
Polymorphisms in COMT and OPRM1 Collectively Contribute to Chronic Shoulder Pain and Disability in South African Breast Cancer Survivors’


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design, Participants, and Settings
2.2. Instruments
2.2.1. Shoulder Pain and Disability Index (SPADI)
2.2.2. SNP Selection and Genotyping
2.3. Statistical Analysis
3. Results
3.1. Participants’ Characteristics
3.2. COMT SNP Genotype Effects on Demographical and Clinical Characteristics
3.3. COMT SNP Frequencies
3.4. Genotype and Allele Frequency Distribution of COMT
3.5. Inferred COMT Haplotypes
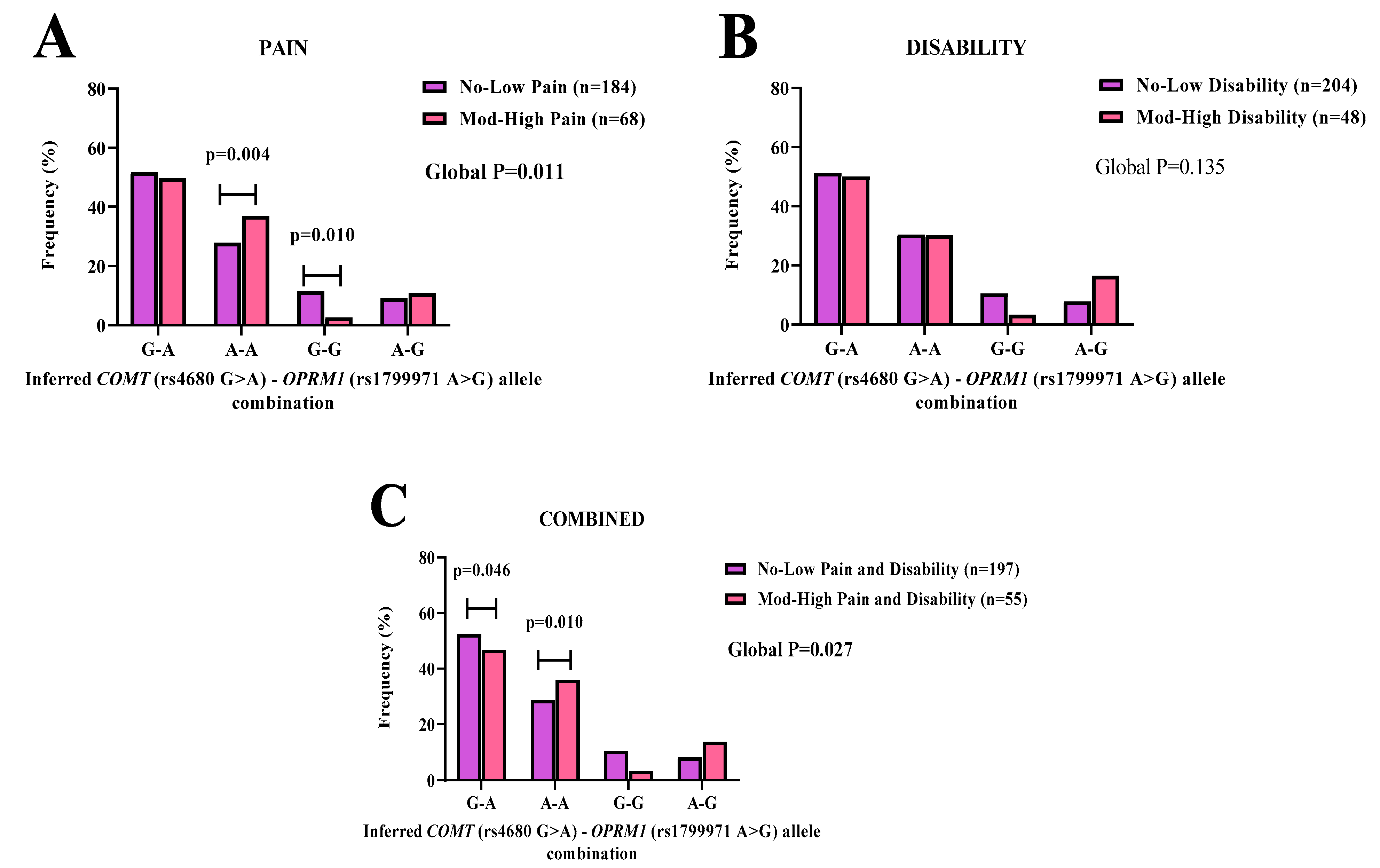
3.6. COMT-OPRM1 Allelic Combinations
3.7. Bioinformatic Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hidding, J.T.; Beurskens, C.H.; van der Wees, P.J.; van Laarhoven, H.W.; Nijhuis-van der Sanden, M.W. Treatment related impairments in arm and shoulder in patients with breast cancer: A systematic review. PLoS ONE 2014, 9, e96748. [Google Scholar] [CrossRef]
- Shamley, D.; Srinaganathan, R.; Oskrochi, R.; Lascurain-Aguirrebena, I.; Sugden, E. Three-dimensional scapulothoracic motion following treatment for breast cancer. Breast Cancer Res. Treat. 2009, 118, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Guyatt, G.H.; Kennedy, S.A.; Romerosa, B.; Kwon, H.Y.; Kaushal, A.; Chang, Y.; Craigie, S.; de Almeida, C.P.B.; Couban, R.J.; et al. Predictors of persistent pain after breast cancer surgery: A systematic review and meta-analysis of observational studies. Can. Med Assoc. J. 2016, 188, E352–E361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salz, T.; Lavery, J.A.; Lipitz-Snyderman, A.N.; Boudreau, D.M.; Moryl, N.; Gillespie, E.F.; Korenstein, D. Trends in Opioid Use Among Older Survivors of Colorectal, Lung, and Breast Cancers. J. Clin. Oncol. 2019, 37, 1001–1011. [Google Scholar] [CrossRef] [PubMed]
- Knisely, M.R.; Conley, Y.P.; Smoot, B.; Paul, S.M.; Levine, J.D.; Miaskowski, C. Associations Between Catecholaminergic and Serotonergic Genes and Persistent Arm Pain Severity Following Breast Cancer Surgery. J. Pain 2019, 20, 1100–1111. [Google Scholar] [CrossRef] [PubMed]
- Andersen, K.G.; Duriaud, H.M.; Jensen, H.E.; Kroman, N.; Kehlet, H. Predictive factors for the development of persistent pain after breast cancer surgery. Pain 2015, 156, 2413–2422. [Google Scholar] [CrossRef]
