
Short-Term Responses of Soil Microbial Communities to Changes in Air Temperature, Soil Moisture and UV Radiation




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Standard Reference Soil
2.2. Soil Invertebrates
2.3. Experimental Soil Incubations
2.4. Community Level Physiological Profiles
2.5. DNA Extraction and Quantitative PCR
2.6. Illumina High-Throughput Sequencing
2.7. Inference of Soil Microbiome Functions from 16S rRNA Gene Sequencing Data
2.8. Statistical Analysis
3. Results
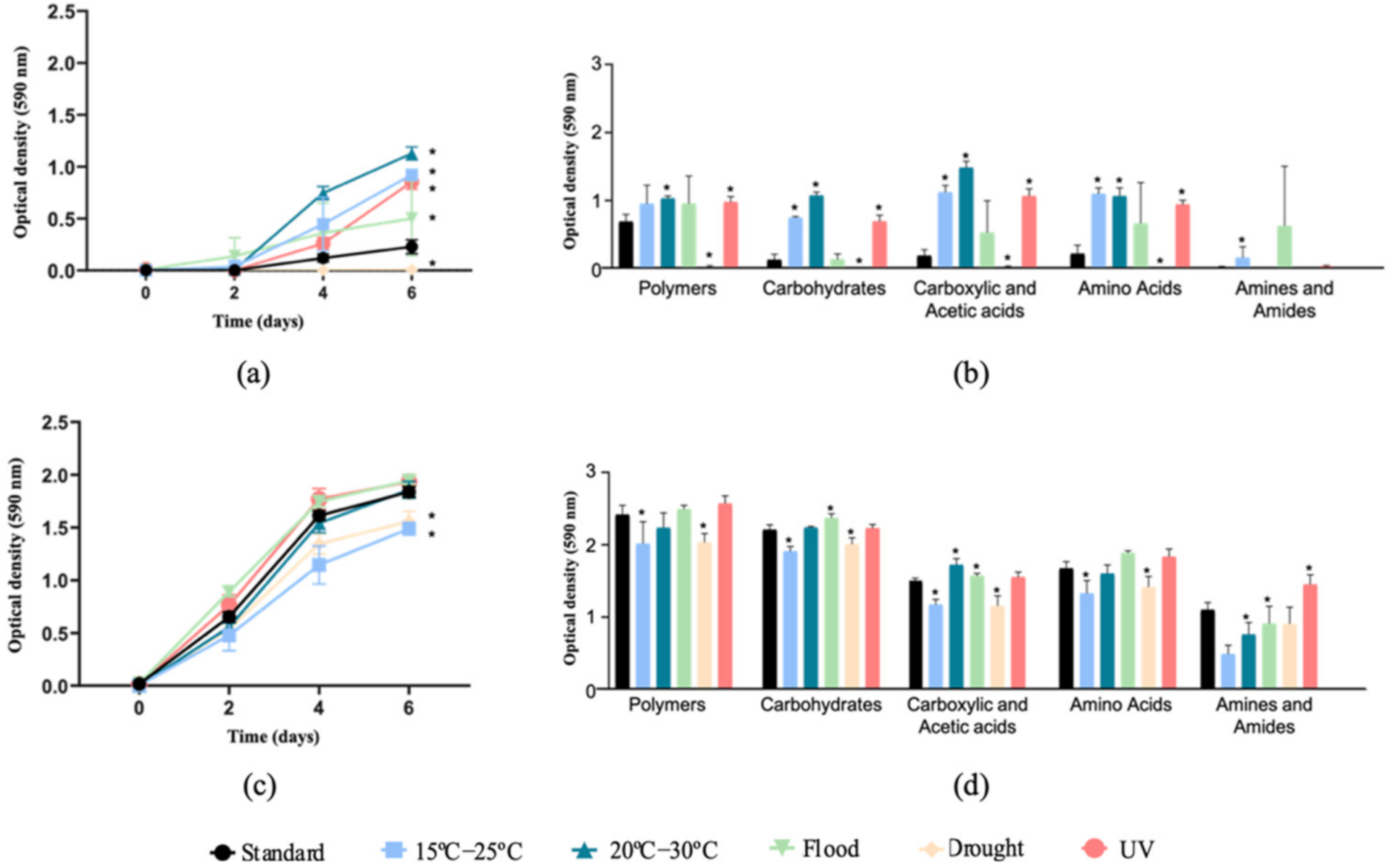
3.1. Community Level Physiological Profiles
3.2. Bacterial Community Structure and Composition
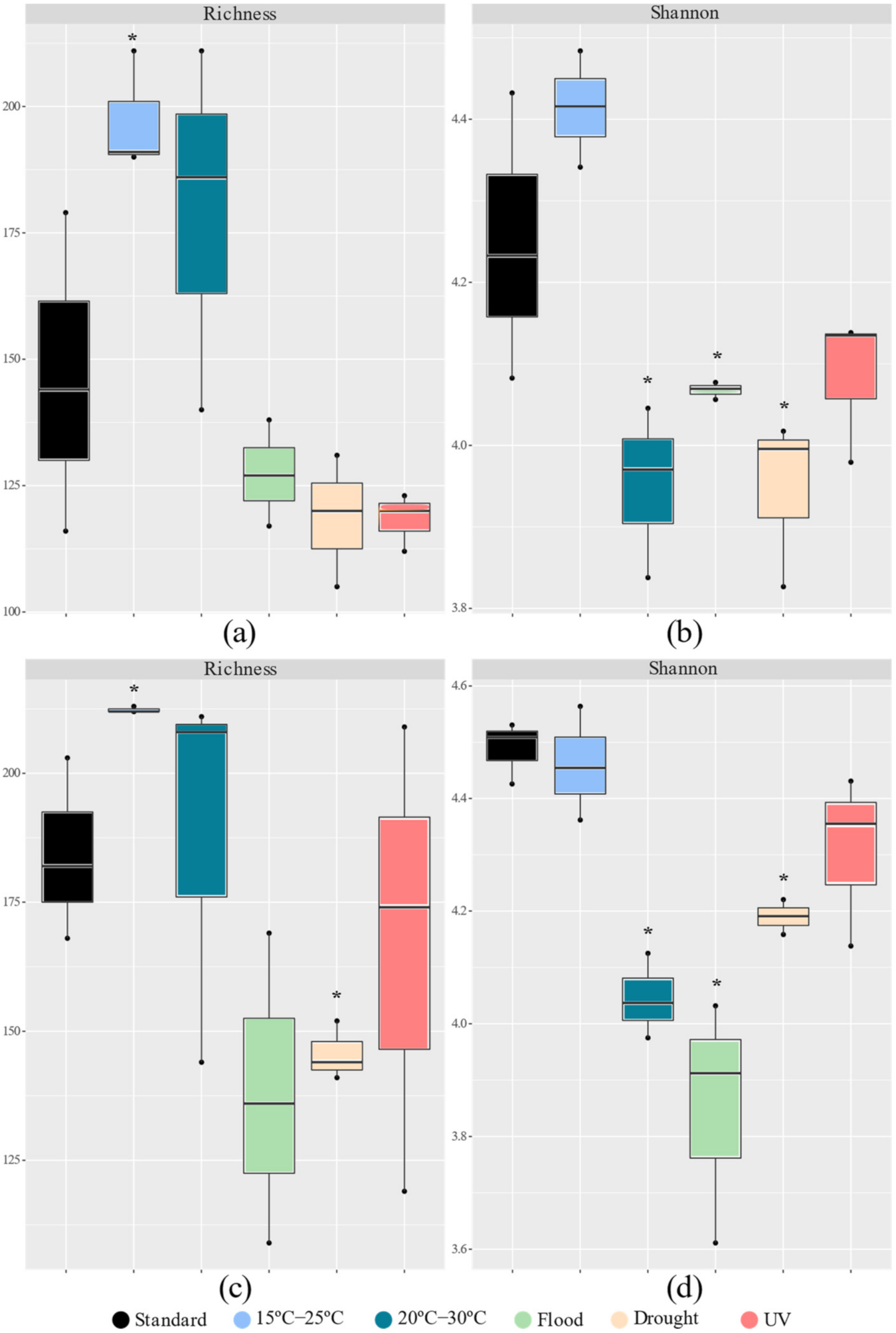
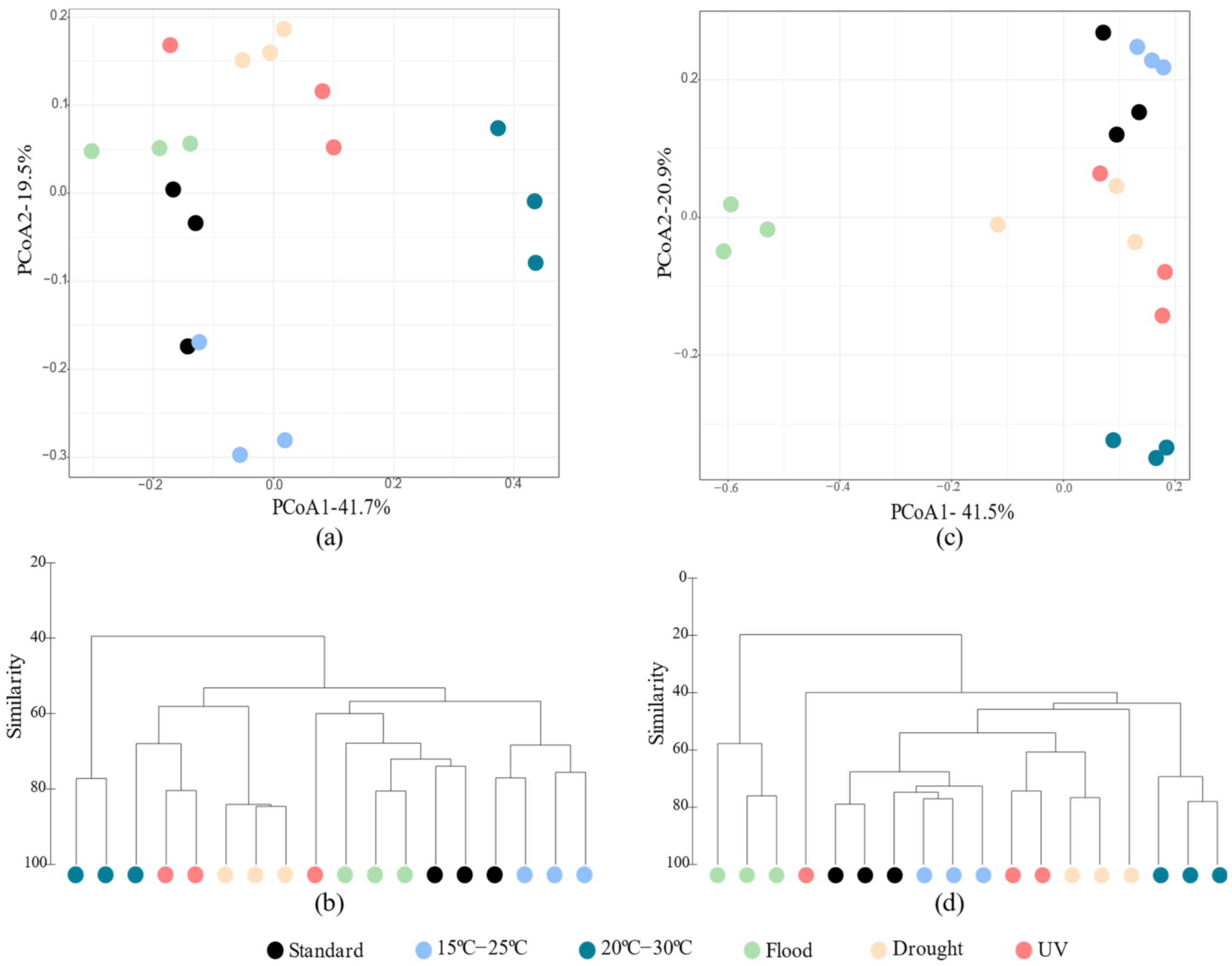
3.2.1. Bacterial Abundance and α and β-diversity
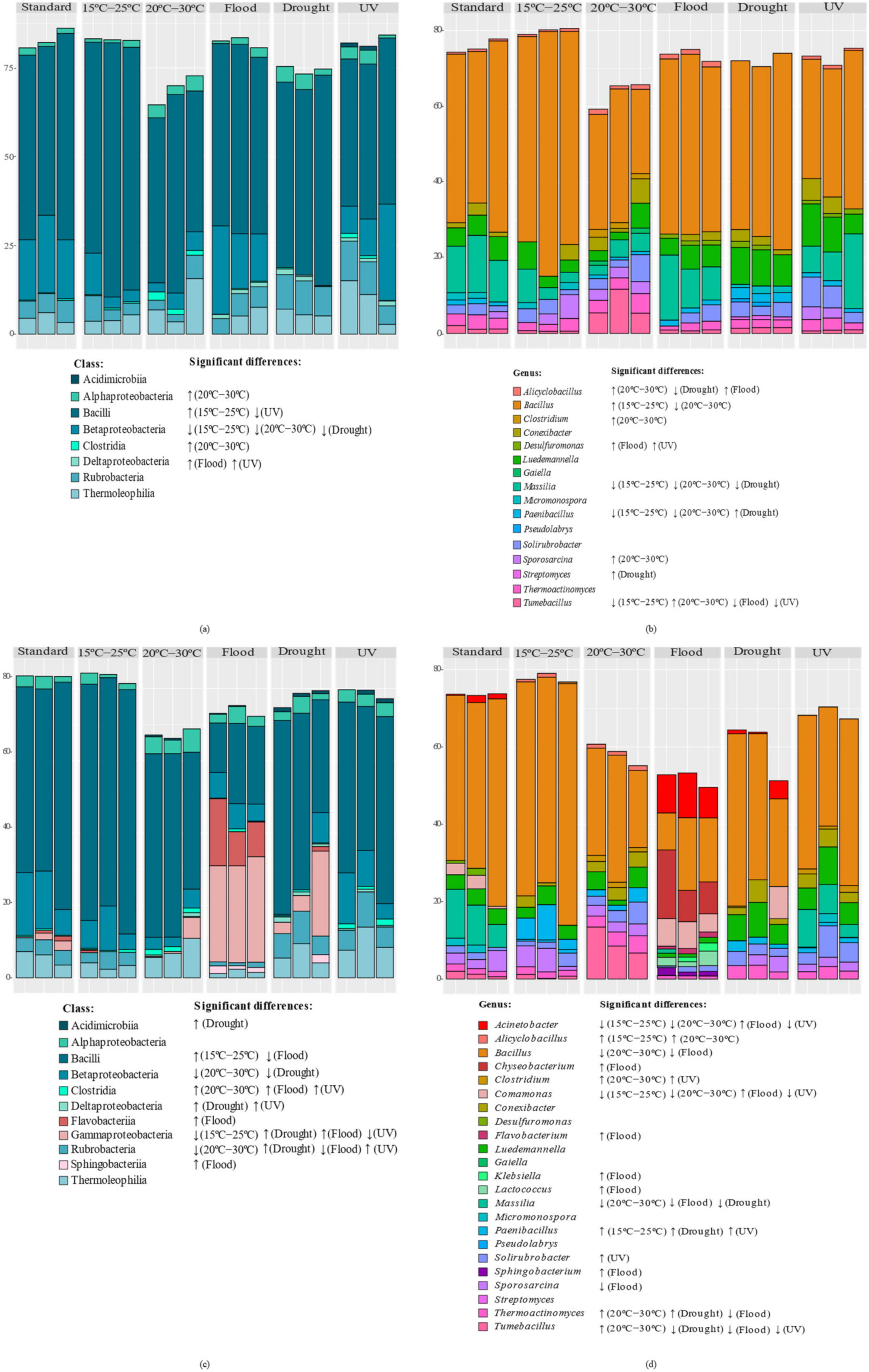
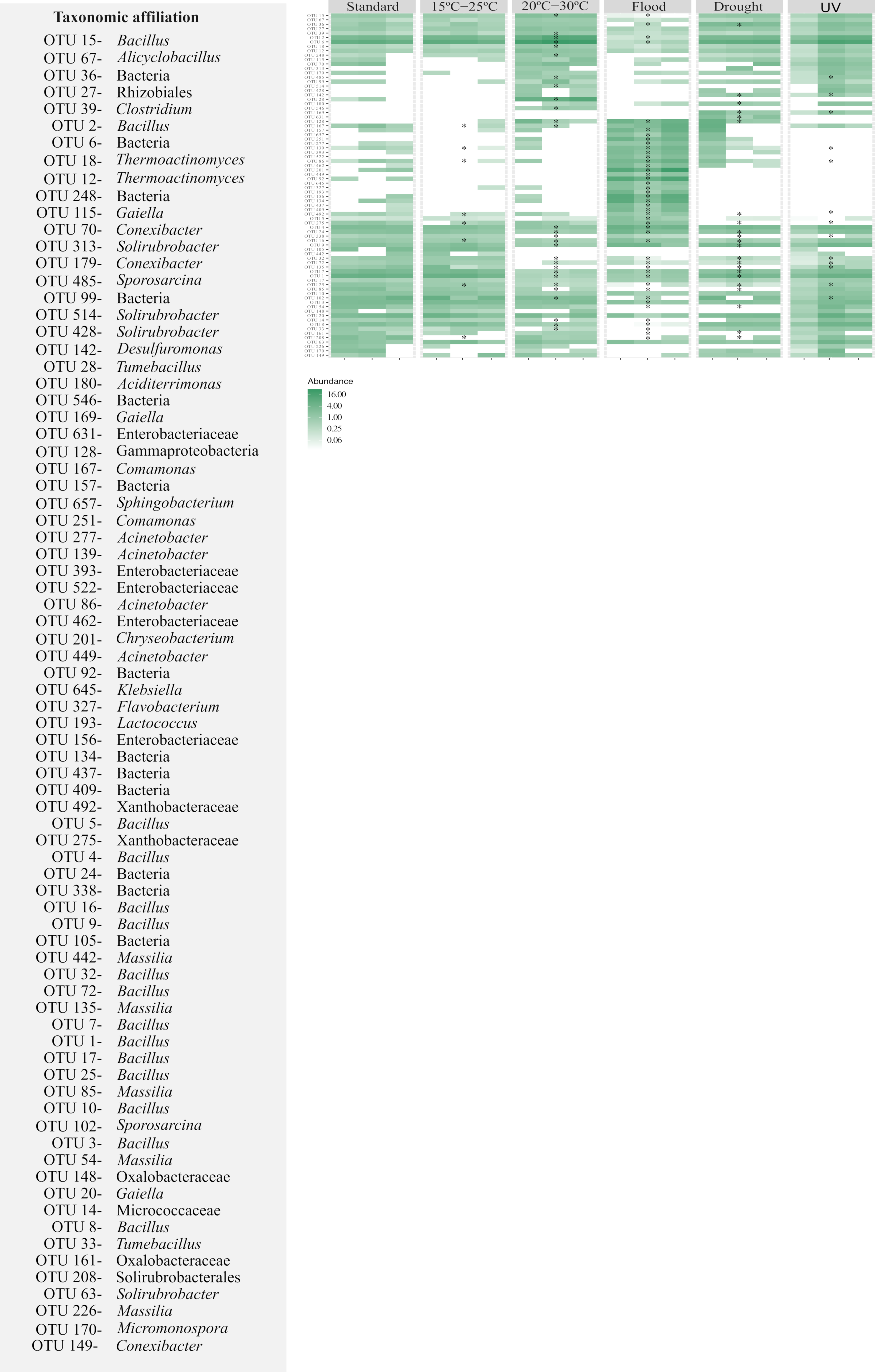
3.2.2. Taxonomic Composition of Bacterial Communities
3.3. Predicted Functional Pathways
4. Discussion
4.1. Effects of Rising Air Temperature
4.2. Effects of Flood and Drought Conditions
4.3. Effects of UV Radiation Exposure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Miyazaki, T.; Kato, C. World Soil Resources at Stake. In Human Geoscience; Himiyama, Y., Satake, K., Oki, T., Eds.; Springer: Singapore, 2020; pp. 75–85. [Google Scholar] [CrossRef]
- Wilhelm, R.C.; van Es, H.M.; Buckley, D.H. Predicting measures of soil health using the microbiome and supervised machine learning. Soil Biol. Biochem. 2022, 164, 108472. [Google Scholar] [CrossRef]
- Jansson, J.K.; Hofmockel, K.S. The Soil Microbiome—From Metagenomics to Metaphenomics. Curr. Opin. Microbiol. 2018, 43, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Dubey, A.; Malla, M.A.; Khan, F.; Chowdhary, K.; Yadav, S.; Kumar, A.; Sharma, S.; Khare, P.K.; Khan, M.L. Soil microbiome: A key player for conservation of soil health under changing climate. Biodivers. Conserv. 2019, 28, 2405–2429. [Google Scholar] [CrossRef]
