
Exosomal miRNA Profile in Small-for-Gestational-Age Children: A Potential Biomarker for Catch-Up Growth

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. Clinical Characteristics of the Study Subjects
2.3. Sample Collection
2.4. Exosome Isolation from Human Serum
2.5. Serum and Exosomal miRNA Next-Generation Sequencing (NGS)
2.6. Differential miRNA Expression Analysis
2.7. Statistical Analysis
3. Results
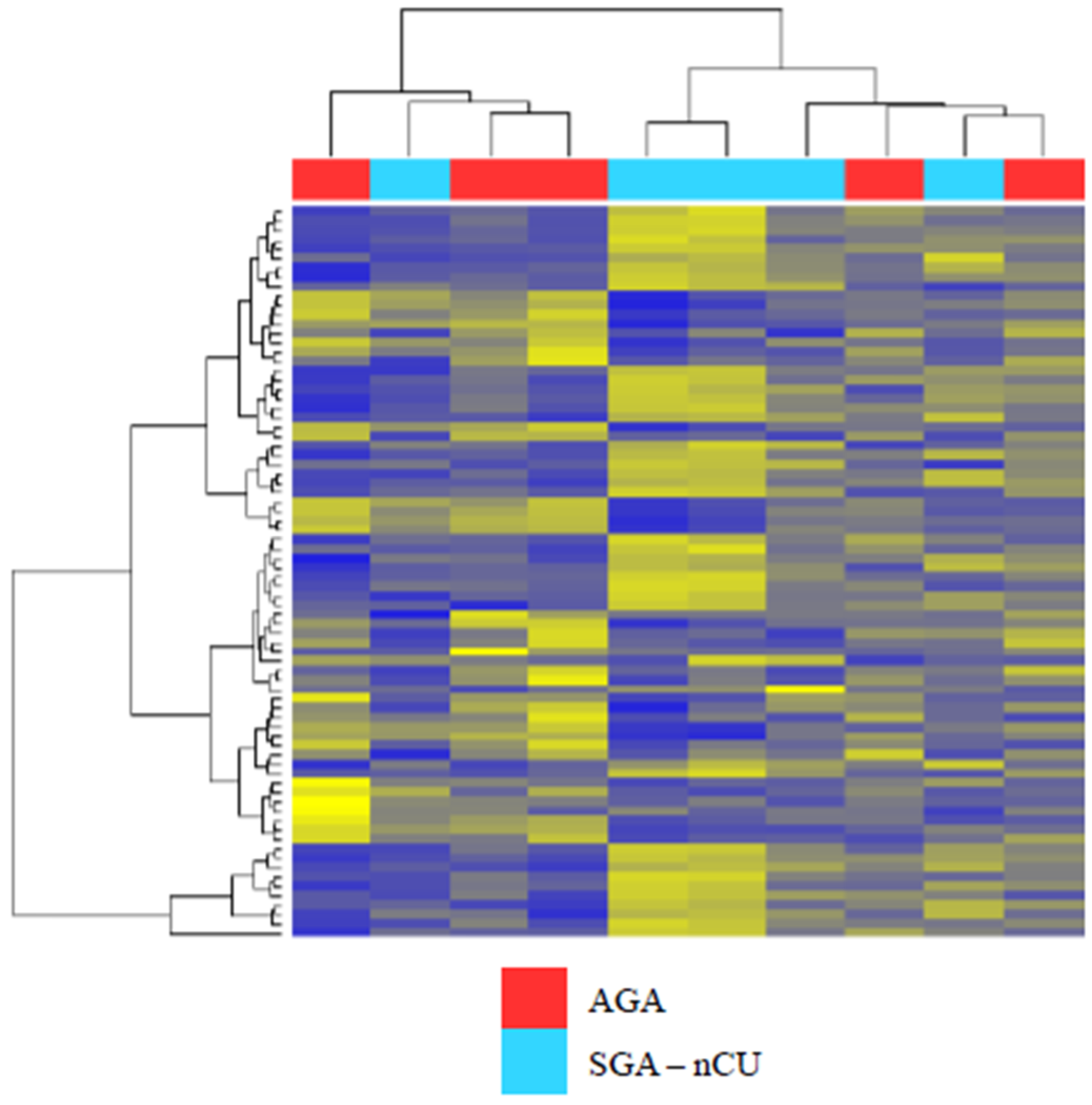
3.1. Profile of Circulating Serum miRNA Expression in SGA-nCU and AGA
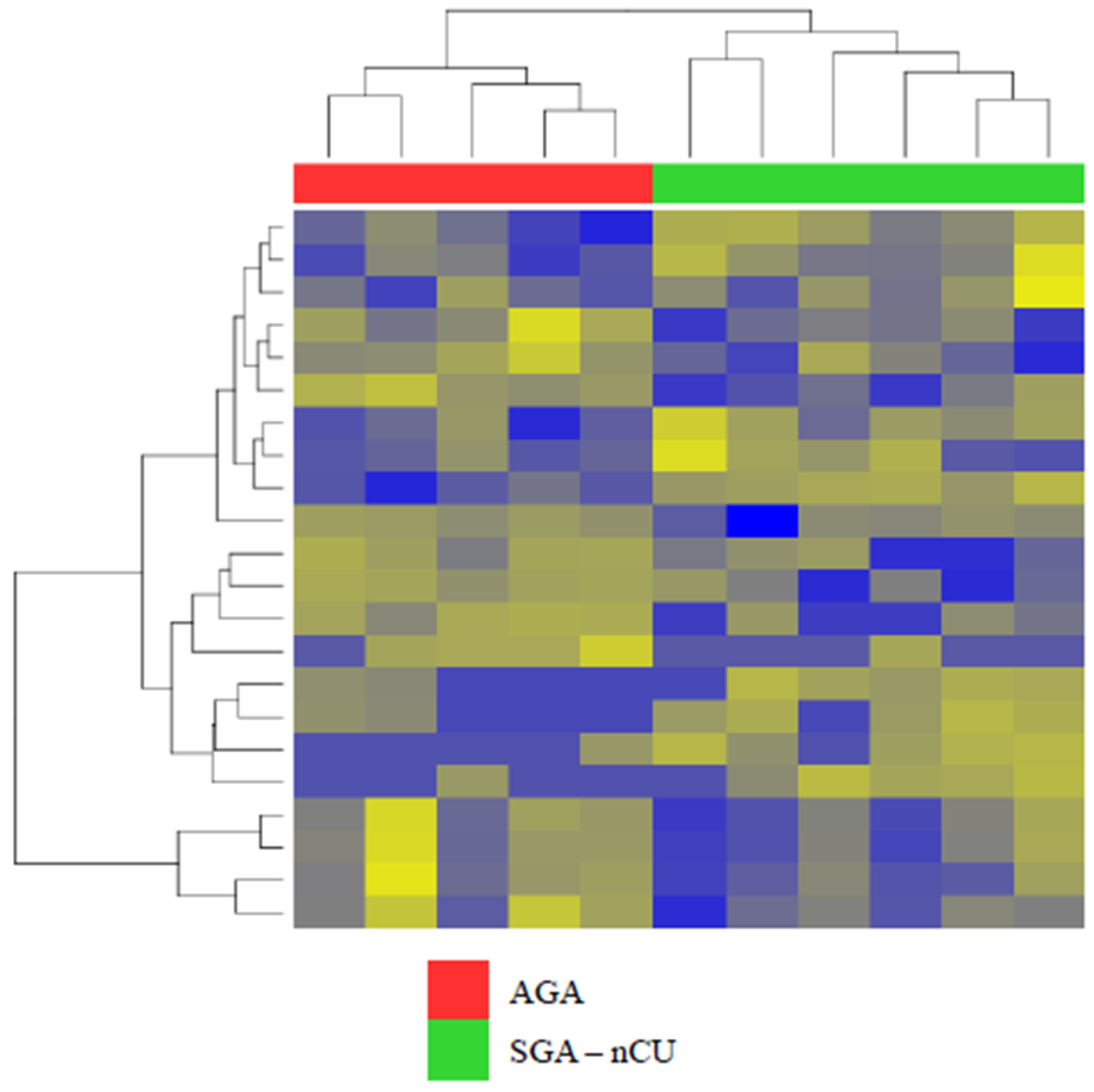
3.2. Exosomal miRNA Profiles Show Different Expression Patterns than Those Collected from Serum
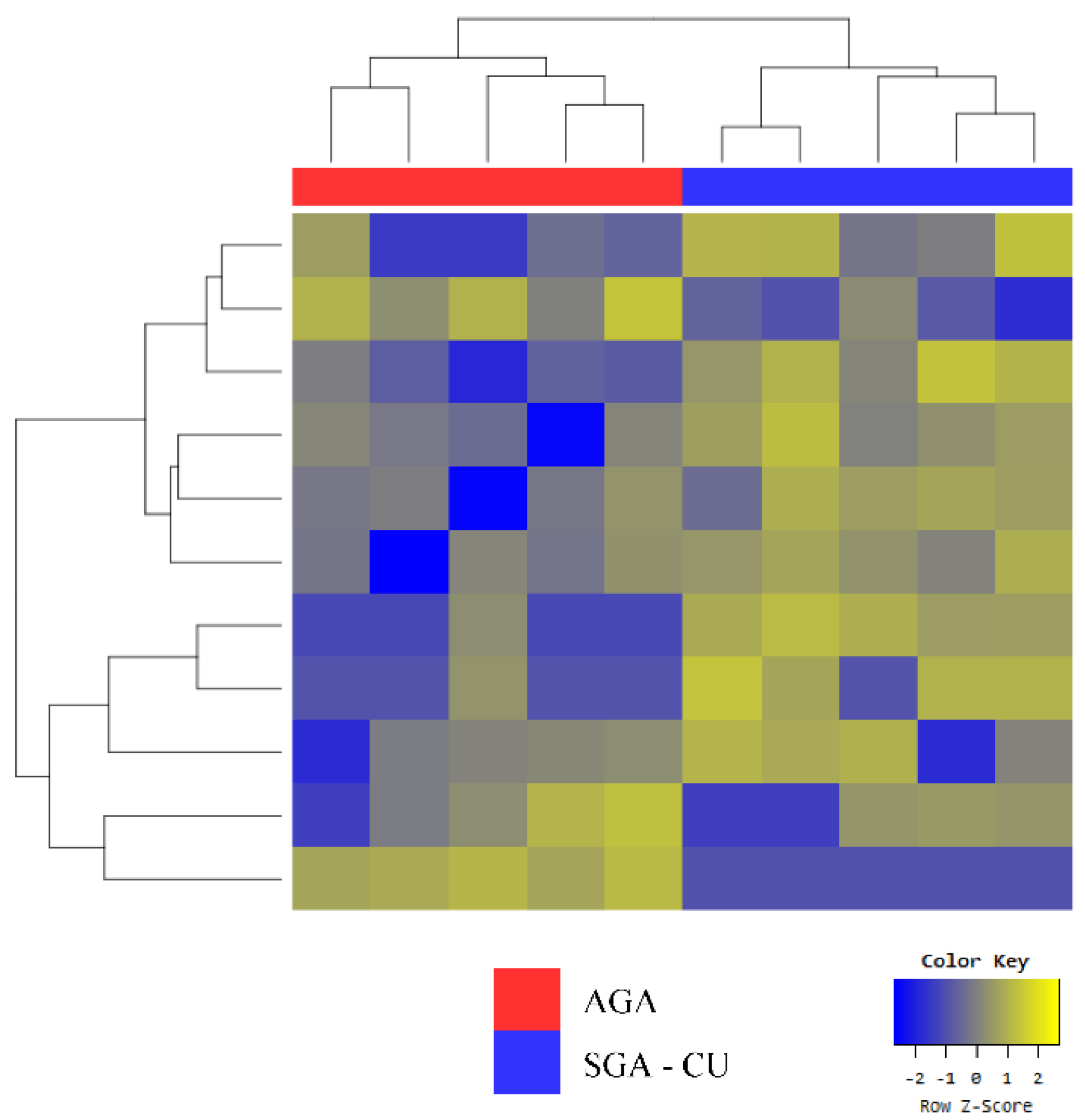
3.3. Exosomal miRNA Profiles of SGA-CU Were More Similar to Those of AGA than Those of SGA-nCU
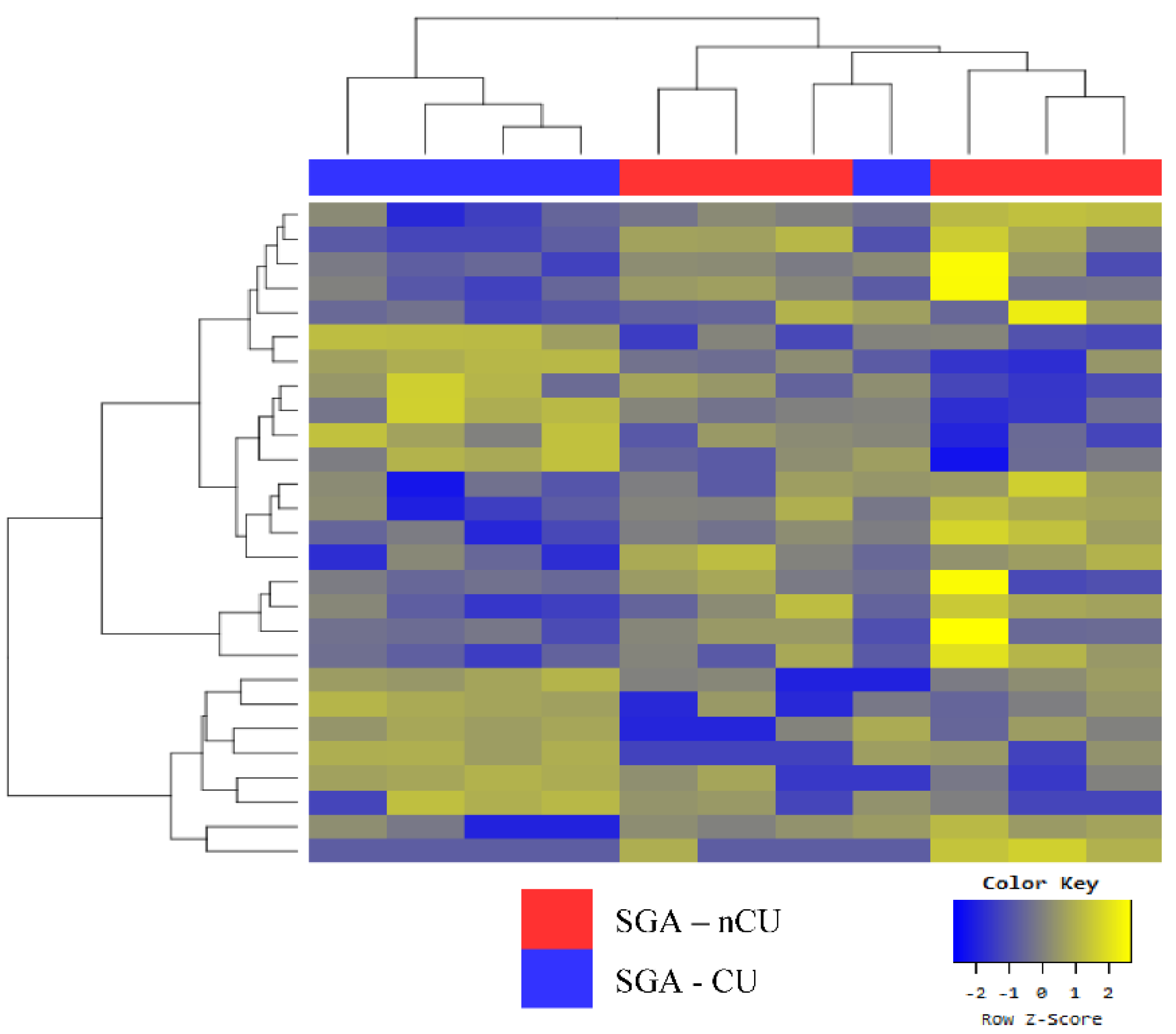
3.4. Exosomal miRNA Profiles of SGA-nCU and SGA-CU Show Potential Differences
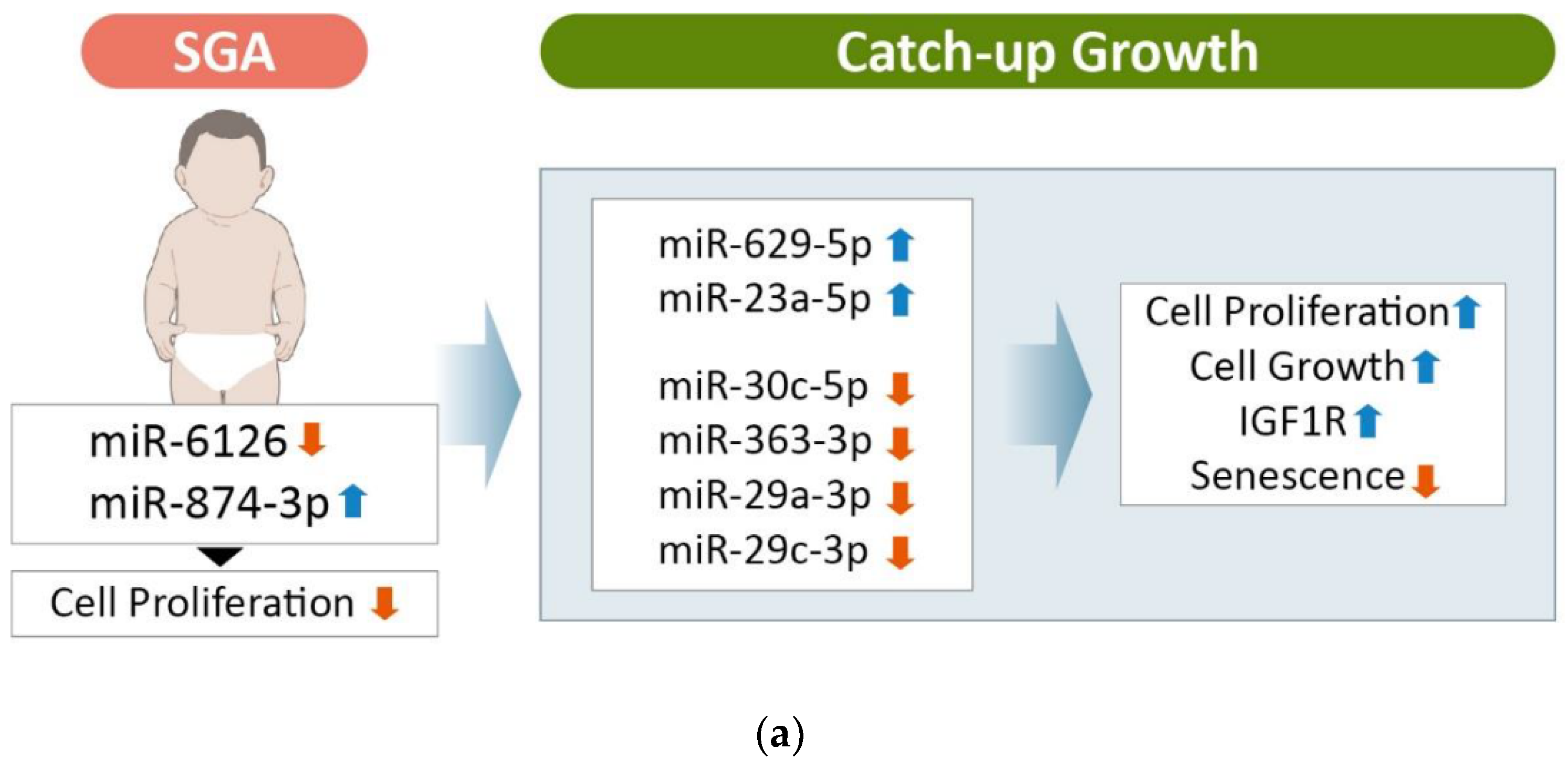
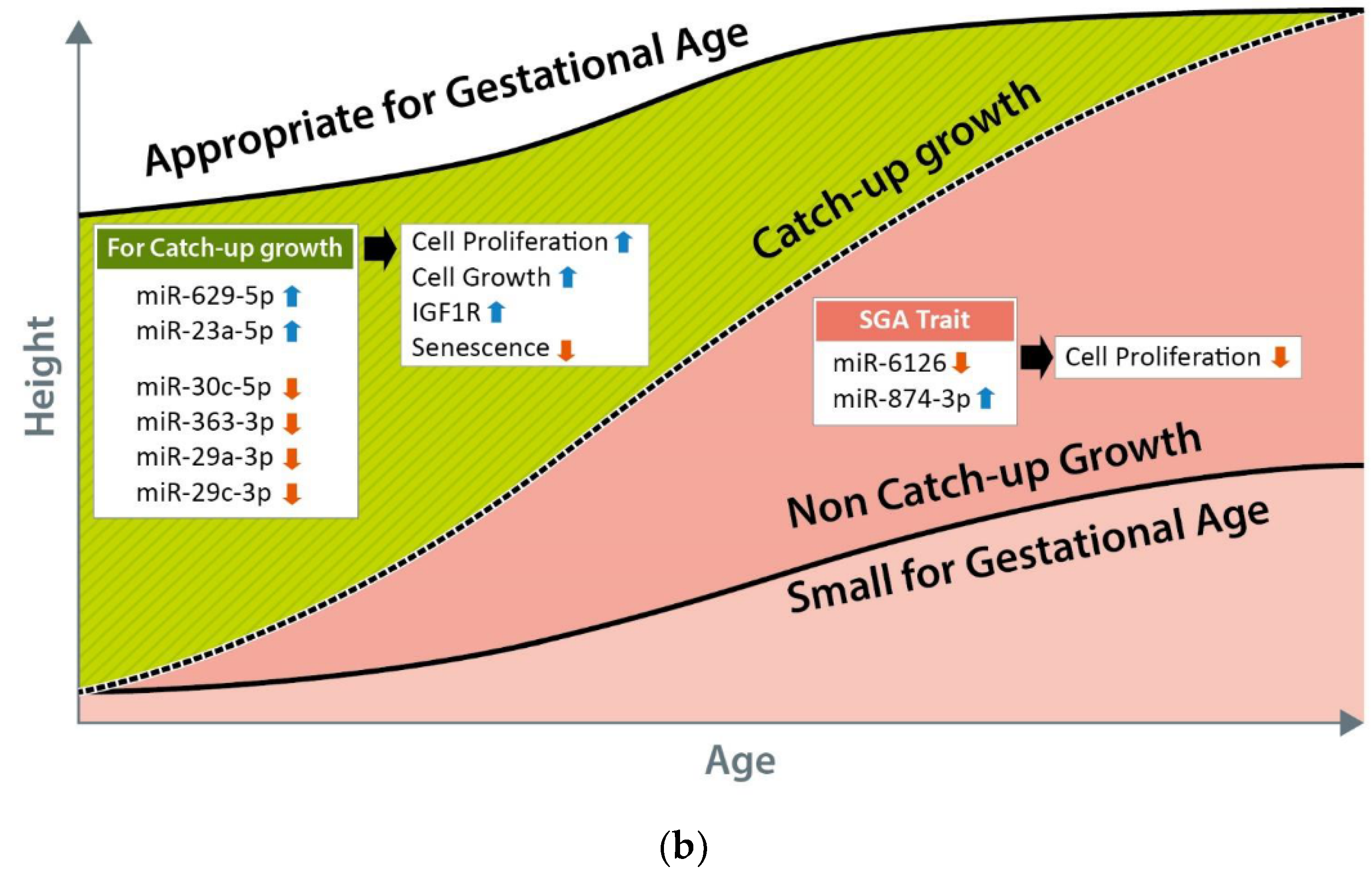
3.5. Comparison of the Exosomal miRNA Profiles of AGA, SGA-CU, and SGA-nCU Children
4. Discussion
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Clayton, P.; Cianfarani, S.; Czernichow, P.; Johannsson, G.; Rapaport, R.; Rogol, A. Management of the Child Born Small for Gestational Age through to Adulthood: A Consensus Statement of the International Societies of Pediatric Endocrinology and the Growth Hormone Research Society. J. Clin. Endocrinol. Metab. 2007, 92, 804–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battaglia, F.C.; Lubchenco, L.O. A practical classification of newborn infants by weight and gestational age. J. Pediatr. 1967, 71, 159–163. [Google Scholar] [CrossRef]
- Finken, M.J.J.; Van Der Steen, M.; Smeets, C.C.J.; Walenkamp, M.J.E.; De Bruin, C.; Hokken-Koelega, A.C.S.; Wit, J.M. Children Born Small for Gestational Age: Differential Diagnosis, Molecular Genetic Evaluation, and Implications. Endocr. Rev. 2018, 39, 851–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campisi, S.C.; Carbone, S.; Zlotkin, S. Catch-Up Growth in Full-Term Small for Gestational Age Infants: A Systematic Review. Adv. Nutr. Int. Rev. J. 2019, 10, 104–111. [Google Scholar] [CrossRef]
- Stalman, S.E.; Solanky, N.; Ishida, M.; Alemán-Charlet, C.; Abu-Amero, S.; Alders, M.; Alvizi, L.; Baird, W.; Demetriou, C.; Henneman, P.; et al. Genetic Analyses in Small-for-Gestational-Age Newborns. J. Clin. Endocrinol. Metab. 2018, 103, 917–925. [Google Scholar] [CrossRef]