- Dimova, V.; Lotsch, J.; Huhne, K.; Winterpacht, A.; Heesen, M.; Parthum, A.; Weber, P.G.; Carbon, R.; Griessinger, N.; Sittl, R.; et al. Association of genetic and psychological factors with persistent pain after cosmetic thoracic surgery. J. Pain Res. 2015, 8, 829–844. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.Y.; Wang, P.; Zhao, D.F.; Gonzalez, F.J.; Fan, Y.F.; Xia, Y.L.; Ge, G.B.; Yang, L. Analytical methodologies for sensing catechol-O-methyltransferase activity and their applications. J. Pharm. Anal. 2021, 11, 15–27. [Google Scholar] [CrossRef]
- Bates, G.W.; Edman, C.D.; Porter, J.C.; MacDonald, P.C. Catechol-O-methyltransferase activity in erythrocytes of pregnant women. Am. J. Obstet. Gynecol. 1978, 131, 555–557. [Google Scholar] [CrossRef]
- Paravati, S.; Rosani, A.; Warrington, S.J. Physiology, Catecholamines. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Garcha, A.S.; Cohen, D.L. Catecholamine excess: Pseudopheochromocytoma and beyond. Adv. Chronic Kidney Dis. 2015, 22, 218–223. [Google Scholar] [CrossRef]
- Diatchenko, L.; Slade, G.D.; Nackley, A.G.; Bhalang, K.; Sigurdsson, A.; Belfer, I.; Goldman, D.; Xu, K.; Shabalina, S.A.; Shagin, D.; et al. Genetic basis for individual variations in pain perception and the development of a chronic pain condition. Hum. Mol. Genet. 2005, 14, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Schlereth, T.; Birklein, F. The sympathetic nervous system and pain. Neuromolecular. Med. 2008, 10, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Bjorland, S.; Moen, A.; Schistad, E.; Gjerstad, J.; Roe, C. Genes associated with persistent lumbar radicular pain; a systematic review. BMC Musculoskelet. Disord. 2016, 17, 500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nackley, A.G.; Shabalina, S.A.; Tchivileva, I.E.; Satterfield, K.; Korchynskyi, O.; Makarov, S.S.; Maixner, W.; Diatchenko, L. Human catechol-O-methyltransferase haplotypes modulate protein expression by altering mRNA secondary structure. Science 2006, 314, 1930–1933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machius, M.; Declerck, N.; Huber, R.; Wiegand, G. Kinetic stabilization of Bacillus licheniformis α-amylase through introduction of hydrophobic residues at the surface. J. Biol. Chem. 2003, 278, 11546–11553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmack, K.; Rossler, H.; Sekutowicz, M.; Brandl, E.J.; Muller, D.J.; Petrovic, P.; Sterzer, P. Linking unfounded beliefs to genetic dopamine availability. Front. Hum. Neurosci. 2015, 9, 521. [Google Scholar] [CrossRef] [Green Version]
