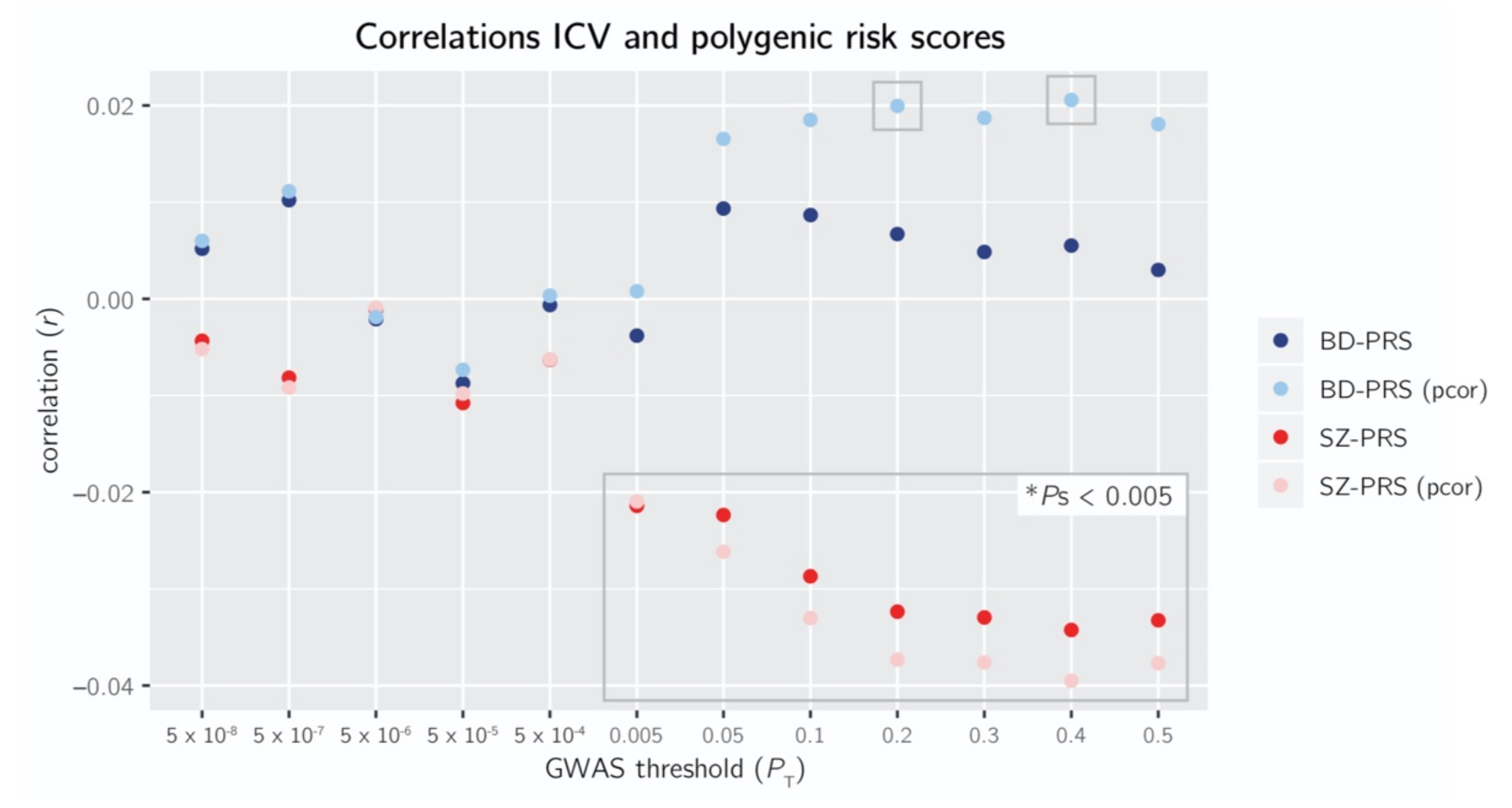{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sample
2.2. Image Acquisition and Processing
2.3. Polygenic Scoring
2.4. Statistical Analyses
3. Results
3.1. Relationship Schizophrenia Polygenic Risk Score and Intracranial Volume
3.2. Relationship Bipolar Disorder Polygenic Risk Score and Intracranial Volume
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pearlson, G.D. Etiologic, phenomenologic, and endophenotypic overlap of schizophrenia and bipolar disorder. Annu. Rev. Clin. Psychol. 2015, 11, 251–281. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Ripke, S.; Neale, B.M.; Faraone, S.V.; Purcell, S.M.; Perlis, R.H.; Mowry, B.J.; Thapar, A.; Goddard, M.E.; Witte, J.S.; et al. Genetic relationship between five psychiatric disorders estimated from genome-wide SNPs. Nat. Genet. 2013, 45, 984–994. [Google Scholar] [PubMed] [Green Version]
- Lichtenstein, P.; Yip, B.H.; Björk, C.; Pawitan, Y.; Cannon, T.D.; Sullivan, P.F.; Hultman, C.M. Common genetic determinants of schizophrenia and bipolar disorder in Swedish families: A population-based study. Lancet 2009, 373, 234–239. [Google Scholar] [CrossRef] [Green Version]
- Anttila, V.; Bulik-Sullivan, B.; Finucane, H.K.; Walters, R.K.; Bras, J.; Duncan, L.; Escott-Price, V.; Falcone, G.J.; Gormley, P.; Malik, R.; et al. Analysis of shared heritability in common disorders of the brain. Science 2018, 360, eaap8757. [Google Scholar] [PubMed] [Green Version]
- Van Erp, T.G.M.; Hibar, D.P.; Rasmussen, J.M.; Glahn, D.C.; Pearlson, G.D.; Andreassen, O.A.; Agartz, I.; Westlye, L.T.; Haukvik, U.K.; Dale, A.M.; et al. Subcortical brain volume abnormalities in 2028 individuals with schizophrenia and 2540 healthy controls via the ENIGMA consortium. Mol. Psychiatry 2016, 21, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Haijma, S.V.; Van Haren, N.; Cahn, W.; Koolschijn, P.C.M.P.; Hulshoff Pol, H.E.; Kahn, R.S. Brain volumes in schizophrenia: A meta-analysis in over 18,000 subjects. Schizophr. Bull. 2013, 39, 1129–1138. [Google Scholar] [CrossRef] [PubMed]
- Okada, N.; Fukunaga, M.; Yamashita, F.; Koshiyama, D.; Yamamori, H.; Ohi, K.; Yasuda, Y.; Fujimoto, M.; Watanabe, Y.; Yahata, N.; et al. Abnormal asymmetries in subcortical brain volume in schizophrenia. Mol. Psychiatry 2016, 21, 1460–1466. [Google Scholar] [CrossRef] [Green Version]
- Hibar, D.P.; Westlye, L.T.; Van Erp, T.G.M.; Rasmussen, J.; Leonardo, C.D.; Faskowitz, J.; Haukvik, U.K.; Hartberg, C.B.; Doan, N.T.; Agartz, I.; et al. Subcortical volumetric abnormalities in bipolar disorder. Mol. Psychiatry 2016, 21, 1710–1716. [Google Scholar] [CrossRef] [Green Version]