- Lee, A.C.; Kozuki, N.; Cousens, S.; Stevens, G.A.; Blencowe, H.; Silveira, M.F.; Sania, A.; Rosen, H.E.; Schmiegelow, C.; Adair, L.S.; et al. Estimates of burden and consequences of infants born small for gestational age in low and middle income countries with INTERGROWTH-21(st) standard: Analysis of CHERG datasets. BMJ 2017, 358, j3677. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Kim, D.H.; Lim, J.S. Growth status of children and adolescents born small for gestational age at full term in Korea: Data from the KNHANES-V. J. Pediatr. Endocrinol. Metab. 2020, 33, 743–750. [Google Scholar] [CrossRef]
- Van Wassenaer, A. Neurodevelopmental consequences of being born SGA. Pediatr. Endocrinol. Rev. 2005, 2, 372–377. [Google Scholar]
- Milovanovic, I.; Njuieyon, F.; Deghmoun, S.; Chevenne, D.; Levy-Marchal, C.; Beltrand, J. SGA Children with Moderate Catch-Up Growth Are Showing the Impaired Insulin Secretion at the Age of 4. PLoS ONE 2014, 9, e100337. [Google Scholar] [CrossRef]
- Hong, Y.H.; Chung, S. Small for gestational age and obesity related comorbidities. Ann. Pediatr. Endocrinol. Metab. 2018, 23, 4–8. [Google Scholar] [CrossRef]
- Verkauskiene, R.; Petraitiene, I.; Wikland, K.A. Puberty in Children Born Small for Gestational Age. Horm. Res. Paediatr. 2013, 80, 69–77. [Google Scholar] [CrossRef]
- Lee, P.A.; Chernausek, S.D.; Hokken-Koelega, A.C.S.; Czernichow, P.; The International SGA Advisory Board. International Small for Gestational Age Advisory Board Consensus Development Conference Statement: Management of Short Children Born Small for Gestational Age, April 24–October 1, 2001. Pediatrics 2003, 111, 1253–1261. [Google Scholar] [CrossRef]
- Karlberg, J.; Albertsson-Wikland, K.; Kwan, E.; Lam, B.; Low, L. The Timing of Early Postnatal Catch-Up Growth in Normal, Full-Term Infants Born Short for Gestational Age. Horm. Res. 1997, 48, 17–24. [Google Scholar] [CrossRef]
- Paz, I.; Seidman, D.S.; Danon, Y.L.; Laor, A.; Stevenson, D.K.; Gale, R. Are children born small for gestational age at increased risk of short stature? Am. J. Dis. Child. 1993, 147, 337–339. [Google Scholar] [CrossRef]
- Karlberg, J.; Albertsson-Wikland, K. Growth in Full-Term Small-for-Gestational-Age Infants: From Birth to Final Height. Pediatr. Res. 1995, 38, 733–739. [Google Scholar] [CrossRef] [Green Version]
- Van Dijk, M.; Mulder, P.; Houdijk, M.; Mulder, J.; Noordam, K.; Odink, R.J.; Rongen-Westerlaken, C.; Voorhoeve, P.; Waelkens, J.; Stokvis-Brantsma, J.; et al. High serum levels of growth hormone (GH) and insulin-like growth factor-I (IGF-I) during high-dose GH treatment in short children born small for gestational age. J. Clin. Endocrinol. Metab. 2006, 91, 1390–1396. [Google Scholar] [CrossRef] [Green Version]
