
The Effect of Host miRNAs on Prognosis in COVID-19: miRNA-155 May Promote Severity via Targeting Suppressor of Cytokine Signaling 1 (SOCS1) Gene

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
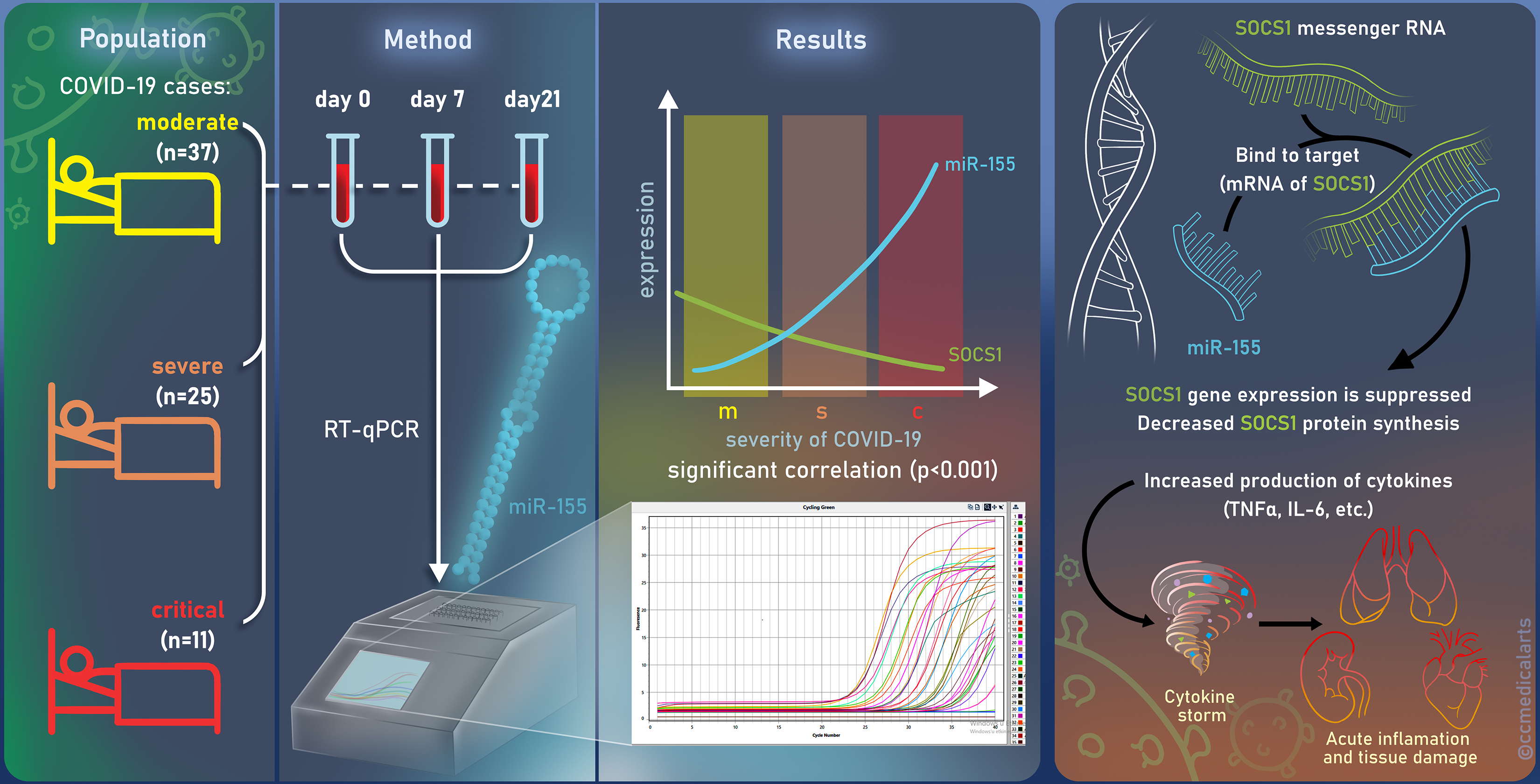
2.1. Patients
2.2. Data and Sample Collection
2.3. Sample Preparation
2.4. Real-Time qPCR Amplification and Detection
2.5. Quantitation of SOCS1 Expression
2.6. Statistical Analysis
3. Results
3.1. Demographic Characteristic and Laboratory Findings of Patients
3.2. Quantitative RT-PCR Analyses of miR-155-5p and SOCS1 Expression in the Moderate, Severe, and Critical COVID-19 Patients
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Farr, R.J.; Rootes, C.L.; Rowntree, L.C.; Nguyen, T.H.; Hensen, L.; Kedzierski, L.; Stewart, C.R. Altered microRNA expression in COVID-19 patients enables identification of SARS-CoV-2 infection. PLoS Pathog. 2021, 17, e1009759. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Barbu, M.G.; Condrat, C.E.; Thompson, D.C.; Bugnar, O.L.; Cretoiu, D.; Toader, O.D.; Suciu, N.; Voinea, S.C. MicroRNA Involvement in Signaling Pathways during Viral Infection. Front. Cell Dev. Biol. 2020, 8, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinilla, L.; Benitez, I.D.; González, J.; Torres, G.; Barbé, F.; de Gonzalo-Calvo, D. Peripheral blood microRNAs and