- Hulshoff Pol, H.E.; Van Baal, G.C.M.; Schnack, H.G.; Brans, R.G.H.; Van der Schot, A.C.; Brouwer, R.M.; Van Haren, N.E.M.; Lepage, C.; Collins, D.L.; Evans, A.C.; et al. Overlapping and segregating structural brain abnormalities in twins with schizophrenia or bipolar disorder. Arch. Gen. Psychiatry 2012, 69, 349–359. [Google Scholar]
- De Zwarte, S.M.C.; Brouwer, R.M.; Agartz, I.; Alda, M.; Aleman, A.; Alpert, K.I.; Bearden, C.E.; Bertolino, A.; Bois, C.; Bonvino, A.; et al. The association between familial risk and brain abnormalities is disease-specific: An ENIGMA–Relatives study of schizophrenia and bipolar disorder. Biol. Psychiatry 2019, 86, 545–556. [Google Scholar] [CrossRef] [Green Version]
- Peper, J.S.; Brouwer, R.M.; Boomsma, D.I.; Kahn, R.S.; Hulshoff Pol, H.E. Genetic influences on human brain structure: A review of brain imaging studies in twins. Hum. Brain Mapp. 2007, 28, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Cardno, A.G.; Gottesman, I.I. Twin studies of schizophrenia: From bow-and-arrow concordances to star wars Mx and functional genomics. Am. J. Med. Genet. 2000, 97, 12–17. [Google Scholar] [CrossRef]
- McGuffin, P.; Rijsdijk, F.; Andrew, M.; Sham, P.; Katz, R.; Cardno, A. The heritability of bipolar affective disorder and the genetic relationship to unipolar depression. Arch. Gen. Psychiatry 2003, 60, 497–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullins, N.; Forstner, A.J.; O’Connell, K.S.; Coombes, B.; Coleman, J.R.I.; Qiao, Z.; Als, T.D.; Bigdeli, T.B.; Børte, S.; Bryois, J.; et al. Genome-wide association study of more than 40,000 bipolar disorder cases provides new insights into the underlying biology. Nat. Genet. 2021, 53, 817–829. [Google Scholar] [CrossRef]
- Franke, B.; Stein, J.L.; Ripke, S.; Anttila, V.; Hibar, D.P.; Van Hulzen, K.J.E.; Arias-Vasquez, A.; Smoller, J.W.; Nichols, T.E.; Neale, M.C.; et al. Genetic influences on schizophrenia and subcortical brain volumes: Large-scale proof of concept. Nat. Neurosci. 2016, 19, 420–431. [Google Scholar] [CrossRef]
- Smeland, O.B.; Wang, Y.; Frei, O.; Li, W.; Hibar, D.P.; Franke, B.; Bettella, F.; Witoelar, A.; Djurovic, S.; Chen, C.-H.; et al. Genetic overlap between schizophrenia and volumes of hippocampus, putamen, and intracranial volume indicates shared molecular genetic mechanisms. Schizophr. Bull. 2018, 44, 854–864. [Google Scholar] [CrossRef]
- Lee, P.H.; Baker, J.T.; Holmes, A.J.; Jahanshad, N.; Ge, T.; Jung, J.Y.; Cruz, Y.; Manoach, D.S.; Hibar, D.P.; Faskowitz, J.; et al. Partitioning heritability analysis reveals a shared genetic basis of brain anatomy and schizophrenia. Mol. Psychiatry 2016, 21, 1680–1689. [Google Scholar] [CrossRef] [Green Version]