- Khalil, H.; Sereika, S.M.; Dai, F.; Alexander, S.; Conley, Y.; Gruen, G.; Meng, L.; Siska, P.; Tarkin, I.; Henker, R. OPRM1 and COMT Gene-Gene Interaction Is Associated With Postoperative Pain and Opioid Consumption After Orthopedic Trauma. Biol. Res. Nurs. 2017, 19, 170–179. [Google Scholar] [CrossRef] [Green Version]
- Sadhasivam, S.; Chidambaran, V.; Olbrecht, V.A.; Esslinger, H.R.; Zhang, K.; Zhang, X.; Martin, L.J. Genetics of pain perception, COMT and postoperative pain management in children. Pharmacogenomics 2014, 15, 277–284. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Belfer, I.; Nouraie, M.; Zeng, Q.; Goel, R.; Chu, Y.; Krasiy, I.; Krishnamurti, L. Association of genetic variation in COMT gene with pain related to sickle cell disease in patients from the walk-PHaSST study. J. Pain Res. 2018, 11, 537–543. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, N.; Kidd, K.K.; Pakstis, A.J.; Speed, W.C.; Li, H.; Tarnok, Z.; Barta, C.; Kajuna, S.L.; Kidd, J.R. The complex global pattern of genetic variation and linkage disequilibrium at catechol-O-methyltransferase. Mol. Psychiatry 2010, 15, 216–225. [Google Scholar] [CrossRef]
- De Wit, E.; Delport, W.; Rugamika, C.E.; Meintjes, A.; Moller, M.; van Helden, P.D.; Seoighe, C.; Hoal, E.G. Genome-wide analysis of the structure of the South African Coloured Population in the Western Cape. Hum. Genet. 2010, 128, 145–153. [Google Scholar] [CrossRef]
- Ikediobi, O.; Aouizerat, B.; Xiao, Y.; Gandhi, M.; Gebhardt, S.; Warnich, L. Analysis of pharmacogenetic traits in two distinct South African populations. Hum. Genom. 2011, 5, 265–282. [Google Scholar] [CrossRef] [Green Version]
- Kowarik, M.C.; Einhauser, J.; Jochim, B.; Buttner, A.; Tolle, T.R.; Riemenschneider, M.; Platzer, S.; Berthele, A. Impact of the COMT Val(108/158)Met polymorphism on the mu-opioid receptor system in the human brain: Mu-opioid receptor, met-enkephalin and β-endorphin expression. Neurosci. Lett. 2012, 506, 214–219. [Google Scholar] [CrossRef]
- Yao, P.; Ding, Y.Y.; Wang, Z.B.; Ma, J.M.; Hong, T.; Pan, S.N. Effect of gene polymorphism of COMT and OPRM1 on the preoperative pain sensitivity in patients with cancer. Int. J. Clin. Exp. Med. 2015, 8, 10036–10039. [Google Scholar]
- Corder, G.; Castro, D.C.; Bruchas, M.R.; Scherrer, G. Endogenous and Exogenous Opioids in Pain. Annu. Rev. Neurosci. 2018, 41, 453–473. [Google Scholar] [CrossRef]
