
A Bird’s-Eye View of Endangered Species Conservation: Avian Genomics and Stem Cell Approaches for Green Peafowl (Pavo muticus)


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Peafowl and Green Peafowl
3. Avian and Peafowl Development
4. Avian and Peafowl Genomics and Genetics
Avian and Peafowl Omics Resources
5. Avian and Peafowl Stem Cell Biology
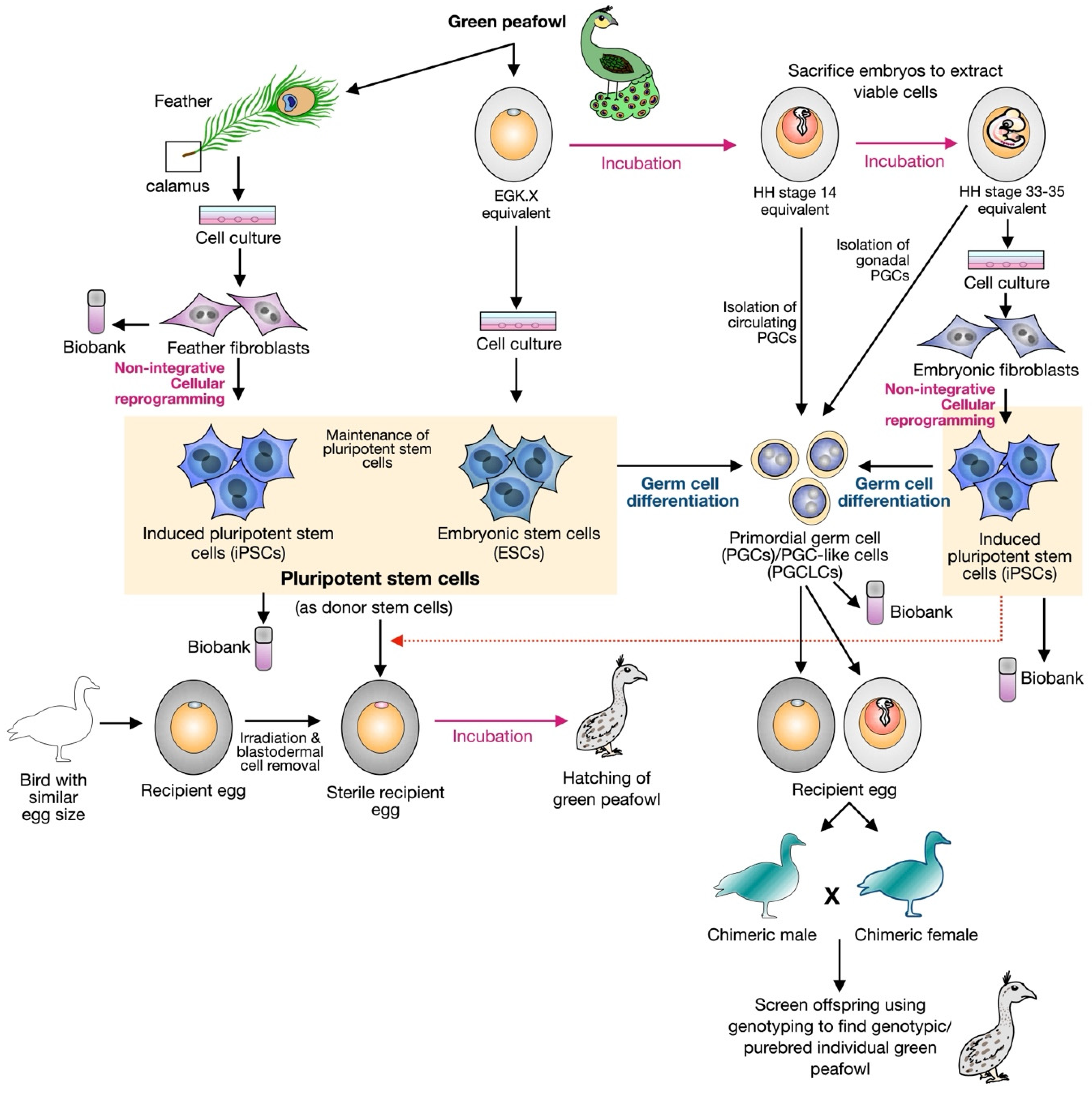
Approaches for Applying Stem Cell Research in Peafowl Conservation
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IUCN. 2020. Available online: www.iucn.org/about (accessed on 19 July 2023).