- Van der Merwe, C.; Passchier, R.; Mufford, M.; Ramesar, R.; Dalvie, S.; Stein, D.J. Polygenic risk for schizophrenia and associated brain structural changes: A systematic review. Compr. Psychiatry 2019, 88, 77–82. [Google Scholar] [CrossRef]
- Janiri, D.; Kotzalidis, G.D.; Di Luzio, M.; Giuseppin, G.; Simonetti, A.; Janiri, L.; Sani, G. Genetic neuroimaging of bipolar disorder: A systematic 2017-2020 update. Psychiatr. Genet. 2021, 31, 50–64. [Google Scholar] [CrossRef]
- Buckner, R.L.; Head, D.; Parker, J.; Fotenos, A.F.; Marcus, D.; Morris, J.C.; Snyder, A.Z. A unified approach for morphometric and functional data analysis in young, old, and demented adults using automated atlas-based head size normalization: Reliability and validation against manual measurement of total intracranial volume. Neuroimage 2004, 23, 724–738. [Google Scholar] [CrossRef]
- Ripke, S.; Neale, B.M.; Corvin, A.; Walters, J.T.R.; Farh, K.-H.; Holmans, P.A.; Lee, P.; Bulik-Sullivan, B.; Collier, D.A.; Huang, H.; et al. Biological insights from 108 schizophrenia-associated genetic loci. Nature 2014, 511, 421–427. [Google Scholar]
- Stahl, E.A.; Breen, G.; Forstner, A.J.; McQuillin, A.; Ripke, S.; Trubetskoy, V.; Mattheisen, M.; Wang, Y.; Coleman, J.R.I.; Gaspar, H.A.; et al. Genome-wide association study identifies 30 loci associated with bipolar disorder. Nat. Genet. 2019, 51, 793–803. [Google Scholar] [CrossRef] [PubMed]
- Price, A.L.; Weale, M.E.; Patterson, N.; Myers, S.R.; Need, A.C.; Shianna, K.V.; Ge, D.; Rotter, J.I.; Torres, E.; Taylor, K.D.; et al. Long-range LD can confound genome scans in admixed populations. Am. J. Hum. Genet. 2008, 83, 132–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Ji, L. Adjusting multiple testing in multilocus analyses using the eigenvalues of a correlation matrix. Heredity 2005, 95, 221–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyholt, D.R. A simple correction for multiple testing for single-nucleotide polymorphisms in linkage disequilibrium with each other. Am. J. Hum. Genet. 2004, 74, 765–769. [Google Scholar] [CrossRef] [Green Version]
- Sgouros, S.; Goldin, J.H.; Hockley, A.D.; Wake, M.J.; Natarajan, K. Intracranial volume change in childhood. J. Neurosurg. 1999, 91, 610–616. [Google Scholar] [CrossRef]
- Courchesne, E.; Chisum, H.J.; Townsend, J.; Cowles, A.; Covington, J.; Egaas, B.; Harwood, M.; Hinds, S.; Press, G.A. Normal brain development and aging: Quantitative analysis at in vivo MR imaging in healthy volunteers. Radiology 2000, 216, 672–682. [Google Scholar] [CrossRef]