the COVID-19 patient: Methodological considerations, technical challenges and practice points. RNA Biol. 2021, 18, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kowdley, K.V. MicroRNAs in Common Human Diseases. Genom. Proteom. Bioinform. 2012, 10, 246–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, F.R.; Tang, L.J.; He, Y.; Garcia, R.C. An update on the role of miRNA-155 in pathogenic microbial infections. Microbes Infect. 2015, 17, 613–621. [Google Scholar] [CrossRef]
- Yang, S.; Li, F.; Jia, S.; Zhang, K.; Jiang, W.; Shang, Y.; Chen, M. Early secreted antigen ESAT-6 of Mycobacterium tuberculosis promotes apoptosis of macrophages via targeting the microRNA155-SOCS1 interaction. Cell. Physiol. Biochem. 2015, 35, 1276–1288. [Google Scholar] [CrossRef]
- Gangemi, S.; Tonacci, A. AntagomiRs: A novel therapeutic strategy for challenging COVID-19 cytokine storm. Cytokine Growth Factor Rev. 2021, 58, 111–113. [Google Scholar] [CrossRef]
- Epigenetic Tools as Prognostic Predictors in Covid19. Available online: https://www.clinicaltrials.gov/ct2/show/NCT04411563 (accessed on 24 May 2022).
- COVID-19 Treatment Guidelines Panel. Coronavirus Disease 2019 (COVID-19) Treatment Guidelines. National Institutes of Health. Available online: https://www.covid19treatmentguidelines.nih.gov/ (accessed on 25 December 2021).