- Singh, B.; Mal, G.; Gautam, S.K.; Mukesh, M. Biotechnology for Wildlife. In Advances in Animal Biotechnology; Springer International Publishing: Cham, Switzerland, 2019; pp. 501–513. [Google Scholar]
- Comizzoli, P. Biotechnologies for wildlife fertility preservation. Anim. Front. 2015, 5, 73–78. [Google Scholar] [CrossRef]
- Barna, J.; Végi, B.; Liptói, K.; Patakiné Várkonyi, E. Chapter 13—Reproductive technologies in avian species. In Reproductive Technologies in Animals; Presicce, G.A., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 193–228. [Google Scholar]
- Comizzoli, P.; Holt, W.V. Breakthroughs and new horizons in reproductive biology of rare and endangered animal species. Biol. Reprod. 2019, 101, 514–525. [Google Scholar] [CrossRef]
- Bolton, R.L.; Mooney, A.; Pettit, M.T.; Bolton, A.E.; Morgan, L.; Drake, G.J.; Appeltant, R.; Walker, S.L.; Gillis, J.D.; Hvilsom, C. Resurrecting biodiversity: Advanced assisted reproductive technologies and biobanking. Reprod. Fertil. 2022, 3, R121–R146. [Google Scholar] [CrossRef]
- Mastromonaco, G.F.; Songsasen, N. Chapter 7—Reproductive technologies for the conservation of wildlife and endangered species. In Reproductive Technologies in Animals; Presicce, G.A., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 99–117. [Google Scholar]
- Sun, Y.; Li, Y.; Zong, Y.; Mehaisen, G.M.K.; Chen, J. Poultry genetic heritage cryopreservation and reconstruction: Advancement and future challenges. J. Anim. Sci. Biotechnol. 2022, 13, 115. [Google Scholar] [CrossRef] [PubMed]
- Tajima, A. Conservation of Avian Genetic Resources. J. Poult. Sci. 2013, 50, 1–8. [Google Scholar] [CrossRef]
- Jaiswal, S.K.; Gupta, A.; Saxena, R.; Prasoodanan, V.P.K.; Sharma, A.K.; Mittal, P.; Roy, A.; Shafer, A.B.A.; Vijay, N.; Sharma, V.K. Genome Sequence of Peacock Reveals the Peculiar Case of a Glittering Bird. Front. Genet. 2018, 9, 392. [Google Scholar] [CrossRef]
- Zhou, T.C.; Sha, T.; Irwin, D.M.; Zhang, Y.P. Complete mitochondrial genome of the Indian peafowl (Pavo cristatus), with phylogenetic analysis in phasianidae. Mitochondrial DNA 2015, 26, 912–913. [Google Scholar] [CrossRef]
- Zhang, X.; Lin, C.; Li, H.; Liu, S.; Wang, Q.; Yang, S.; Shi, M.; Sahu, S.K.; Zhu, Y.; Wang, J.; et al. Chromosome-Level Genome Assembly of the Green Peafowl (Pavo muticus). Genome Biol. Evol. 2022, 14, evac015. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chen, H.; Ouyang, J.; Huang, M.; Zhang, H.; Zheng, S.; Xi, S.; Tang, H.; Gao, Y.; Xiong, Y.; et al. A high-quality assembly reveals genomic characteristics, phylogenetic status, and causal genes for leucism plumage of Indian peafowl. Gigascience 2022, 11, giac018. [Google Scholar] [CrossRef] [PubMed]
- Hohenlohe, P.A.; Funk, W.C.; Rajora, O.P. Population genomics for wildlife conservation and management. Mol. Ecol. 2021, 30, 62–82. [Google Scholar] [CrossRef]
- Du, H.Y.; Zhang, X.Y.; Dinh, T.D.; Ma, Y.; Zong, C.; Li, G.L.; Dahmer, T.D.; Xu, Y.C. Identification of hybrid green peafowl using mitochondrial and nuclear markers. Conserv. Genet. Resour. 2020, 12, 669–683. [Google Scholar] [CrossRef]