- Iñiguez, G.; Ong, K.; Bazaes, R.; Avila, A.; Salazar, T.; Dunger, D.; Mericq, V. Longitudinal Changes in Insulin-Like Growth Factor-I, Insulin Sensitivity, and Secretion from Birth to Age Three Years in Small-for-Gestational-Age Children. J. Clin. Endocrinol. Metab. 2006, 91, 4645–4649. [Google Scholar] [CrossRef] [Green Version]
- Gat-Yablonski, G.; Phillip, M. Nutritionally-Induced Catch-Up Growth. Nutrients 2015, 7, 517–551. [Google Scholar] [CrossRef] [Green Version]
- Karolina, D.S.; Tavintharan, S.; Armugam, A.; Sepramaniam, S.; Pek, S.L.T.; Wong, M.T.K.; Lim, S.C.; Sum, C.F.; Jeyaseelan, K. Circulating miRNA Profiles in Patients with Metabolic Syndrome. J. Clin. Endocrinol. Metab. 2012, 97, E2271–E2276. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Guo, J.; Zhang, Y.; Chen, J.; Guo, X.; et al. Characterization of microRNAs in serum: A novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef] [Green Version]
- Marques-Rocha, J.L.; Samblas-García, M.; Milagro, F.I.; Bressan, J.; Martínez, J.A.; Marti, A. Noncoding RNAs, cytokines, and inflammation-related diseases. FASEB J. 2015, 29, 3595–3611. [Google Scholar] [CrossRef] [Green Version]
- Galasso, M.; Sana, M.E.; Volinia, S. Non-coding RNAs: A key to future personalized molecular therapy? Genome Med. 2010, 2, 12. [Google Scholar] [CrossRef] [Green Version]
- Mas-Parés, B.; Xargay-Torrent, S.; Bonmatí, A.; Lizarraga-Mollinedo, E.; Martínez-Calcerrada, J.M.; Carreras-Badosa, G.; Prats-Puig, A.; de Zegher, F.; Ibáñez, L.; López-Bermejo, A.; et al. Umbilical cord microRNA in small-for-gestational age children and association with catch-up growth: A pilot study. J. Clin. Endocrinol. Metab. 2019, 104, 5285–5298. [Google Scholar] [CrossRef]
- Marzano, F.; Faienza, M.F.; Caratozzolo, M.F.; Brunetti, G.; Chiara, M.; Horner, D.S.; Annese, A.; D’Erchia, A.M.; Consiglio, A.; Pesole, G.; et al. Pilot study on circulating miRNA signature in children with obesity born small for gestational age and appropriate for gestational age. Pediatr. Obes. 2018, 13, 803–811. [Google Scholar] [CrossRef]
- Lim, J.S.; Lim, S.W.; Ahn, J.H.; Song, B.S.; Shim, K.S.; Hwang, I.T. New Korean reference for birth weight by gestational age and sex: Data from the Korean Statistical Information Service (2008–2012). Ann. Pediatr. Endocrinol. Metab. 2014, 19, 146–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inzaghi, E.; Kistner, A.; Germani, D.; Deodati, A.; Vanpee, M.; Legnevall, L.; Berinder, K.; Cianfarani, S. A prospective case-control study on miRNA circulating levels in subjects born small for gestational age (SGA) evaluated from childhood into young adulthood. PLoS ONE 2020, 15, e0228075. [Google Scholar] [CrossRef] [Green Version]