- Tuck, M.K.; Chan, D.W.; Chia, D.; Godwin, A.K.; Grizzle, W.E.; Krueger, K.E.; Rom, W.; Sanda, M.; Sorbara, L.; Stass, S.; et al. Standard operating procedures for serum and plasma collection: Early detection research network consensus statement standard operating procedure integration working group. J. Proteome Res. 2009, 8, 113–117. [Google Scholar] [CrossRef] [Green Version]
- Bustin, S.A. How to speed up the polymerase chain reaction. Biomol. Detect. Quantif. 2017, 20, 10–14. [Google Scholar] [CrossRef]
- Hong, L.Z.; Zhou, L.; Zou, R.; Khoo, C.M.; Chew, A.L.S.; Chin, C.L.; Shih, S.J. Systematic evaluation of multiple qPCR platforms, NanoString and miRNA-Seq for microRNA biomarker discovery in human biofluids. Sci. Rep. 2021, 11, 4435. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cheng, X.; Dong, Q.; Zhou, C.; Wang, Y.; Song, B.; Ba, Y. The characteristics of laboratory tests at admission and the risk factors for adverse clinical outcomes of severe and critical COVID-19 patients. BMC Infect. Dis. 2021, 21, 371. [Google Scholar] [CrossRef]
- Wu, J.; Wu, X.; Zeng, W.; Guo, D.; Fang, Z.; Chen, L.; Li, C. Chest CT findings in patients with coronavirus disease 2019 and its relationship with clinical features. Investig. Radiol. 2020, 55, 257. [Google Scholar] [CrossRef]
- Poblador-Plou, B.; Carmona-Pírez, J.; Ioakeim-Skoufa, I.; Poncel-Falco, A.; Bliek-Bueno, K.; Cano-del Pozo, M.; EpiChron Group. Baseline chronic comorbidity and mortality in laboratory-confirmed COVID-19 cases: Results from the PRECOVID study in Spain. Int. J. Environ. Res. Public Health 2020, 17, 5171. [Google Scholar] [CrossRef]
- Girardi, E.; López, P.; Pfeffer, S. On the importance of host microRNAs during viral infection. Front. Genet. 2018, 9, 439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leon-Icaza, S.A.; Zeng, M.; Rosas-Taraco, A.G. microRNAs in viral acute respiratory infections: Immune regulation, biomarkers, therapy, and vaccines. ExRNA 2019, 1, 1. [Google Scholar] [CrossRef] [Green Version]
- Inchley, C.S.; Sonerud, T.; Fjærli, H.O.; Nakstad, B. Nasal mucosal microRNA expression in children with respiratory syncytial virus infection. BMC Infect. Dis. 2015, 15, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Choi, E.J.; Lee, I.; Lee, Y.S.; Bao, X. Non-coding RNAs and their role in respiratory syncytial virus (RSV) and human metapneumovirus (hMPV) infections. Viruses 2020, 12, 345. [Google Scholar] [CrossRef] [Green Version]
- Garg, A.; Seeliger, B.; Derda, A.A.; Xiao, K.; Gietz, A.; Scherf, K.; Thum, T. Circulating cardiovascular microRNAs in critically ill COVID-19 patients. Eur. J. Heart Fail. 2021, 23, 468–475. [Google Scholar] [CrossRef]