- Dong, F.; Kuo, H.C.; Chen, G.L.; Wu, F.; Shan, P.F.; Wang, J.; Chen, D.; Lei, F.M.; Hung, C.M.; Liu, Y.; et al. Population genomic, climatic and anthropogenic evidence suggest the role of human forces in endangerment of green peafowl (Pavo muticus). Proc. R. Soc. B Biol. Sci. 2021, 288, 20210073. [Google Scholar] [CrossRef] [PubMed]
- Katayama, M.; Fukuda, T.; Kaneko, T.; Nakagawa, Y.; Tajima, A.; Naito, M.; Ohmaki, H.; Endo, D.; Asano, M.; Nagamine, T.; et al. Induced pluripotent stem cells of endangered avian species. Commun. Biol. 2022, 5, 1049. [Google Scholar] [CrossRef]
- Han, J.Y.; Lee, H.C.; Park, T.S. Germline-competent stem cell in avian species and its application. Asian J. Androl. 2015, 17, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Rosselló, R.A.; Chen, C.C.; Dai, R.; Howard, J.T.; Hochgeschwender, U.; Jarvis, E.D. Mammalian genes induce partially reprogrammed pluripotent stem cells in non-mammalian vertebrate and invertebrate species. eLife 2013, 2, e00036. [Google Scholar] [CrossRef] [PubMed]
- Fuet, A.; Pain, B. Chicken induced pluripotent stem cells: Establishment and characterization. Methods Mol. Biol. 2017, 1650, 211–228. [Google Scholar]
- Lu, Y.; West, F.D.; Jordan, B.J.; Beckstead, R.B.; Jordan, E.T.; Stice, S.L. Generation of Avian Induced Pluripotent Stem Cells. Methods Mol. Biol. 2015, 1330, 89–99. [Google Scholar]
- Lu, Y.; West, F.D.; Jordan, B.J.; Mumaw, J.L.; Jordan, E.T.; Gallegos-Cardenas, A.; Beckstead, R.B.; Stice, S.L. Avian-induced pluripotent stem cells derived using human reprogramming factors. Stem Cells Dev. 2012, 21, 394–403. [Google Scholar] [CrossRef]
- Lu, Y.; West, F.D.; Jordan, B.J.; Jordan, E.T.; West, R.C.; Yu, P.; He, Y.; Barrios, M.A.; Zhu, Z.; Petitte, J.N.; et al. Induced pluripotency in chicken embryonic fibroblast results in a germ cell fate. Stem Cells Dev. 2014, 23, 1755–1764. [Google Scholar] [CrossRef]
- Yu, P.; Lu, Y.; Jordan, B.J.; Liu, Y.; Yang, J.Y.; Hutcheson, J.M.; Ethridge, C.L.; Mumaw, J.L.; Kinder, H.A.; Beckstead, R.B.; et al. Nonviral minicircle generation of induced pluripotent stem cells compatible with production of chimeric chickens. Cell. Reprogram. 2014, 16, 366–378. [Google Scholar] [CrossRef]
- Mukhtar, N.; Ahmad, T.; Zee-Waqar, M. Exotic avian species. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2022. [Google Scholar]
- Rajpoot, A.; Kumar, V.P.; Arunachalam, K.; Rasaily, S.S. National bird, Indian peafowl (Pavo cristatus): Using DNA technology for species identification from degraded sample from Uttarakhand, India. Forensic Sci. Int. Anim. Environ. 2021, 1, 100004. [Google Scholar] [CrossRef]
- Beebe, W. A Monograph of the Pheasants; H.F. & G. Witherby: London, UK, 1922; Volume IV, p. 242. [Google Scholar]
- Coles, B.H. 13—Galliformes. In Handbook of Avian Medicine, 2nd ed.; Tully, T.N., Dorrestein, G.M., Jones, A.K., Cooper, J.E., Eds.; W.B. Saunders: Edinburgh, Scotland, 2009; pp. 309–334. [Google Scholar]
- Deignan, H.G. Museum of Natural History Checklist of the Birds of Thailand; Smithsonian Institution: Washington, DC, USA, 1963. [Google Scholar]