- Ren, Z.Y.; Xu, X.Q.; Bao, Y.P.; He, J.; Shi, L.; Deng, J.H.; Gao, X.J.; Tang, H.L.; Wang, Y.M.; Lu, L. The impact of genetic variation on sensitivity to opioid analgesics in patients with postoperative pain: A systematic review and meta-analysis. Pain Physician 2015, 18, 131–152. [Google Scholar]
- Hwang, I.C.; Park, J.Y.; Myung, S.K.; Ahn, H.Y.; Fukuda, K.; Liao, Q. OPRM1 A118G gene variant and postoperative opioid requirement: A systematic review and meta-analysis. Anesthesiology 2014, 121, 825–834. [Google Scholar] [CrossRef] [Green Version]
- Saiz-Rodriguez, M.; Ochoa, D.; Herrador, C.; Belmonte, C.; Roman, M.; Alday, E.; Koller, D.; Zubiaur, P.; Mejia, G.; Hernandez-Martinez, M.; et al. Polymorphisms associated with fentanyl pharmacokinetics, pharmacodynamics and adverse effects. Basic Clin. Pharmacol. Toxicol. 2019, 124, 321–329. [Google Scholar] [CrossRef] [Green Version]
- Firfirey, F.; September, A.V.; Shamley, D. ABCB1 and OPRM1 single-nucleotide polymorphisms collectively modulate chronic shoulder pain and dysfunction in South African breast cancer survivors. Pharmacogenomics 2022, 23, 513–530. [Google Scholar] [CrossRef]
- Reyes-Gibby, C.C.; Shete, S.; Rakvag, T.; Bhat, S.V.; Skorpen, F.; Bruera, E.; Kaasa, S.; Klepstad, P. Exploring joint effects of genes and the clinical efficacy of morphine for cancer pain: OPRM1 and COMT gene. Pain 2007, 130, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Little, J.; Higgins, J.P.; Ioannidis, J.P.; Moher, D.; Gagnon, F.; Von Elm, E.; Khoury, M.J.; Cohen, B.; Davey-Smith, G.; Grimshaw, J. STrengthening the REporting of Genetic Association Studies (STREGA)—an extension of the STROBE statement. Genet. Epidemiol. Off. Publ. Int. Genet. Epidemiol. Soc. 2009, 33, 581–598. [Google Scholar]
- Tengrup, I.; Tennvall-Nittby, L.; Christiansson, I.; Laurin, M. Arm morbidity after breast-conserving therapy for breast cancer. Acta Oncol. 2000, 39, 393–397. [Google Scholar] [CrossRef] [Green Version]
- MacDermid, J.C.; Solomon, P.; Prkachin, K. The Shoulder Pain and Disability Index demonstrates factor, construct and longitudinal validity. BMC Musculoskelet. Disord. 2006, 7, 12. [Google Scholar] [CrossRef]
- Gauderman, W.; Morrison, J. QUANTO 1.1: A Computer Program for Power and Sample Size Calculations for Genetic-epidemiology Studies, Version 1.2.4. 2006. Available online: http://hydra.usc.edu/gxe (accessed on 22 November 2022).