- Donyavi, T.; Bokharaei-Salim, F.; Baghi, H.B.; Khanaliha, K.; Janat-Makan, M.A.; Karimi, B.; Kiani, S.J. Acute and post-acute phase of COVID-19: Analyzing expression patterns of miRNA-29a-3p, 146a-3p, 155–5p, and let-7b-3p in PBMC. Int. Immunopharmacol. 2021, 97, 107641. [Google Scholar] [CrossRef] [PubMed]
- Abbasi-Kolli, M.; Nahand, J.S.; Kiani, S.J.; Khanaliha, K.; Khatami, A.; Taghizadieh, M.; Bokharaei-Salim, F. The expression patterns of MALAT-1, NEAT-1, THRIL, and miR-155-5p in the acute to the post-acute phase of COVID-19 disease. Braz. J. Infect. Dis. 2022, 26, 102354. [Google Scholar] [CrossRef]
- Visacri, M.B.; Nicoletti, A.S.; Pincinato, E.C.; Loren, P.; Saavedra, N.; Saavedra, K.; Moriel, P. Role of miRNAs as biomarkers of COVID-19: A scoping review of the status and future directions for research in this field. Biomark. Med. 2021, 15, 1785–1795. [Google Scholar] [CrossRef] [PubMed]
- Kassif-Lerner, R.; Zloto, K.; Rubin, N.; Asraf, K.; Doolman, R.; Paret, G.; Nevo-Caspi, Y. miR-155: A Potential Biomarker for Predicting Mortality in COVID-19 Patients. J. Pers. Med. 2022, 12, 324. [Google Scholar] [CrossRef] [PubMed]
- Pairo-Castineira, E.; Clohisey, S.; Klaric, L.; Bretherick, A.D.; Rawlik, K.; Pasko, D.; Baillie, J.K. Genetic mechanisms of critical illness in COVID-19. Nature 2021, 591, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Gemmati, D.; Bramanti, B.; Serino, M.L.; Secchiero, P.; Zauli, G.; Tisato, V. COVID-19 and individual genetic susceptibility/receptivity: Role of ACE1/ACE2 genes, immunity, inflammation and coagulation. Might the double X-chromosome in females be protective against SARS-CoV-2 compared to the single X-chromosome in males? Int. J. Mol. Sci. 2020, 21, 3474. [Google Scholar] [CrossRef]
- Velavan, T.P.; Pallerla, S.R.; Rüter, J.; Augustin, Y.; Kremsner, P.G.; Krishna, S.; Meyer, C.G. Host genetic factors determining COVID-19 susceptibility and severity. EBioMedicine 2021, 72, 103629. [Google Scholar] [CrossRef]
- Zhang, S.; Amahong, K.; Sun, X.; Lian, X.; Liu, J.; Sun, H.; Qiu, Y. The miRNA: A small but powerful RNA for COVID-19. Brief. Bioinform. 2021, 22, 1137–1149. [Google Scholar] [CrossRef]
- Li, C.; Hu, X.; Li, L.; Li, J.H. Differential microRNA expression in the peripheral blood from human patients with COVID-19. J. Clin. Lab. Anal. 2020, 34, e23590. [Google Scholar] [CrossRef]
- AbdelHamid, S.G.; Refaat, A.A.; Benjamin, A.M.; Elmawardy, L.A.; Elgendy, L.A.; Manolly, M.M.; Hamdy, N.M. Deciphering epigenetic (s) role in modulating susceptibility to and severity of COVID-19 infection and/or outcome: A systematic rapid review. Environ. Sci. Pollut. Res. 2021, 28, 54209–54221. [Google Scholar] [CrossRef]