- McGowan, P.J.K.; Duckworth, J.W.; Xianji, W.; Van Balen, B.; Xiaojun, Y.; Mohd, U.; Khan, K.M.; Yatim, S.H.; Thanga, L.; Setiawan, I.; et al. A review of the status of the Green Peafowl Pavo muticus and recommendations for future action. Bird Conserv. Int. 1998, 8, 331–348. [Google Scholar] [CrossRef]
- Sukumal, N.; Dowell, S.D.; Savini, T. Micro-habitat selection and population recovery of the Endangered Green Peafowl Pavo muticus in western Thailand: Implications for conservation guidance. Bird Conserv. Int. 2017, 27, 414–430. [Google Scholar] [CrossRef]
- Urban, M.C. Climate change. Accelerating extinction risk from climate change. Science 2015, 348, 571–573. [Google Scholar] [CrossRef] [PubMed]
- Jose, V.S.; Nameer, P.O. The expanding distribution of the Indian Peafowl (Pavo cristatus) as an indicator of changing climate in Kerala, southern India: A modelling study using MaxEnt. Ecol. Indic. 2020, 110, 105930. [Google Scholar] [CrossRef]
- Wang, B.; Ye, W.; Xu, Y.; Zhong, X.; Zhang, J.; Yang, N.; Yang, B.; Zhou, C. Climate change affects Galliformes taxonomic, phylogenetic and functional diversity indexes, shifting conservation priority areas in China. Divers. Distrib. 2023, 29, 409–422. [Google Scholar] [CrossRef]
- Li, B.; Liang, C.; Song, P.; Liu, D.; Qin, W.; Jiang, F.; Gu, H.; Gao, H.; Zhang, T. Threatened birds face new distribution under future climate change on the Qinghai-Tibet Plateau (QTP). Ecol. Indic. 2023, 150, 110217. [Google Scholar] [CrossRef]
- Liu, Z.; Tian, S.; Lu, S.; Zhu, Z.; Peng, Y.; Li, X.; An, L.; Li, J.; Xu, J.; Wang, Y. Climate and land-use changes threaten the effectiveness of protected areas for protecting Galliformes in Southeast Asia. Front. Ecol. Evol 2023, 11, 1216769. [Google Scholar] [CrossRef]
- Cao, A.; Shi, X. The Effects of Climate Change on Habitat Connectivity: A Case Study of the Brown-Eared Pheasant in China. Land 2022, 11, 806. [Google Scholar] [CrossRef]
- Asgharzadeh, M.; Alesheikh, A.A.; Yousefi, M. Disentangling the impacts of climate and land cover changes on habitat suitability of common pheasant Phasianus colchicus along elevational gradients in Iran. Environ. Sci. Pollut. Res. 2023, 30, 60958–60966. [Google Scholar] [CrossRef]
- Pearce-Higgins, J.W. Climate Change and the UK’s Birds; British Trust for Ornithology Report: Thetford, UK, 2021; p. 50. [Google Scholar]
- Root, T.L.; Price, J.T.; Hall, K.R.; Schneider, S.H.; Rosenzweig, C.; Pounds, J.A. Fingerprints of global warming on wild animals and plants. Nature 2003, 421, 57–60. [Google Scholar] [CrossRef]
- Kong, D.; Wu, F.; Shan, P.; Gao, J.; Yan, D.; Luo, W.; Yang, X. Status and distribution changes of the endangered Green Peafowl (Pavo muticus) in China over the past three decades (1990s–2017). Avian Res. 2018, 9, 18. [Google Scholar] [CrossRef]
- Rose, P. Evidence for Aviculture: Identifying Research Needs to Advance the Role of Ex Situ Bird Populations in Conservation Initiatives and Collection Planning. Birds 2021, 2, 77–95. [Google Scholar] [CrossRef]
- Green, R.E.; Braun, E.L.; Armstrong, J.; Earl, D.; Nguyen, N.; Hickey, G.; Vandewege, M.W.; St John, J.A.; Capella-Gutierrez, S.; Castoe, T.A.; et al. Three crocodilian genomes reveal ancestral patterns of evolution among archosaurs. Science 2014, 346, 1254449. [Google Scholar] [CrossRef] [PubMed]