- Onyshchenko, K.V.; Voitsitskyi, T.V.; Grygorenko, V.M.; Saidakova, N.O.; Pereta, L.V.; Onyschuk, A.P.; Skrypkina, I.Y. Expression of micro-RNA hsa-miR-30c-5p and hsa-miR-138-1 in renal cell carcinoma. Exp. Oncol. 2020, 42, 115–119. [Google Scholar]
- Song, S.; Long, M.; Yu, G.; Cheng, Y.; Yang, Q.; Liu, J.; Wang, Y.; Sheng, J.; Wang, L.; Wang, Z.; et al. Urinary exosome miR-30c-5p as a biomarker of clear cell renal cell carcinoma that inhibits progression by targeting HSPA5. J. Cell. Mol. Med. 2019, 23, 6755–6765. [Google Scholar] [CrossRef]
- Gao, B.-H.; Wu, H.; Wang, X.; Ji, L.-L.; Chen, C. MiR-30c-5p inhibits high glucose-induced EMT and renal fibrogenesis by down-regulation of JAK1 in diabetic nephropathy. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 1338–1349. [Google Scholar]
- Gao, C.; Qian, H.; Shi, Q.; Zhang, H. MicroRNA-363-3p serves as a diagnostic biomarker of acute myocardial infarction and regulates vascular endothelial injury by targeting KLF2. Cardiovasc. Diagn. Ther. 2020, 10, 421–430. [Google Scholar] [CrossRef]
- Zheng, J.; Zhu, S.; Xu, H.; Li, J.; Tang, H.; Zhou, Y.; Huang, Z.; Liu, G. miR-363-3p inhibits rat lung alveolar type II cell proliferation by downregulating STRA6 expression and induces cell apoptosis via cellular oxidative stress and G1-phase cell cycle arrest. Transl. Pediatr. 2021, 10, 2095–2105. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, T.; Huang, H.; Jiang, Y.; Yang, L.; Lin, Z.; He, H.; Liu, T.; Wu, B.; Chen, J.; et al. miR-363-3p inhibits tumor growth by targeting PCNA in lung adenocarcinoma. Oncotarget 2017, 8, 20133–20144. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Liu, J.; Hu, H.; Duan, Q.; Chen, J.; Li, L. MiR-363-3p attenuates neonatal hypoxic-ischemia encephalopathy by targeting DUSP5. Neurosci. Res. 2021, 171, 103–113. [Google Scholar] [CrossRef]
- Dong, J.; Geng, J.; Tan, W. MiR-363-3p suppresses tumor growth and metastasis of colorectal cancer via targeting SphK2. Biomed. Pharmacother. 2018, 105, 922–931. [Google Scholar] [CrossRef]
- Wang, X.; Liu, S.; Cao, L.; Zhang, T.; Yue, D.; Wang, L.; Ping, Y.; He, Q.; Zhang, C.; Wang, M.; et al. miR-29a-3p suppresses cell proliferation and migration by downregulating IGF1R in hepatocellular carcinoma. Oncotarget 2017, 8, 86592–86603. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Zhang, W.; Wu, Z.; Liu, Y.; Shi, Y.; Gong, J.; Shen, W.; Liu, C. miR-29c-3p regulates DNMT3B and LATS1 methylation to inhibit tumor progression in hepatocellular carcinoma. Cell Death Dis. 2019, 10, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.