- Jansson, J.K.; Hofmockel, K.S. Soil Microbiomes and Climate Change. Nat. Rev. Microbiol. 2020, 18, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.K.; Bardgett, R.D.; Smith, P.; Reay, D.S. Microorganisms and Climate Change: Terrestrial Feedbacks and Mitigation Options. Nat. Rev. Microbiol. 2010, 8, 779–790. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.J.; Edmonds, J.; Hartin, C.A.; Mundra, A.; Calvin, K. Near-Term Acceleration in the Rate of Temperature Change. Nat. Clim. Chang. 2015, 5, 333–336. [Google Scholar] [CrossRef]
- McHugh, T.A.; Compson, Z.; Van Gestel, N.; Hayer, M.; Ballard, L.; Haverty, M.; Hines, J.; Irvine, N.; Krassner, D.; Lyons, T.; et al. Climate Controls Prokaryotic Community Composition in Desert Soils of the Southwestern United States. FEMS Microbiol. Ecol. 2017, 93, fix116. [Google Scholar] [CrossRef]
- Chowdhury, T.R.; Lee, J.-Y.; Bottos, E.M.; Brislawn, C.J.; Iii, R.A.W.; Bramer, L.M.; Brown, J.; Zucker, J.D.; Kim, Y.-M.; Jumpponen, A.; et al. Metaphenomic Responses of a Native Prairie Soil Microbiome to Moisture Perturbations. mSystems 2019, 4, e00061-19. [Google Scholar] [CrossRef] [Green Version]
- Melillo, J.M.; Frey, S.D.; DeAngelis, K.M.; Werner, W.J.; Bernard, M.J.; Bowles, F.P.; Pold, G.; Knorr, M.A.; Grandy, A.S. Long-Term Pattern and Magnitude of Soil Carbon Feedback to the Climate System in a Warming World. Science 2017, 358, 101–105. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef] [Green Version]