- Yonezawa, T.; Segawa, T.; Mori, H.; Campos, P.F.; Hongoh, Y.; Endo, H.; Akiyoshi, A.; Kohno, N.; Nishida, S.; Wu, J.; et al. Phylogenomics and Morphology of Extinct Paleognaths Reveal the Origin and Evolution of the Ratites. Curr. Biol. 2017, 27, 68–77. [Google Scholar] [CrossRef]
- Nagai, H.; Mak, S.S.; Weng, W.; Nakaya, Y.; Ladher, R.; Sheng, G. Embryonic development of the emu, Dromaius novaehollandiae. Dev. Dyn. 2011, 240, 162–175. [Google Scholar] [CrossRef]
- Eyal-Giladi, H.; Kochav, S. From cleavage to primitive streak formation: A complementary normal table and a new look at the first stages of the development of the chick. I. General morphology. Dev. Biol. 1976, 49, 321–337. [Google Scholar] [CrossRef]
- Hamburger, V.; Hamilton, H.L. A series of normal stages in the development of the chick embryo. J. Morphol. 1951, 88, 49–92. [Google Scholar] [CrossRef] [PubMed]
- Hedges, S.B.; Dudley, J.; Kumar, S. TimeTree: A public knowledge-base of divergence times among organisms. Bioinformatics 2006, 22, 2971–2972. [Google Scholar] [CrossRef]
- Feng, S.; Stiller, J.; Deng, Y.; Armstrong, J.; Fang, Q.; Reeve, A.H.; Xie, D.; Chen, G.; Guo, C.; Faircloth, B.C.; et al. Dense sampling of bird diversity increases power of comparative genomics. Nature 2020, 587, 252–257. [Google Scholar] [CrossRef]
- Wu, L.; Jiao, X.; Zhang, D.; Cheng, Y.; Song, G.; Qu, Y.; Lei, F. Comparative Genomics and Evolution of Avian Specialized Traits. Curr. Genomics. 2021, 22, 496–511. [Google Scholar] [CrossRef] [PubMed]
- Prum, R.O.; Berv, J.S.; Dornburg, A.; Field, D.J.; Townsend, J.P.; Lemmon, E.M.; Lemmon, A.R. A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature 2015, 526, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Grzywacz, B.; Skórka, P. Genome size versus geographic range size in birds. PeerJ 2021, 9, e10868. [Google Scholar] [CrossRef] [PubMed]
- Organ, C.L.; Shedlock, A.M.; Meade, A.; Pagel, M.; Edwards, S.V. Origin of avian genome size and structure in non-avian dinosaurs. Nature 2007, 446, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Li, C.; Li, Q.; Li, B.; Larkin, D.M.; Lee, C.; Storz, J.F.; Antunes, A.; Greenwold, M.J.; Meredith, R.W.; et al. Comparative genomics reveals insights into avian genome evolution and adaptation. Science 2014, 346, 1311–1320. [Google Scholar] [CrossRef]
- Graves, J.A. Avian sex, sex chromosomes, and dosage compensation in the age of genomics. Chromosome Res. 2014, 22, 45–57. [Google Scholar] [CrossRef]
- Smith, C.A.; Major, A.T.; Estermann, M.A. The Curious Case of Avian Sex Determination. Trends Genet. 2021, 37, 496–497. [Google Scholar] [CrossRef]
- Kratochvíl, L.; Gamble, T.; Rovatsos, M. Sex chromosome evolution among amniotes: Is the origin of sex chromosomes non-random? Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2021, 376, 20200108. [Google Scholar] [CrossRef]