- Dell. Inc. Dell Statistica (Data Analysis Software System) [Computer Program]. Version 13. 2016. Available online: www.statsoft.com (accessed on 30 April 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; ISBN 3-900051-07-0. [Google Scholar]
- Warnes, G.; Gorjanc, wcfG.; Leisch, F.; Man, M. _genetics: Population Genetics_. R package version 1.3.8.1.3. 2021. Available online: https://CRAN.R-project.org/package=genetics (accessed on 22 November 2022).
- González, J.; Armengol, L.; Guinó, E.; Solé, X.; Moreno, V. SNPassoc: SNPs-based whole genome association studies. R Package Version 2014, 1, 2–9. [Google Scholar]
- Sinnwell, J.P.; Schaid, D. Statistical Methods for Haplotypes When Linkage Phase Is Ambiguous; Mayo Clinic Division of Health Sciences Research: Rochester, MN, USA, 2011. [Google Scholar]
- Doong, S.H.; Dhruva, A.; Dunn, L.B.; West, C.; Paul, S.M.; Cooper, B.A.; Elboim, C.; Abrams, G.; Merriman, J.D.; Langford, D.J.; et al. Associations between cytokine genes and a symptom cluster of pain, fatigue, sleep disturbance, and depression in patients prior to breast cancer surgery. Biol. Res. Nurs. 2015, 17, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Henker, R.A.; Lewis, A.; Dai, F.; Lariviere, W.R.; Meng, L.; Gruen, G.S.; Sereika, S.M.; Pape, H.; Tarkin, I.S.; Gowda, I. The associations between OPRM 1 and COMT genotypes and postoperative pain, opioid use, and opioid-induced sedation. Biol. Res. Nurs. 2013, 15, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Jauand, M.; Sitges, C.; Rodriguez, V.; Picornell, A.; Ramon, M.; Buskila, D.; Montoya, P. Pain sensitivity in fibromyalgia is associated with catechol-O-methyltransferase (COMT) gene. Eur. J. Pain 2013, 17, 16–27. [Google Scholar] [CrossRef]
- Baumbauer, K.M.; Ramesh, D.; Perry, M.; Carney, K.B.; Julian, T.; Glidden, N.; Dorsey, S.G.; Starkweather, A.R.; Young, E.E. Contribution of COMT and BDNF Genotype and Expression to the Risk of Transition From Acute to Chronic Low Back Pain. Clin. J. Pain 2020, 36, 430–439. [Google Scholar] [CrossRef]
- Shamley, D. A Cross-Disciplinary Look at Shoulder Pain and Dysfunction after Treatment for Breast Cancer. Int. J. Cancer Clin. Res. 2015, 2. [Google Scholar] [CrossRef]
- Chimusa, E.R.; Meintjies, A.; Tchanga, M.; Mulder, N.; Seoighe, C.; Soodyall, H.; Ramesar, R. A genomic portrait of haplotype diversity and signatures of selection in indigenous southern African populations. PLoS Genet. 2015, 11, e1005052. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Lipska, B.K.; Halim, N.; Ma, Q.D.; Matsumoto, M.; Melhem, S.; Kolachana, B.S.; Hyde, T.M.; Herman, M.M.; Apud, J.; et al. Functional analysis of genetic variation in catechol-O-methyltransferase (COMT): Effects on mRNA, protein, and enzyme activity in postmortem human brain. Am. J. Hum. Genet. 2004, 75, 807–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vizan, P.; Di Croce, L.; Aranda, S. Functional and Pathological Roles of AHCY. Front. Cell Dev. Biol. 2021, 9, 654344. [Google Scholar] [CrossRef]