- Hutchins, D.A.; Jansson, J.K.; Remais, J.V.; Rich, V.I.; Singh, B.K.; Trivedi, P. Climate change microbiology—problems and perspectives. Nat. Rev. Microbiol. 2019, 17, 391–396. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Climate Change 2013: The Physical Science Basis. In Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; 1535p. [Google Scholar]
- IPCC. Climate Change 2014: Impacts, Adaptation, and Vulnerability. In Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Field, C.B., Barros, V.R., Dokken, D.J., Mach, K.J., Mastrandrea, M.D., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; 1132p. [Google Scholar]
- IPCC. Global Warming of 1.5 °C. In An IPCC Special Report on the Impacts of Global Warming of 1.5 °C Above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Masson-Delmotte, V., Zhai, P., Pörtner, H.-O., Roberts, D., Skea, J., Shukla, P.R., Pirani, A., Moufouma-Okia, W., Péan, C., Pidcock, R., et al., Eds.; The Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2018. [Google Scholar]
- Didden, W.M. Ecology of terrestrial Enchytraeidae. Pedobiologia 1993, 37, 2–29. [Google Scholar]
- Didden, W.; Römbke, J. Enchytraeids as Indicator Organisms for Chemical Stress in Terrestrial Ecosystems. Ecotoxicol. Environ. Saf. 2001, 50, 25–43. [Google Scholar] [CrossRef] [PubMed]
- van Gestel, C. Soil ecotoxicology: State of the art and future directions. Zookeys 2012, 176, 275–296. [Google Scholar] [CrossRef] [Green Version]
- Lima, M.P.; Cardoso, D.N.; Soares, A.M.; Loureiro, S. Carbaryl Toxicity Prediction to Soil Organisms under High and Low Temperature Regimes. Ecotoxicol. Environ. Saf. 2015, 114, 263–272. [Google Scholar] [CrossRef]
- Blau, K.; Casadevall, L.; Wolters, B.; Van den Meersche, T.; Kreuzig, R.; Smalla, K.; Jechalke, S. Soil Texture-Depending Effects of Doxycycline and Streptomycin Applied with Manure on the Bacterial Community Composition and Resistome. FEMS Microbiol. Ecol. 2018, 94, fix145. [Google Scholar] [CrossRef]