-W.; Wang, E.-W.; Li, L.-X.; Yi, S.-H.; Li, L.-C.; Xu, F.-L.; Wang, D.-L.; Wu, Y.-Z.; Nian, W.-Q. A regulatory loop involving miR-29c and Sp1 elevates the TGF-β1 mediated epithelial-to-mesenchymal transition in lung cancer. Oncotarget 2016, 7, 85905–85916. [Google Scholar] [CrossRef] [Green Version]
- Niu, M.; Gao, D.; Wen, Q.; Wei, P.; Pan, S.; Shuai, C.; Ma, H.; Xiang, J.; Li, Z.; Fan, S.; et al. MiR-29c regulates the expression of miR-34c and miR-449a by targeting DNA methyltransferase 3a and 3b in nasopharyngeal carcinoma. BMC Cancer 2016, 16, 218. [Google Scholar] [CrossRef] [Green Version]
- Shang, J.; Yao, Y.; Fan, X.; Shangguan, L.; Li, J.; Liu, H.; Zhou, Y. miR-29c-3p promotes senescence of human mesenchymal stem cells by targeting CNOT6 through p53–p21 and p16–pRB pathways. Biochim. Biophys. Acta 2016, 1863, 520–532. [Google Scholar] [CrossRef]
- Tao, X.; Yang, X.; Wu, K.; Yang, L.; Huang, Y.; Jin, Q.; Chen, S. miR-629–5p promotes growth and metastasis of hepatocellular carcinoma by activating β-catenin. Exp. Cell Res. 2019, 380, 124–130. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, H.; Fan, L.; Mou, J.; Yin, Y.; Peng, C.; Chen, Y.; Lu, H.; Zhao, L.; Tao, Z.; et al. MiR-629-5p promotes the invasion of lung adenocarcinoma via increasing both tumor cell invasion and endothelial cell permeability. Oncogene 2020, 39, 3473–3488. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Lu, S.; Li, J.; Yu, Q.; Liu, L.; Li, Q. MiR-629-5p promotes colorectal cancer progression through targetting CXXC finger protein 4. Biosci. Rep. 2018, 38, BSR20180613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, L.; Shen, Z.; Qian, H.; Zhou, S.; Chen, Y. Knockdown of miR-629 Inhibits Ovarian Cancer Malignant Behaviors by Targeting Testis-Specific Y-Like Protein 5. DNA Cell Biol. 2017, 36, 1108–1116. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Li, Q.; Wu, J.; Hu, W.; Jiang, J.; Shi, L.; Yang, X.; Zhu, D.; Ji, M.; Wu, C. MiR-629 promotes human pancreatic cancer progression by targeting FOXO3. Cell Death Dis. 2017, 8, e3154. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Jiang, R.; Chu, X.; Wang, F.; Sun, X.; Wang, Y.; Pang, L. Overexpression of microRNA -23a-5p induces myocardial infarction by promoting cardiomyocyte apoptosis through inhibited of PI3K / AKT signalling pathway. Cell Biochem. Funct. 2020, 38, 1047–1055. [Google Scholar] [CrossRef]