- Kessler, R.C.; Amminger, G.P.; Aguilar-Gaxiola, S.; Alonso, J.; Lee, S.; Ustün, T.B. Age of onset of mental disorders: A review of recent literature. Curr. Opin. Psychiatry 2007, 20, 359–364. [Google Scholar] [CrossRef]
- Rajji, T.K.; Ismail, Z.; Mulsant, B.H. Age at onset and cognition in schizophrenia: Meta-analysis. Br. J. Psychiatry 2009, 195, 286–293. [Google Scholar] [CrossRef] [Green Version]
- Leboyer, M.; Henry, C.; Paillere-Martinot, M.-L.; Bellivier, F. Age at onset in bipolar affective disorder: A review. Bipolar Disord. 2005, 7, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.M.; Lewis, S.W. Is schizophrenia a neurodevelopmental disorder? Br. Med. J. (Clin. Res. Ed.). 1987, 295, 681–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasrallah, H.A. Neurodevelopmental aspects of bipolar affective disorder. Biol. Psychiatry 1991, 29, 1–2. [Google Scholar] [CrossRef]
- Weinberger, D.R. Implications of normal brain development for the pathogenesis of schizophrenia. Arch. Gen. Psychiatry 1987, 44, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.M.; Sham, P.; Van Os, J.; Zanelli, J.; Cannon, M.; McDonald, C. A developmental model for similarities and dissimilarities between schizophrenia and bipolar disorder. Schizophr. Res. 2004, 71, 405–416. [Google Scholar] [CrossRef]
- Parellada, M.; Gomez-Vallejo, S.; Burdeus, M.; Arango, C. Developmental differences between schizophrenia and bipolar disorder. Schizophr. Bull. 2017, 43, 1176–1189. [Google Scholar] [CrossRef]
- Walker, J.; Curtis, V.; Shaw, P.; Murray, R.M. Schizophrenia and bipolar disorder are distinguished mainly by differences in neurodevelopment. Neurotox. Res. 2002, 4, 427–436. [Google Scholar] [CrossRef]
- Gurholt, T.P.; Lonning, V.; Nerland, S.; Jørgensen, K.N.; Haukvik, U.K.; Alloza, C.; Arango, C.; Barth, C.; Bearden, C.E.; Berk, M.; et al. Intracranial and subcortical volumes in adolescents with early-onset psychosis: A multisite mega-analysis from the ENIGMA consortium. Hum. Brain Mapp. 2022, 43, 373–384. [Google Scholar] [CrossRef]
- Jalbrzikowski, M.; Hayes, R.A.; Wood, S.J.; Nordholm, D.; Zhou, J.H.; Fusar-Poli, P.; Uhlhaas, P.J.; Takahashi, T.; Sugranyes, G.; Kwak, Y.B.; et al. Association of structural magnetic resonance imaging measures with psychosis onset in individuals at clinical high risk for developing psychosis. JAMA Psychiatry 2021, 78, 753–766. [Google Scholar]
- Cross-Disorder Group of the Psychiatric Genomics Consortium. Identification of risk loci with shared effects on five major psychiatric disorders: A genome-wide analysis. Lancet 2013, 381, 1371–1379. [Google Scholar] [CrossRef]