- Mori, S.; Hatori, N.; Kawaguchi, N.; Hamada, Y.; Shih, T.C.; Wu, C.Y.; Lam, K.S.; Matsuura, N.; Yamamoto, H.; Takada, Y.K.; et al. The integrin-binding defective FGF2 mutants potently suppress FGF2 signalling and angiogenesis. Biosci. Rep. 2017, 37, BSR20170173. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Hand, W.; Alexov, E. Opioid Addiction and Opioid Receptor Dimerization: Structural Modeling of the OPRD1 and OPRM1 Heterodimer and Its Signaling Pathways. Int. J. Mol. Sci. 2021, 22, 10290. [Google Scholar] [CrossRef]
- Gonzalez-Nunez, V.; González, A.J.; Barreto-Valer, K.; Rodríguez, R.E. In Vivo Regulation of the μ Opioid Receptor: Role of the Endogenous Opioid Agents. Mol. Med. 2013, 19, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Wang, R.T.; Ahn, S.; Park, C.C.; Smith, D.J. A genome-wide map of human genetic interactions inferred from radiation hybrid genotypes. Genome Res. 2010, 20, 1122–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobbin, K.K.; Beer, D.G.; Meyerson, M.; Yeatman, T.J.; Gerald, W.L.; Jacobson, J.W.; Conley, B.; Buetow, K.H.; Heiskanen, M.; Simon, R.M.; et al. Interlaboratory comparability study of cancer gene expression analysis using oligonucleotide microarrays. Clin. Cancer Res. 2005, 11, 565–572. [Google Scholar] [CrossRef]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.B.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene Set Knowledge Discovery with Enrichr. Curr. Protoc. 2021, 1, e90. [Google Scholar] [CrossRef]
- Li, L.; Jia, J.; Liu, X.; Yang, S.; Ye, S.; Yang, W.; Zhang, Y. MicroRNA-16-5p Controls Development of Osteoarthritis by Targeting SMAD3 in Chondrocytes. Curr. Pharm. Des. 2015, 21, 5160–5167. [Google Scholar] [CrossRef]
- Patel, J.N.; Hamadeh, I.S. Pharmacogenomics-guided opioid management. BMJ Support. Palliat. Care 2020, 10, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Matic, M.; Jongen, J.L.; Elens, L.; de Wildt, S.N.; Tibboel, D.; Sillevis Smitt, P.A.; van Schaik, R.H. Advanced cancer pain: The search for genetic factors correlated with interindividual variability in opioid requirement. Pharmacogenomics 2017, 18, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Gilbert-Diamond, D.; Moore, J.H. Analysis of gene-gene interactions. Curr. Protoc. Hum. Genet. 2011, 70, 14. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pain | Disability | Pain and Disability | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Polymorphisms | No-Low | Mod-High | No-Low | Mod-High | No-Low | Mod-High | ||||||
| (n = 184) | (n = 68) | AIC | (n = 204) | (n = 48) | (n = 197) | (n = 55) | AIC | |||||
| COMT | ||||||||||||
| rs6269 A > G | ||||||||||||
| G/G | 38.9 (68) | 35.4 (23) | 38.5 (75) | 35.6 (16) | 37.8 (71) | 38.5 (20) | ||||||
| A/G | 45.1 (79) | 46.2 (30) | 45.1 (88) | 46.7 (21) | 46.3 (87) | 42.3 (22) | ||||||
| A/A | 16.0 (28) | 18.5 (12) | 16.4 (32) | 17.8 (8) | 16.0 (30) | 19.2 (10) | ||||||
| G allele | 61.4 (215) | 58.5 (76) | 61.0 (238) | 58.9 (53) | 60.9 (229) | 59.6 (62) | ||||||