- ISO 11274; Soil Quality—Determination of the Water-Retention Characteristic. ISO (International Organization for Standardization): Geneva, Switzerland, 1998.
- Jänsch, S.; Amorim, M.J.; Römbke, J. Identification of the Ecological Requirements of Important Terrestrial Ecotoxicological Test Species. Environ. Rev. 2005, 13, 51–83. [Google Scholar] [CrossRef]
- Kuperman, R.; Amorim, M.; Römbke, J.; Lanno, R.; Checkai, R.; Dodard, S.; Sunahara, G.; Scheffczyk, A. Adaptation of the Enchytraeid Toxicity Test for Use with Natural Soil Types. Eur. J. Soil Biol. 2006, 42 (Suppl. 1), S234–S243. [Google Scholar] [CrossRef]
- van Gestel, C.A.; Borgman, E.; Verweij, R.A.; Ortiz, M.D. The Influence of Soil Properties on the Toxicity of Molybdenum to Three Species of Soil Invertebrates. Ecotoxicol. Environ. Saf. 2011, 74, 1–9. [Google Scholar] [CrossRef]
- OECD. Guidelines for Testing Chemicals No. 217-Soil Microorganisms: Carbon Transformation Test; Organization for Economic Co-operation and Development: Paris, France, 2000. [Google Scholar]
- OECD. Guidelines for Testing Chemicals No. 216-Soil Microorganisms: Nitrogen Transformation Test; Organization for Economic Co-operation and Development: Paris, France, 2000. [Google Scholar]
- Samarajeewa, A.D.; Velicogna, J.R.; Princz, J.I.; Subasinghe, R.M.; Scroggins, R.P.; Beaudette, L.A. Effect of Silver Nano-Particles on Soil Microbial Growth, Activity and Community Diversity in a Sandy Loam Soil. Environ. Pollut. 2017, 220, 504–513. [Google Scholar] [CrossRef]