- Ruderfer, D.M.; Ripke, S.; McQuillin, A.; Boocock, J.; Stahl, E.A.; Whitehead Pavlides, J.M.; Mullins, N.; Charney, A.W.; Ori, A.P.S.; Olde Loohuis, L.M.; et al. Genomic dissection of bipolar disorder and schizophrenia, including 28 subphenotypes. Cell 2018, 173, 1705–1715. [Google Scholar] [CrossRef] [Green Version]
- Van Rooij, D.; Anagnostou, E.; Arango, C.; Auzias, G.; Behrmann, M.; Busatto, G.F.; Calderoni, S.; Daly, E.; Deruelle, C.; Di Martino, A.; et al. Cortical and subcortical brain morphometry differences between patients with autism spectrum disorder and healthy individuals across the lifespan: Results from the ENIGMA ASD Working Group. Am. J. Psychiatry 2018, 175, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Hoogman, M.; Bralten, J.; Hibar, D.P.; Mennes, M.; Zwiers, M.P.; Schweren, L.S.J.; Van Hulzen, K.J.E.; Medland, S.E.; Shumskaya, E.; Jahanshad, N.; et al. Subcortical brain volume differences in participants with attention deficit hyperactivity disorder in children and adults: A cross-sectional mega-analysis. Lancet Psychiatry 2017, 4, 310–319. [Google Scholar] [CrossRef] [Green Version]
- Ching, C.R.K.; Gutman, B.A.; Sun, D.; Villalon Reina, J.; Ragothaman, A.; Isaev, D.; Zavaliangos-Petropulu, A.; Lin, A.; Jonas, R.K.; Kushan, L.; et al. Mapping subcortical brain alterations in 22q11.2 deletion syndrome: Effects of deletion size and convergence with idiopathic neuropsychiatric illness. Am. J. Psychiatry 2020, 177, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Caspi, Y.; Brouwer, R.M.; Schnack, H.G.; Van de Nieuwenhuijzen, M.E.; Cahn, W.; Kahn, R.S.; Niessen, W.J.; Van der Lugt, A.; Pol, H.H. Changes in the intracranial volume from early adulthood to the sixth decade of life: A longitudinal study. Neuroimage 2020, 220, 116842. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Zwarte, S.M.C.; Brouwer, R.M.; Kahn, R.S.; van Haren, N.E.M. Schizophrenia and Bipolar Polygenic Risk Scores in Relation to Intracranial Volume. Genes 2022, 13, 695. https://doi.org/10.3390/genes13040695
de Zwarte SMC, Brouwer RM, Kahn RS, van Haren NEM. Schizophrenia and Bipolar Polygenic Risk Scores in Relation to Intracranial Volume. Genes. 2022; 13(4):695. https://doi.org/10.3390/genes13040695
Chicago/Turabian Stylede Zwarte, Sonja M. C., Rachel M. Brouwer, René S. Kahn, and Neeltje E. M. van Haren. 2022. "Schizophrenia and Bipolar Polygenic Risk Scores in Relation to Intracranial Volume" Genes 13, no. 4: 695. https://doi.org/10.3390/genes13040695
APA Stylede Zwarte, S. M. C., Brouwer, R. M., Kahn, R. S., & van Haren, N. E. M. (2022). Schizophrenia and Bipolar Polygenic Risk Scores in Relation to Intracranial Volume. Genes, 13(4), 695. https://doi.org/10.3390/genes13040695