- Khan, M.; Sany, M.; Us, R.; Islam, M.; Islam, A.B.M.M. Epigenetic regulator miRNA pattern differences among SARS-CoV, SARS-CoV-2, and SARS-CoV-2 world-wide isolates delineated the mystery behind the epic pathogenicity and distinct clinical characteristics of pandemic COVID-19. Front. Genet. 2015, 11, 765. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Tang, M.; Zong, P.; Liu, H.; Zhang, T.; Liu, Y.; Zhao, Y. MiRNA-155 Regulates the Th17/Treg Ratio by Targeting SOCS1 in Severe Acute Pancreatitis. Front. Physiol. 2018, 9, 686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.F.; Gasteiger, G.; Yu, I.S.; Chaudhry, A.; Hsin, J.P.; Lu, Y.; Rudensky, A.Y. A single miRNA-mRNA interaction affects the immune response in a context-and cell-type-specific manner. Immunity 2015, 43, 52–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Variable | Moderate (n = 37) | Severe (n = 25) | Critically Ill (n = 11) | p-Value | |
|---|---|---|---|---|---|
| Age (year) | Mean ± SD | 56.05 ± 13.72 | 59.64 ± 14.84 | 51.27 ± 16.95 | 0.279 * |
| Gender | Female, n (%) | 18 (48.65) | 7 (28.00) | 4 (36.36) | 0.257 + |
| Male, n (%) | 19 (51.35) | 18 (72.00) | 7 (63.64) | ||
| Chest CT Findings | Mild, n (%) | 16 (43.24) | 3 (12.00) | 1 (9.09) | |
| Moderate, n (%) | 17 (45.95) | 7 (28.00) | 3 (27.27) | 0.0001 + | |
| Severe, n (%) | 4 (10.81) | 15 (60.00) | 7 (63.64) | ||
| Patient status 28 days after hospital admission | Discharge, n (%) | 30 (81.08) | 8 (32.00) | 0 (0.00) | |
| Continuing treatment | 6 (16.22) | 17 (68.00) | 8 (72.73) | 0.0001 + | |
| Mortality, n (%) | 1 (2.70) | 0 (0.00) | 3 (27.27) |
| Moderate (n: 37) | Severe (n: 25) | Critically Ill (n: 11) | p-Value | ||
|---|---|---|---|---|---|
| Glucose (mg/dL) | Mean ± SD | 134.77 ± 46.97 | 144.86 ± 63.95 | 161.75 ± 66.43 | 0.368 * |
| Urea (mg/dL) | Mean ± SD | 34.81 ± 20.53 | 38.14 ± 18.74 | 31.68 ± 12.3 | 0.614 * |
| Creatinine (mg/dL) | Mean ± SD | 2.57 ± 10.02 | 0.97 ± 0.31 | 0.83 ± 0.31 | 0.325 ‡ |
| Median (IQR) | 0.85 (0.66–1.02) | 0.94 (0.79–1.13) | 0.74 (0.55–1.04) | ||
| eGFR (mL/min/1.7) | Mean ± SD | 89.46 ± 26.23 | 84 ± 24.74 | 101.64 ± 14.86 | 0.067 ‡ |
| Median (IQR) | 92 (78.5–106) | 81 (63–98) | 97 (89–116) | ||
| AST (U/L) | Mean ± SD | 46.53 ± 32.97 | 60.68 ± 44.1 | 27.9 ± 11.49 | 0.027 ‡ |
| Median (IQR) | 33 (25.4–54.5) | 48 (31.7–76) | 27 (18–35) | ||
| ALT (U/L) | Mean ± SD | 42.69 ± 43.92 | 60.68 ± 60.56 | 23.54 ± 13 | 0.162 ‡ |
| Median (IQR) | 26 (15.75–56.5) | 46.2 (17–78.5) | 19 (12.4–30.5) | ||
| GGT (U/L) | Mean ± SD | 98.93 ± 245.24 | 62.05 ± 52.39 | 71.2 ± 81.38 | 0.861 ‡ |
| Median (IQR) | 42.25 (25.5–80) | 50 (25–76) | 38.8 (14.73–119.43) | ||
| LDH (U/L) | Mean ± SD | 340.64 ± 180.01 | 369.2 ± 151.32 | 321.91 ± 137.81 | 0.398 ‡ |
| Median (IQR) | 282.5 (243–369.5) | 353 (244.5–478) | 274 (208–472) | ||
| CK (U/L) | Mean ± SD | 145.36 ± 145.04 | 380.91 ± 525.99 | 141.31 ± 106.76 | 0.542 ‡ |