- Frąc, M.; Oszust, K.; Lipiec, J. Community Level Physiological Profiles (CLPP), Characterization and Microbial Activity of Soil Amended with Dairy Sewage Sludge. J. Sens. 2012, 12, 3253–3268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muyzer, G.; De Waal, E.C.; Uitterlinden, A.G. Profiling of Complex Microbial Populations by Denaturing Gradient Gel Electrophoresis Analysis of Polymerase Chain Reaction-Amplified Genes Coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brankatschk, R.; Bodenhausen, N.; Zeyer, J.; Bürgmann, H. Simple Absolute Quantification Method Correcting for Quantitative PCR Efficiency Variations for Microbial Community Samples. Appl. Environ. Microbiol. 2012, 78, 4481–4489. [Google Scholar] [CrossRef] [Green Version]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. J. Bioinform. 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Eren, A.M.; Morrison, H.G.; Lescault, P.J.; Reveillaud, J.; Vineis, J.H.; Sogin, M.L. Minimum entropy decomposition: Unsupervised oligotyping for sensitive partitioning of high-throughput marker gene sequences. ISME J. 2015, 9, 968–979. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Angly, F.E.; Dennis, P.G.; Skarshewski, A.; Vanwonterghem, I.; Hugenholtz, P.; Tyson, G.W. CopyRighter: A Rapid Tool for Improving the Accuracy of Microbial Community Profiles through Lineage-Specific Gene Copy Number Correction. Microbiome 2014, 2, 11. [Google Scholar] [CrossRef]
- Iwai, S.; Weinmaier, T.; Schmidt, B.L.; Albertson, D.G.; Poloso, N.J.; Dabbagh, K.; DeSantis, T.Z. Piphillin: Improved Prediction of Metagenomic Content by Direct Inference from Human Microbiomes. PLoS ONE 2016, 1, e0166104. [Google Scholar] [CrossRef] [Green Version]
- The Vegan Package. R Package Version 2.2-0. 2018. Available online: http://CRAN.R-project.org/ (accessed on 18 May 2021).