- Kurkewich, J.L.; Hansen, J.; Klopfenstein, N.; Zhang, H.; Wood, C.; Boucher, A.; Hickman, J.; Muench, D.E.; Grimes, H.L.; Dahl, R. The miR-23a~27a~24-2 microRNA cluster buffers transcription and signaling pathways during hematopoiesis. PLoS Genet. 2017, 13, e1006887. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Tan, H.-Y.; Feng, Y.-G.; Zhang, C.; Chen, F.; Feng, Y. microRNA-23a in Human Cancer: Its Roles, Mechanisms and Therapeutic Relevance. Cancers 2018, 11, 7. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Huang, Y.; Gu, J.; Zhang, J.; Yang, J.; Liu, S.; Xie, C.; Fan, Y.; Wang, H. miR-23a-5p inhibits cell proliferation and invasion in pancreatic ductal adenocarcinoma by suppressing ECM1 expression. Am. J. Transl. Res. 2019, 11, 2983–2994. [Google Scholar]
- Ganesan, S.; Palani, H.K.; Lakshmanan, V.; Balasundaram, N.; Abu Alex, A.; David, S.; Venkatraman, A.; Korula, A.; George, B.; Balasubramanian, P.; et al. Stromal cells downregulate miR-23a-5p to activate protective autophagy in acute myeloid leukemia. Cell Death Dis. 2019, 10, 1–14. [Google Scholar] [CrossRef]
- Quan, J.; Jin, L.; Pan, X.; He, T.; Lai, Y.; Chen, P.; Lin, C.; Yang, S.; Zeng, H.; Lai, Y. Oncogenic miR-23a-5p is associated with cellular function in RCC. Mol. Med. Rep. 2017, 16, 2309–2317. [Google Scholar] [CrossRef] [Green Version]
- Stanczyk, J.; Pedrioli, D.M.; Brentano, F.; Sanchez-Pernaute, O.; Kolling, C.; Gay, R.E.; Detmar, M.; Gay, S.; Kyburz, D. Altered expression of MicroRNA in synovial fibroblasts and synovial tissue in rheumatoid arthritis. Arthritis Rheum. 2008, 58, 1001–1009. [Google Scholar] [CrossRef]
- Yang, J.-X.; Xie, P.; Li, Y.-S.; Wen, T.; Yang, X.-C. Osteoclast-derived miR-23a-5p-containing exosomes inhibit osteogenic differentiation by regulating Runx2. Cell. Signal. 2019, 70, 109504. [Google Scholar] [CrossRef]
- Li, Y.; Chen, X.; Xue, W.; Liang, J.; Wang, L. MiR-874 Inhibits Cell Proliferation, Migration, and Invasion of Glioma Cells and Correlates with Prognosis of Glioma Patients. NeuroMolecular Med. 2020, 23, 247–255. [Google Scholar] [CrossRef]
- Kanlikilicer, P.; Rashed, M.R.; Bayraktar, R.; Mitra, R.; Ivan, C.; Aslan, B.; Zhang, X.; Filant, J.; Silva, A.M.; Rodriguez-Aguayo, C.; et al. Ubiquitous Release of Exosomal Tumor Suppressor miR-6126 from Ovarian Cancer Cells. Cancer Res. 2016, 76, 7194–7207. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total (n = 10) | SGA-nCU | AGA | p-Value |
|---|---|---|---|
| n (%) | 5 (50%) | 5 (50%) | - |
| Boys/girls | 3/2 | 2/3 | - |
| GA (weeks) | 38.80 ± 1.64 | 39.40 ± 1.34 | 0.548 |
| Birth weight (kg) | 2.44 ± 0.18 | 3.34 ± 0.27 | 0.008 |
| CA (year) | 4.54 ± 0.28 | 4.63 ± 0.28 | 0.690 |
| Height (cm) | 97.22 ± 1.96 | 106.94 ± 1.66 | 0.008 |