- Deviatiiarov, R.; Nagai, H.; Ismagulov, G.; Stupina, A.; Wada, K.; Ide, S.; Toji, N.; Zhang, H.; Sukparangsi, W.; Intarapat, S.; et al. Dosage compensation of Z sex chromosome genes in avian fibroblast cells. Genome Biol. 2023, 24, 213. [Google Scholar] [CrossRef]
- Park, T.S.; Kang, K.S.; Han, J.Y. Current genomic editing approaches in avian transgenesis. Gen. Comp. Endocrinol. 2013, 190, 144–148. [Google Scholar] [CrossRef]
- Park, J.S.; Lee, K.Y.; Han, J.Y. Precise Genome Editing in Poultry and Its Application to Industries. Genes 2020, 11, 1182. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, D.H.; Lee, K. Current Approaches and Applications in Avian Genome Editing. Int. J. Mol. Sci. 2020, 21, 3937. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.M.; Woo, S.J.; Han, J.Y. Strategies for the Generation of Gene Modified Avian Models: Advancement in Avian Germline Transmission, Genome Editing, and Applications. Genes 2023, 14, 899. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.J.; Choi, J.W.; Kim, S.Y.; Park, K.J.; Kim, T.M.; Lee, Y.M.; Kim, H.; Lim, J.M.; Han, J.Y. Reproduction of wild birds via interspecies germ cell transplantation. Biol. Reprod. 2008, 79, 931–937. [Google Scholar] [CrossRef]
- Lee, J.; Kim, D.H.; Karolak, M.C.; Shin, S.; Lee, K. Generation of genome-edited chicken and duck lines by adenovirus-mediated in vivo genome editing. Proc. Natl. Acad. Sci. USA 2022, 119, e2214344119. [Google Scholar] [CrossRef]
- Garda, S.; Schwarz, J.M.; Schuelke, M.; Leser, U.; Seelow, D. Public data sources for regulatory genomic features. Med. Genet. 2021, 33, 167–177. [Google Scholar] [CrossRef]
- Pan, Z.; Wang, Y.; Wang, M.; Wang, Y.; Zhu, X.; Gu, S.; Zhong, C.; An, L.; Shan, M.; Damas, J.; et al. An atlas of regulatory elements in chicken: A resource for chicken genetics and genomics. Sci. Adv. 2023, 9, eade1204. [Google Scholar] [CrossRef]
- Chakraborty, A.; Mondal, S.; Mahajan, S.; Sharma, V.K. High-quality genome assemblies provide clues on the evolutionary advantage of blue peafowl over green peafowl. Heliyon 2023, 9, e18571. [Google Scholar] [CrossRef]
- Gee, G.F.; Bertschinger, H.; Donoghue, A.M.; Blanco, J.; Soley, J. Reproduction in nondomestic birds: Physiology, semen collection, artificial insemination and cryopreservation. Avian Poult. Biol. Rev. 2004, 15, 47–101. [Google Scholar] [CrossRef]
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic stem cell lines derived from human blastocysts. Science 1998, 282, 1145–1147. [Google Scholar] [CrossRef]
- Martin, G.R. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc. Natl. Acad. Sci. USA 1981, 78, 7634–7638. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef]
- Wang, L.; Su, Y.; Huang, C.; Yin, Y.; Chu, A.; Knupp, A.; Tang, Y. NANOG and LIN28 dramatically improve human cell reprogramming by modulating LIN41 and canonical WNT activities. Biol. Open 2019, 8, bio047225. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, M.; Yamaguchi, K.; Nakamura, T.; Shibukawa, R.; Kodanaka, I.; Ichisaka, T.; Kawamura, Y.; Mochizuki, H.; Goshima, N.; Yamanaka, S. Direct reprogramming of somatic cells is promoted by maternal transcription factor Glis1. Nature 2011, 474, 225–229. [Google Scholar] [CrossRef]