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing Austria, Vienna. 2020. Available online: https://www.R-project.org/ (accessed on 18 May 2021).
- Hayden, H.L.; Mele, P.M.; Bougoure, D.S.; Allan, C.Y.; Norng, S.; Piceno, Y.M.; Brodie, E.L.; DeSantis, T.Z.; Andersen, G.L.; Williams, A.L.; et al. Changes in the Microbial Community Structure of Bacteria, Archaea and Fungi in Response to Elevated CO2 and Warming in an Australian Native Grassland Soil. Environ. Microbiol. 2012, 14, 3081–3096. [Google Scholar] [CrossRef]
- Romero-Olivares, A.L.; Allison, S.D.; Treseder, K.K. Soil microbes and their response to experimental warming over time: A meta-analysis of field studies. Soil Biol. Biochem. 2017, 107, 32–40. [Google Scholar] [CrossRef] [Green Version]
- Walker, T.W.; Kaiser, C.; Strasser, F.; Herbold, C.W.; Leblans, N.I.; Woebken, D. Microbial temperature sensitivity and biomass change explain soil carbon loss with warming. Nat. Clim. Chang. 2018, 8, 885–889. [Google Scholar] [CrossRef] [PubMed]
- Shakoor, A.; Ashraf, F.; Shakoor, S.; Mustafa, A.; Rehman, A.; Altaf, M.M. Biogeochemical transformation of greenhouse gas emissions from terrestrial to atmospheric environment and potential feedback to climate forcing. Environ. Sci. Pollut. Res. 2020, 27, 38513–38536. [Google Scholar] [CrossRef] [PubMed]
- Mooshammer, M.; Hofhansl, F.; Frank, A.H.; Wanek, W.; Hämmerle, I.; Leitner, S.; Schnecker, J.; Wild, B.; Watzka, M.; Keiblinger, K.M.; et al. Decoupling of microbial carbon, nitrogen, and phosphorus cycling in response to extreme temperature events. Sci. Adv. 2017, 3, e1602781. [Google Scholar] [CrossRef] [Green Version]
- Sheik, C.S.; Beasley, W.H.; Elshahed, M.S.; Zhou, X.; Luo, Y.; Krumholz, L.R. Effect of Warming and Drought on Grassland Microbial Communities. ISME J. 2011, 5, 1692–1700. [Google Scholar] [CrossRef] [Green Version]
- Haugwitz, M.S.; Bergmark, L.; Priemé, A.; Christensen, S.; Beier, C.; Michelsen, A. Soil microorganisms respond to five years of climate change manipulations and elevated atmospheric CO2 in a temperate heath ecosystem. Plant Soil 2014, 374, 211–222. [Google Scholar] [CrossRef]
- Zhang, B.; Chen, S.; He, X.; Liu, W.; Zhao, Q.; Zhao, L.; Tian, C. Responses of soil microbial communities to experimental warming in alpine grasslands on the Qinghai-Tibet Plateau. PLoS ONE 2014, 9, e103859. [Google Scholar] [CrossRef] [Green Version]
- Hassan, N.; Anesio, A.M.; Rafiq, M.; Holtvoeth, J.; Bull, I.; Haleem, A.; Shah, A.A.; Hasan, F. Temperature Driven Membrane Lipid Adaptation in Glacial Psychrophilic Bacteria. Front. Microbiol. 2020, 11, 824. [Google Scholar] [CrossRef]
- Wu, J.; Xiong, J.; Hu, C.; Shi, Y.; Wang, K.; Zhang, D. Temperature Sensitivity of Soil Bacterial Community along Contrasting Warming Gradient. Appl. Soil Ecol. 2015, 94, 40–48. [Google Scholar] [CrossRef]
- Lin, Y.; Jia, Z.; Wang, D.; Chiu, Y. Effects of temperature on the composition and diversity of bacterial communities in bamboo soils at different elevations. Biogeosciences 2017, 14, 4879–4889. [Google Scholar] [CrossRef] [Green Version]
- Cavicchioli, R.; Ripple, W.J.; Timmis, K.N.; Azam, F.; Bakken, L.R.; Baylis, M. Scientists’ Warning to Humanity: Microorganisms and Climate Change. Nat. Rev. Microbiol. 2019, 17, 569–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeAngelis, K.M.; Pold, G.; Topã§uoäÿlu, B.D.; Van Diepen, L.T.A.; Varney, R.M.; Blanchard, J.; Emelillo, J.; Frey, S.D. Long-Term Forest Soil Warming Alters Microbial Communities in Temperate Forest Soils. Front. Microbiol. 2015, 6, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imam, S.; Noguera, D.R.; Donohue, T.J. CceR and AkgR Regulate Central Carbon and Energy Metabolism in Alphaproteobacteria. mBio 2015, 6, e02461-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yergeau, E.; Bokhorst, S.; Kang, S.; Zhou, J.; Greer, C.W.; Aerts, R.; Kioalchuk, G.A. Shifts in soil microorganisms in response to warming are consistent across a range of Antarctic environments. ISME J. 2012, 6, 692–702. [Google Scholar] [CrossRef] [Green Version]