| Height SDS | –2.17 ± 0.19 | 0.15 ± 0.45 | 0.008 |
| Weight (kg) | 13.62 ± 1.62 | 17.30 ± 1.83 | 0.008 |
| Weight SDS | –2.51 ± 1.01 | –0.28 ± 0.79 | 0.008 |
| BMI SDS | –1.26 ± 1.15 | –0.62 ± 1.15 | 0.690 |
| Total (n = 16) | SGA | AGA | p-Value | |
|---|---|---|---|---|
| SGA-nCU | SGA-CU | |||
| n (%) | 6 (37.5) | 5 (31.25) | 5 (31.25) | - |
| Boys/girls | 3/3 | 2/3 | 2/3 | - |
| GA (weeks) | 38.60 ± 1.36 | 38.20 ± 1.09 | 39.20 ± 1.30 | 0.353 |
| Birth weight (kg) | 2.39 ± 0.24 | 2.06 ± 0.23 | 3.47 ± 0.29 | 0.003 |
| CA (year) | 4.44 ± 0.31 | 5.93 ± 0.77 | 5.87 ± 1.06 | 0.018 |
| Height (cm) | 96.66 ± 1.89 | 111.66 ± 3.78 | 117.58 ± 7.92 | 0.004 |
| Height SDS | –2.10 ± 0.27 | –0.61 ± 0.39 | 0.69 ± 0.82 | 0.001 |
| Weight (kg) | 14.16 ± 1.74 | 17.96 ± 1.88 | 25.50 ± 7.42 | 0.003 |
| Weight SDS | –2.06 ± 1.14 | –1.17 ± 0.67 | 1.19 ± 1.15 | 0.006 |
| BMI SDS | –0.62 ± 1.21 | –1.17 ± 0.85 | 1.12 ± 1.52 | 0.025 |
| miRNA | Fold Change SGA-nCU/AGA | p-Value |
|---|---|---|
| hsa-miR-29b-3p | 7.03 | <0.001 |
| hsa-miR-4448 | 5.34 | 0.000 |
| hsa-miR-141-3p | 4.97 | 0.001 |
| hsa-miR-29a-3p | 4.18 | 0.004 |
| hsa-miR-144-5p | 4.14 | 0.003 |
| hsa-miR-26b-5p | 4.00 | 0.006 |
| hsa-miR-29c-3p | 3.93 | 0.008 |
| hsa-let-7g-5p | 3.81 | 0.002 |
| hsa-miR-32-5p | 3.48 | 0.011 |
| hsa-miR-96-5p | 3.45 | 0.012 |
| hsa-miR-543 | –2.56 | 0.031 |
| hsa-miR-493-3p | –2.64 | 0.002 |
| hsa-miR-125b-1-3p | –2.66 | 0.017 |
| hsa-miR-409-3p | –2.86 | 0.013 |
| hsa-miR-3120-5p | –2.90 | 0.030 |
| hsa-miR-485-3p | –2.92 | 0.032 |
| hsa-miR-758-3p | –2.98 | 0.002 |
| hsa-miR-654-5p | –3.30 | 0.002 |
| hsa-miR-370-3p | –3.35 | 0.005 |
| hsa-miR-127-3p | –3.49 | 0.005 |
| miRNA | Fold Change SGA-nCU/AGA | p-Value |
|---|---|---|
| hsa-miR-3651 | 11.17 | 0.006 |
| hsa-miR-874-3p | 3.65 | 0.002 |
| hsa-miR-363-3p | 3.37 | 0.007 |
| hsa-miR-29a-3p | 2.44 | 0.049 |
| hsa-miR-15a-5p | 2.44 | 0.043 |
| hsa-miR-29c-3p | 2.43 | 0.009 |
| hsa-miR-30c-5p | 2.09 | 0.043 |
| hsa-miR-505-5p | –2.05 | 0.039 |
| hsa-let-7d-5p | –2.07 | 0.044 |
| hsa-miR-629-5p | –2.24 | 0.040 |
| hsa-miR-6126 | –2.35 | 0.049 |
| hsa-let-7f-5p | –2.78 | 0.033 |
| hsa-let-7a-5p | –2.87 | 0.029 |
| hsa-let-7e-5p | –2.93 | 0.007 |
| hsa-miR-23a-5p | –3.25 | 0.004 |
| hsa-miR-28-5p | –4.80 | 0.002 |
| hsa-miR-1-3p | –4.80 | 0.004 |
| (a) | ||||
|---|---|---|---|---|
| miRNA | Fold Change SGA-CU/AGA | p-Value | ||
| hsa-miR-1285-5p | 3.90 | 0.006 | ||
| hsa-miR-874-3p | 3.29 | 0.004 | ||
| hsa-miR-339-3p | 3.06 | 0.027 | ||
| hsa-miR-143-3p | 2.08 | 0.038 | ||
| hsa-miR-6126 | –2.12 | 0.027 | ||