| p value 1 | 0.848 | 0.937 | 0.787 | |||||||||
| G Allele p value 2 | 0.600 | 0.721 | 0.821 | |||||||||
| HWE | 0.532 | 0.617 | 0.457 | 1.000 | 0.651 | 0.406 | ||||||
| rs4633 C > T | ||||||||||||
| T/T | 34.3 (60) | 29.2 (19) | 34.9 (68) | 24.4 (11) | 35.1 (66) | 25 (13) | ||||||
| C/T | 46.9 (82) | 47.7 (31) | 44.6 (87) | 57.8 (26) | 45.7 (86) | 51.9 (27) | ||||||
| C/C | 18.9 (33) | 23.1 (15) | 20.5 (40) | 17.8 (8) | 19.1 (36) | 23.1 (12) | ||||||
| T allele | 57.7 (202) | 53.1 (69) | 57.2 (223) | 53.3 (48) | 58.0 (218) | 51.0 (53) | ||||||
| p value 1 | 0.557 | 0.178 | 0.261 | |||||||||
| T Allele p value 2 | 0.407 | 0.556 | 0.220 | |||||||||
| HWE | 0.546 | 0.628 | 0.154 | 0.389 | 0.307 | 1.000 | ||||||
| rs4818 C > G | ||||||||||||
| C/C | 52.0 (89) | 52.3 (34) | 51.6 (99) | 54.5 (24) | 50.8 (94) | 56.9 (29) | ||||||
| C/G | 43.3 (74) | 43.1 (28) | 43.8 (84) | 40.9 (18) | 44.3 (82) | 39.2 (20) | ||||||
| G/G | 4.7 (8) | 4.6 (3) | 4.7 (9) | 4.5 (2) | 4.9 (9) | 3.9 (2) | ||||||
| G allele | 26.3 (90) | 26.2 (34) | 26.6 (102) | 25.0 (22) | 27.0 (100) | 23.5 (24) | ||||||
| p value 1 | 0.480 | 0.880 | 0.618 | |||||||||
| G Allele p value 2 | 1.000 | 0.893 | 0.527 | |||||||||
| HWE | 0.247 | 0.526 | 0.201 | 0.702 | 0.199 | 0.707 | ||||||
| rs4680 G > A | ||||||||||||
| G/G | 41.0 (71) | 27.7 (18) | 0.015 a | 268.3 | 39.9 (77) | 26.7 (12) | 40.9 (76) | 25.0 (13) | 0.009 a | 240.0 | ||
| A/G | 46.2 (80) | 50.8 (33) | 0.382 b | 273.4 | 45.6 (88) | 55.6 (25) | 46.2 (86) | 51.9 (27) | 0.342 b | 245.9 | ||
| A/A | 12.7 (22) | 21.5 (14) | 0.050 c | 270.4 | 14.5 (28) | 17.8 (8) | 12.9 (24) | 23.1 (12) | 0.041 c | 242.6 | ||
| A allele | 35.8 (124) | 46.9 (61) | 37.3 (144) | 45.6 (41) | 36.0 (134) | 49.0 (51) | ||||||
| p value 1 | 0.024 | 268.7 | 0.113 | 0.015 | 240.3 | |||||||
| A Allele p value 2 | 0.035 | 0.152 | 0.017 | |||||||||
| HWE | 0.874 | 1.000 | 0.550 | 0.564 | 0.877 | 1.000 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Firfirey, F.; Shamley, D.; September, A.V. Polymorphisms in COMT and OPRM1 Collectively Contribute to Chronic Shoulder Pain and Disability in South African Breast Cancer Survivors’. Genes 2023, 14, 9. https://doi.org/10.3390/genes14010009
Firfirey F, Shamley D, September AV. Polymorphisms in COMT and OPRM1 Collectively Contribute to Chronic Shoulder Pain and Disability in South African Breast Cancer Survivors’. Genes. 2023; 14(1):9. https://doi.org/10.3390/genes14010009
Chicago/Turabian StyleFirfirey, Firzana, Delva Shamley, and Alison V. September. 2023. "Polymorphisms in COMT and OPRM1 Collectively Contribute to Chronic Shoulder Pain and Disability in South African Breast Cancer Survivors’" Genes 14, no. 1: 9. https://doi.org/10.3390/genes14010009
APA StyleFirfirey, F., Shamley, D., & September, A. V. (2023). Polymorphisms in COMT and OPRM1 Collectively Contribute to Chronic Shoulder Pain and Disability in South African Breast Cancer Survivors’. Genes, 14(1), 9. https://doi.org/10.3390/genes14010009