- Xiong, J.; Sun, H.; Peng, F.; Zhang, H.; Xue, X.; Gibbons, S.; Gilbert, J.A.; Chu, H. Characterizing changes in soil bacterial community structure in response to short-term warming. FEMS Microbiol. Ecol. 2014, 89, 281–292. [Google Scholar] [CrossRef]
- Zhang, K.; Shi, Y.; Jing, X.; He, J.-S.; Sun, R.; Yang, Y.; Shade, A.; Chu, H. Effects of Short-Term Warming and Altered Precipitation on Soil Microbial Communities in Alpine Grassland of the Tibetan Plateau. Front. Microbiol. 2016, 7, 1032. [Google Scholar] [CrossRef] [Green Version]
- Radujković, D.; Verbruggen, E.; Sigurdsson, B.D.; Leblans, N.I.; Janssens, I.A.; Vicca, S.; Weedon, J.T. Prolonged Exposure Does Not Increase Soil Microbial Community Compositional Response to Warming along Geothermal Gradients. FEMS Microbiol. Ecol. 2017, 94. [Google Scholar] [CrossRef] [Green Version]
- De Vries, F.T.; Shade, A. Controls on Soil Microbial Community Stability under Climate Change. Front. Microbiol. 2013, 4, 265. [Google Scholar] [CrossRef] [Green Version]
- Obermeier, M.M.; Gnädinger, F.; Raj, A.C.D.; Obermeier, W.A.; Schmid, C.A.; Balàzs, H.; Schröder, P. Under Temperate Climate, the Conversion of Grassland to Arable Land Affects Soil Nutrient Stocks and Bacteria in a Short Term. Sci. Total Environ. 2020, 703, 135494. [Google Scholar] [CrossRef]
- Foesel, B.U.; Geppert, A.; Rohde, M.; Overmann, J. Parviterribacter Kavangonensis Gen. Nov., Sp. Nov. and Parviterribacter Multiflagellatus Sp. Nov., Novel Members of Parviterribacteraceae Fam. Nov. within the Order Solirubrobacterales, and Emended Descriptions of the Classes Thermoleophilia and Rubrobacteria and Their Orders and Families. Int. J. Syst. Evol. Microbiol. 2016, 66, 652–665. [Google Scholar] [CrossRef]
- Jurburg, S.D.; Nunes, I.; Brejnrod, A.; Jacquiod, S.; Priemé, A.; Sørensen, S.J.; Van Elsas, J.D.; Salles, J.F. Legacy Effects on the Recovery of Soil Bacterial Communities from Extreme Temperature Perturbation. Front. Microbiol. 2017, 8, 1832. [Google Scholar] [CrossRef] [PubMed]
- Yanni, S.F.; Helgason, B.L.; Janzen, H.H.; Ellert, B.H.; Gregorich, E.G. Warming Effects on Carbon Dynamics and Microbial Communities in Soils of Diverse Texture. Soil Biol. Biochem. 2020, 140, 107631. [Google Scholar] [CrossRef]
- Saxena, A.K.; Kumar, M.; Chakdar, H.; Anuroopa, N.; Bagyaraj, D.J. Bacillus Species in Soil as a Natural Resource for Plant Health and Nutrition. J. Appl. Microbiol. 2020, 128, 1583–1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavelle, P.; Decaëns, T.; Aubert, M.; Barot, S.; Blouin, M.; Bureau, F.; Margerie, P.; Mora, P.; Rossi, J.-P. Soil invertebrates and ecosystem services. Eur. J. Soil Biol. 2006, 42, S3–S15. [Google Scholar] [CrossRef]
- Peixoto, S.; Henriques, I.; Loureiro, S. Long-term effects of Cu(OH)2 nanopesticide exposure on soil microbial communities. Environ. Pollut. 2020, 269, 116113. [Google Scholar] [CrossRef]
- Hartmann, M.; Brunner, I.; Hagedorn, F.; Bardgett, R.D.; Stierli, B.; Herzog, C. A Decade of Irrigation Transforms the Soil Microbiome of a Semi-Arid Pine Forest. Mol. Ecol. 2017, 26, 1190–1206. [Google Scholar] [CrossRef]
- Zeng, Q.; Dong, Y.; An, S. Bacterial Community Responses to Soils along a Latitudinal and Vegetation Gradient on the Loess Plateau, China. PLoS ONE 2016, 11, e0152894. [Google Scholar] [CrossRef] [Green Version]
- Davidson, A.L.; Dassa, E.; Orelle, C.; Chen, J. Structure, Function, and Evolution of Bacterial ATP-Binding Cassette Systems. MMBR 2008, 72, 317–364. [Google Scholar] [CrossRef] [Green Version]
- Kotrba, P.; Inui, M.; Yukawa, H. Bacterial Phosphotransferase System (PTS) in Carbohydrate Uptake and Control of Carbon Metabolism. J. Biosci. Bioeng. 2001, 92, 502–517. [Google Scholar] [CrossRef]
- Whitman, W.B. (Ed.) Bergey’s Manual of Systematics of Archaea and Bacteria (410); Wiley: Hoboken, NJ, USA, 2015. [Google Scholar]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an Ecological Classification of Soil Bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Liu, D.; Chen, L.; Zhu, X.; Wang, Y.; Xuan, Y.; Liu, X.; Duan, Y. Klebsiella Pneumoniae SnebYK Mediates Resistance against Heterodera Glycines and Promotes Soybean Growth. Front. Microbiol. 2018, 9, 1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hueso, S.; García, C.; Hernández, T. Severe drought conditions modify the microbial community structure, size and activity in amended and unamended soils. Soil Biol. Biochem. 2012, 50, 167–173. [Google Scholar] [CrossRef]