| hsa-miR-627-5p | –4.26 | 0.044 | ||
| (b) | ||||
| SGA-nCU vs. AGA | SGA-CU vs. AGA | |||
| miRNA | Fold Change | p-Value | Fold Change | p-Value |
| hsa-miR-874-3p | 3.65 | 0.002 | 3.29 | 0.004 |
| hsa-miR-6126 | –2.35 | 0.049 | –2.12 | 0.027 |
| (a) | ||||
|---|---|---|---|---|
| miRNA | Fold Change SGA-nCU/SGA-CU | p-Value | ||
| hsa-miR-342-3p | 3.59 | 0.010 | ||
| hsa-miR-576-5p | 3.55 | 0.001 | ||
| hsa-miR-150-5p | 3.10 | 0.006 | ||
| hsa-miR-192-5p | 2.85 | 0.001 | ||
| hsa-miR-29b-3p | 2.83 | 0.006 | ||
| hsa-miR-101-3p | 2.65 | 0.035 | ||
| hsa-miR-30c-5p | 2.58 | 0.010 | ||
| hsa-miR-363-3p | 2.45 | 0.046 | ||
| hsa-miR-29a-3p | 2.38 | 0.037 | ||
| hsa-miR-25-3p | 2.22 | 0.011 | ||
| hsa-miR-21-5p | 2.17 | 0.012 | ||
| hsa-miR-29c-3p | 2.03 | 0.027 | ||
| hsa-miR-335-5p | –2.04 | 0.012 | ||
| hsa-miR-574-5p | –2.05 | 0.035 | ||
| hsa-miR-193a-5p | –2.37 | 0.035 | ||
| hsa-miR-629-5p | –2.62 | 0.012 | ||
| hsa-miR-23a-5p | –2.85 | 0.005 | ||
| hsa-miR-382-5p | –2.88 | 0.039 | ||
| hsa-miR-326 | –2.91 | 0.007 | ||
| (b) | ||||
| SGA-nCU vs. SGA-CU | SGA-nCU vs. AGA | |||
| miRNA | Fold Change | p-Value | Fold Change | p-Value |
| hsa-miR-30c-5p | 2.58 | 0.010 | 2.09 | 0.043 |
| hsa-miR-363-3p | 2.45 | 0.046 | 3.37 | 0.007 |
| hsa-miR-29a-3p | 2.38 | 0.037 | 2.44 | 0.049 |
| hsa-miR-29c-3p | 2.03 | 0.027 | 2.43 | 0.009 |
| hsa-miR-629-5p | –2.62 | 0.012 | –2.24 | 0.040 |
| hsa-miR-23a-5p | –2.85 | 0.005 | –3.25 | 0.004 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, H.R.; Han, J.-A.; Kim, H.; Lee, H.J.; Shim, Y.S.; Kang, M.J.; Yoon, J.S.; Ryu, S.; Hwang, I.T. Exosomal miRNA Profile in Small-for-Gestational-Age Children: A Potential Biomarker for Catch-Up Growth. Genes 2022, 13, 938. https://doi.org/10.3390/genes13060938
Jeong HR, Han J-A, Kim H, Lee HJ, Shim YS, Kang MJ, Yoon JS, Ryu S, Hwang IT. Exosomal miRNA Profile in Small-for-Gestational-Age Children: A Potential Biomarker for Catch-Up Growth. Genes. 2022; 13(6):938. https://doi.org/10.3390/genes13060938
Chicago/Turabian StyleJeong, Hwal Rim, Jae-A Han, Heeji Kim, Hye Jin Lee, Young Suk Shim, Min Jae Kang, Jong Seo Yoon, Seongho Ryu, and Il Tae Hwang. 2022. "Exosomal miRNA Profile in Small-for-Gestational-Age Children: A Potential Biomarker for Catch-Up Growth" Genes 13, no. 6: 938. https://doi.org/10.3390/genes13060938
APA StyleJeong, H. R., Han, J. -A., Kim, H., Lee, H. J., Shim, Y. S., Kang, M. J., Yoon, J. S., Ryu, S., & Hwang, I. T. (2022). Exosomal miRNA Profile in Small-for-Gestational-Age Children: A Potential Biomarker for Catch-Up Growth. Genes, 13(6), 938. https://doi.org/10.3390/genes13060938