| Median (IQR) | 95 (42.25–206.75) | 119 (45–601) | 83 (65–274) | ||
| Lipase (U/L) | Mean ± SD | 43.73 ± 39.33 | 59.59 ± 43.78 | 36.19 ± 48.77 | 0.044 ‡ |
| Median (IQR) | 31.98 (15.13–62.03) | 49.69 (34.42–79.3) | 19.34 (14.29–24) | ||
| Ca (mg/dL) | Mean ± SD | 8.79 ± 0.63 | 8.6 ± 0.63 | 8.86 ± 0.45 | 0.375 * |
| Phosphorus (mg/dL) | Mean ± SD | 3.11 ± 0.73 | 2.9 ± 0.54 | 3.16 ± 0.96 | 0.481 * |
| Magnesium (mg/dL) | Mean ± SD | 1.98 ± 0.27 | 1.91 ± 0.27 | 1.94 ± 0.24 | 0.611 * |
| Ferritin (μg/L) | Mean ± SD | 336.54 ± 335.75 | 539.91 ± 562.68 | 421.14 ± 432.4 | 0.367 ‡ |
| Median (IQR) | 184.3 (88.15–599.1) | 344 (150–820) | 214.35 (136.65–589.8) | ||
| CRP (mg/L) | Mean ± SD | 71.32 ± 76.58 | 117.87 ± 97.12 | 155.61 ± 73.06 | 0.003 ‡ |
| Median (IQR) | 48.54 (15.58–105.11) | 104.1 (32.38–179.37) | 123.16 (107–188) | ||
| Procalcitonin (ng/mL) | Mean ± SD | 0.16 ± 0.19 | 0.38 ± 0.65 | 2.4 ± 0.8 | 0.0001‡ |
| Median (IQR) | 0.08 (0.04–0.2) | 0.13 (0.06–0.525) | 2.115 (2.03–3.19) | ||
| D-dimer (μg FEU/mL) | Mean ± SD | 0.71 ± 0.98 | 2.25 ± 2.54 | 2.09 ± 0.48 | 0.001 ‡ |
| Median (IQR) | 0.34 (0.22–0.55) | 1.41 (0.34–4) | 2.08 (1.69–2.41) | ||
| PT (s) | Mean ± SD | 15.27 ± 4.76 | 14.49 ± 1.51 | 18.37 ± 10.01 | 0.028 ‡ |
| Median (IQR) | 13.75 (12.95–15.35) | 14.9 (13.35–15.45) | 15.65 (14.48–17.3) | ||
| INR | Mean ± SD | 1.21 ± 0.46 | 1.47 ± 0.84 | 1.53 ± 1.21 | 0.042 ‡ |
| Median (IQR) | 1.055 (1.01–1.215) | 1.15 (1.03–1.31) | 1.245 (1.14–1.38) | ||
| aPTT (s) | Mean ± SD | 41.11 ± 15.39 | 49.6 ± 34.03 | 39.81 ± 8.26 | 0.198 ‡ |
| Median (IQR) | 35.4 (32.9–39.4) | 41.3 (34.4–52.5) | 41.2 (31.6–45.4) | ||
| Fibrinogen (mg/dL) | Mean ± SD | 518 ± 316.87 | 469 ± 98 | 713.88 ± 142.66 | 0.002‡ |
| Median (IQR) | 454 (373–555.5) | 467 (398–526) | 719 (576.75–838.5) | ||
| Troponin I (ng/mL) | Mean ± SD | 46.63 ± 211.57 | 17.86 ± 27.58 | 9.09 ± 5.39 | 0.772 ‡ |
| Median (IQR) | 6 (4–13) | 7 (3.75–21) | 9 (3–13) | ||
| WBC (103/µL) | Mean ± SD | 6.97 ± 3.32 | 7.57 ± 2.87 | 10.58 ± 2.29 | 0.014 * |
| Hemoglobin (g/dL) | Mean ± SD | 12.56 ± 1.84 | 12.52 ± 1.81 | 12.96 ± 1.65 | 0.779 * |
| Hematocrit (%) | Mean ± SD | 37.66 ± 4.26 | 37.39 ± 5.52 | 38.99 ± 5.24 | 0.653 * |
| Platelet (103/µL) | Mean ± SD | 230.25 ± 81.87 | 256 ± 130 | 197 ± 42.71 | 0.224 * |
| Neutrophil (103/µL) | Mean ± SD | 4.91 ± 3.33 | 5.53 ± 2.9 | 5.46 ± 2.69 | 0.321 |
| Median (IQR) | 3.77 (2.4–5.9) | 5.1 (3.5–7) | 5.43 (3.3–8) | ||
| Lymphocyte | Mean ± SD | 1.52 ± 0.82 | 1.38 ± 0.73 | 1.11 ± 0.75 | 0.015 * |
| Neu % | Mean ± SD | 65.42 ± 14.17 | 70.04 ± 16.68 | 68.46 ± 16.82 | 0.514 * |
| Lym % | Mean ± SD | 25.01 ± 12.34 | 21.36 ± 14.19 | 22.45 ± 13.7 | 0.479 ‡ |
| Median (IQR) | 24.1 (15.9–31.2) | 21.8 (9.9–27.5) | 21.7 (10.3–29.9) | ||
| Moderate (n: 37) | Severe (n: 25) | Critical (n: 11) | p‡ | |||