- Schimel, J.; Balser, T.C.; Wallenstein, M. Microbial stress-response physiology and its implications for ecosystem function. Ecology 2007, 88, 1386–1394. [Google Scholar] [CrossRef] [PubMed]
- Fuchslueger, L.; Bahn, M.; Fritz, K.; Hasibeder, R.; Richter, A. Experimental drought reduces the transfer of recently fixed plant carbon to soil microbes and alters the bacterial community composition in a mountain meadow. New Phytol. 2014, 201, 916–927. [Google Scholar] [CrossRef] [PubMed]
- Bouskill, N.J.; Lim, H.C.; Borglin, S.; Salve, R.; Wood, T.E.; Silver, W.L.; Brodie, E.L. Pre-Exposure to Drought Increases the Resistance of Tropical Forest Soil Bacterial Communities to Extended Drought. ISME J. 2013, 7, 384–394. [Google Scholar] [CrossRef]
- Sunde, E.P.; Setlow, P.; Hederstedt, L.; Halle, B. The physical state of water in bacterial spores. Proc. Natl. Acad. Sci. USA 2009, 106, 19334–19339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, J.B.; Rock, C.O. Bacterial lipids: Metabolism and membrane homeostasis. Prog. Lipid Res. 2013, 52, 249–276. [Google Scholar] [CrossRef] [Green Version]
- Jimenez-Diaz, L.; Caballero, A.; Segura, A. Pathways for the Degradation of Fatty Acids in Bacteria. In Aerobic Utilization of Hydrocarbons, Oils and Lipids. Handbook of Hydrocarbon and Lipid Microbiology; Rojo, F., Ed.; Springer: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Taketani, R.G.; Lançoni, M.D.; Kavamura, V.N.; Durrer, A.; Andreote, F.D.; Melo, I.S. Dry Season Constrains Bacterial Phylogenetic Diversity in a Semi-Arid Rhizosphere System. Microb. Ecol. 2017, 73, 153–161. [Google Scholar] [CrossRef]
- Arrange, A.A.; Phelps, T.J.; Benoit, R.E.; Palumbo, A.V.; White, D.C. Bacterial Sensitivity to UV Light as a Model for Ionizing Radiation Resistance. J. Microbiol. Methods 1993, 18, 127–136. [Google Scholar] [CrossRef]
- Formánek, P.; Rejšek, K.; Vranová, V. Effect of Elevated CO2, O3, and UV Radiation on Soils. Sci. World J. 2014, 2014, 730149. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.; Campbell, C.D.; Lee, J.A.; Callaghan, T.V.; Gwynn-Jones, D. Arctic Microorganisms Respond More to Elevated UV-B Radiation than CO2. Nature 2002, 416, 82–83. [Google Scholar] [CrossRef] [PubMed]
- Niemi, R.; Martikainen, P.J.; Silvola, J.; Wulff, A.; Turtola, S.; Holopainen, T. Elevated UV-B Radiation Alters Fluxes of Methane and Carbon Dioxide in Peatland Microcosms. Glob. Chang. Biol. 2002, 8, 361–371. [Google Scholar] [CrossRef]
- Bao, T.; Zhu, R.; Wang, P.; Ye, W.; Ma, D.; Xu, H. Potential Effects of Ultraviolet Radiation Reduction on Tundra Nitrous Oxide and Methane Fluxes in Maritime Antarctica. Sci. Rep. 2018, 8, 3716. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, I.; Alves, M.; Malheiro, C.; Silva, A.R.R.; Loureiro, S.; Henriques, I.; González-Alcaraz, M.N. Short-Term Responses of Soil Microbial Communities to Changes in Air Temperature, Soil Moisture and UV Radiation. Genes 2022, 13, 850. https://doi.org/10.3390/genes13050850
Silva I, Alves M, Malheiro C, Silva ARR, Loureiro S, Henriques I, González-Alcaraz MN. Short-Term Responses of Soil Microbial Communities to Changes in Air Temperature, Soil Moisture and UV Radiation. Genes. 2022; 13(5):850. https://doi.org/10.3390/genes13050850
Chicago/Turabian StyleSilva, Isabel, Marta Alves, Catarina Malheiro, Ana Rita R. Silva, Susana Loureiro, Isabel Henriques, and M. Nazaret González-Alcaraz. 2022. "Short-Term Responses of Soil Microbial Communities to Changes in Air Temperature, Soil Moisture and UV Radiation" Genes 13, no. 5: 850. https://doi.org/10.3390/genes13050850
APA StyleSilva, I., Alves, M., Malheiro, C., Silva, A. R. R., Loureiro, S., Henriques, I., & González-Alcaraz, M. N. (2022). Short-Term Responses of Soil Microbial Communities to Changes in Air Temperature, Soil Moisture and UV Radiation. Genes, 13(5), 850. https://doi.org/10.3390/genes13050850