|---|---|---|---|---|---|---|
| miR-155-5p | Admission to hospital | Mean ± SD | 2.696 ± 2.162 | 4.748 ± 3.269 | 11.651 ± 2.281 | 0.0001 |
| Median (IQR) | 2.15 (1.56–2.96) | 3.98 (3.04–4.66) | 11.527 (9.65–13.55) | |||
| Day 7 | Mean ± SD | 1.925 ± 1.784 | 2.875 ± 2.572 | 10.656 ± 1.436 | 0.0001 | |
| Median (IQR) | 1.25 (1.09–2.01) | 2.46 (1.61–2.99) | 10.627 (9.67–11.68) | |||
| Day 21 | Mean ± SD | 1.33 ± 2.064 | 1.406 ± 1.039 | 10.044 ± 1.805 | 0.0001 | |
| Median (IQR) | 0.95 (0.31–1.23) | 1.09 (0.95–1.81) | 9.657 (8.65–10.85) | |||
| p† | 0.0001 | 0.0001 | 0.003 | |||
| SOCS1 | Admission to hospital | Mean ± SD | 1.921 ± 0.68 | 1.308 ± 0.468 | 0.472 ± 0.149 | 0.0001 |
| Median (IQR) | 1.81 (1.41–2.58) | 1.39 (1.09–1.7) | 0.42 (0.39–0.53) | |||
| Day 7 | Ort ± SS | 2.452 ± 0.584 | 2.119 ± 0.565 | 0.566 ± 0.15 | 0.0001 | |
| Median (IQR) | 2.65 (2.24–2.82) | 2.16 (1.88–2.54) | 0.584 (0.42–0.7) | |||
| Day 21 | Ort ± SS | 2.885 ± 0.71 | 2.714 ± 0.584 | 0.616 ± 0.168 | 0.0001 | |
| Median (IQR) | 2.98 (2.74–3.18) | 2.89 (2.8–3) | 0.618 (0.5–0.72) | |||
| p† | 0.0001 | 0.0001 | 0.02 |
| miR-155-5p on Admission | miR-155-5p 7th Day | miR-155-5p 21st Day | ||
|---|---|---|---|---|
| SOCS1 on admission | r | −0.805 | ||
| p | 0.0001 | |||
| SOCS1 day 7 | r | −0.940 | ||
| p | 0.0001 | |||
| SOCS1 day 21 | r | −0.933 | ||
| p | 0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gedikbasi, A.; Adas, G.; Isiksacan, N.; Kart Yasar, K.; Canbolat Unlu, E.; Yilmaz, R.; Hergunsel, G.O.; Cukurova, Z. The Effect of Host miRNAs on Prognosis in COVID-19: miRNA-155 May Promote Severity via Targeting Suppressor of Cytokine Signaling 1 (SOCS1) Gene. Genes 2022, 13, 1146. https://doi.org/10.3390/genes13071146
Gedikbasi A, Adas G, Isiksacan N, Kart Yasar K, Canbolat Unlu E, Yilmaz R, Hergunsel GO, Cukurova Z. The Effect of Host miRNAs on Prognosis in COVID-19: miRNA-155 May Promote Severity via Targeting Suppressor of Cytokine Signaling 1 (SOCS1) Gene. Genes. 2022; 13(7):1146. https://doi.org/10.3390/genes13071146
Chicago/Turabian StyleGedikbasi, Asuman, Gokhan Adas, Nilgun Isiksacan, Kadriye Kart Yasar, Esra Canbolat Unlu, Rabia Yilmaz, Gulsum Oya Hergunsel, and Zafer Cukurova. 2022. "The Effect of Host miRNAs on Prognosis in COVID-19: miRNA-155 May Promote Severity via Targeting Suppressor of Cytokine Signaling 1 (SOCS1) Gene" Genes 13, no. 7: 1146. https://doi.org/10.3390/genes13071146
APA StyleGedikbasi, A., Adas, G., Isiksacan, N., Kart Yasar, K., Canbolat Unlu, E., Yilmaz, R., Hergunsel, G. O., & Cukurova, Z. (2022). The Effect of Host miRNAs on Prognosis in COVID-19: miRNA-155 May Promote Severity via Targeting Suppressor of Cytokine Signaling 1 (SOCS1) Gene. Genes, 13(7), 1146. https://doi.org/10.3390/genes13071146