- Nakagawa, M.; Koyanagi, M.; Tanabe, K.; Takahashi, K.; Ichisaka, T.; Aoi, T.; Okita, K.; Mochiduki, Y.; Takizawa, N.; Yamanaka, S. Generation of induced pluripotent stem cells without Myc from mouse and human fibroblasts. Nat. Biotechnol. 2008, 26, 101–106. [Google Scholar] [CrossRef]
- Yu, J.; Vodyanik, M.A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S.; Nie, J.; Jonsdottir, G.A.; Ruotti, V.; Stewart, R.; et al. Induced pluripotent stem cell lines derived from human somatic cells. Science 2007, 318, 1917–1920. [Google Scholar] [CrossRef] [PubMed]
- Pain, B.; Clark, M.E.; Shen, M.; Nakazawa, H.; Sakurai, M.; Samarut, J.; Etches, R.J. Long-term in vitro culture and characterisation of avian embryonic stem cells with multiple morphogenetic potentialities. Development 1996, 122, 2339–2348. [Google Scholar] [CrossRef] [PubMed]
- Van De Lavoir, M.C.; Diamond, J.H.; Leighton, P.A.; Mather-Love, C.; Heyer, B.S.; Bradshaw, R.; Kerchner, A.; Hooi, L.T.; Gessaro, T.M.; Swanberg, S.E.; et al. Germline transmission of genetically modified primordial germ cells. Nature 2006, 441, 766–769. [Google Scholar] [CrossRef]
- Whyte, J.; Glover, J.D.; Woodcock, M.; Brzeszczynska, J.; Taylor, L.; Sherman, A.; Kaiser, P.; McGrew, M.J. FGF, Insulin, and SMAD Signaling Cooperate for Avian Primordial Germ Cell Self-Renewal. Stem Cell Rep. 2015, 5, 1171–1182. [Google Scholar] [CrossRef]
- Choi, H.W.; Kim, J.S.; Choi, S.; Ju Hong, Y.; Byun, S.J.; Seo, H.G.; Do, J.T. Mitochondrial remodeling in chicken induced pluripotent stem-like cells. Stem Cells Dev. 2016, 25, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Katayama, M.; Hirayama, T.; Tani, T.; Nishimori, K.; Onuma, M.; Fukuda, T. Chick derived induced pluripotent stem cells by the poly-cistronic transposon with enhanced transcriptional activity. J. Cell. Physiol. 2018, 233, 990–1004. [Google Scholar] [CrossRef] [PubMed]
- Jean, C.; Oliveira, N.M.M.; Intarapat, S.; Fuet, A.; Mazoyer, C.; De Almeida, I.; Trevers, K.; Boast, S.; Aubel, P.; Bertocchini, F.; et al. Transcriptome analysis of chicken ES, blastodermal and germ cells reveals that chick ES cells are equivalent to mouse ES cells rather than EpiSC. Stem Cell Res. 2015, 14, 54–67. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, J.; Glover, J.D.; Taylor, L.; Sang, H.M.; McGrew, M.J. Characterisation and Germline Transmission of Cultured Avian Primordial Germ Cells. PLoS ONE 2010, 5, e15518. [Google Scholar] [CrossRef]
- Choi, J.W.; Kim, S.; Kim, T.M.; Kim, Y.M.; Seo, H.W.; Park, T.S.; Jeong, J.W.; Song, G.; Han, J.Y. Basic fibroblast growth factor activates MEK/ERK cell signaling pathway and stimulates the proliferation of chicken primordial germ cells. PLoS ONE 2010, 5, e12968. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, H.; Zhang, Z.; Shi, Q.; Wang, D.; Zheng, M.; Li, B.; Song, J. Isolation of chicken embryonic stem cell and preparation of chicken chimeric model. Mol. Biol. Rep. 2013, 40, 2149–2156. [Google Scholar] [CrossRef]
- Aubel, P.; Pain, B. Chicken embryonic stem cells: Establishment and characterization. Methods Mol. Biol. 2013, 1074, 137–150. [Google Scholar]
- Kim, Y.M.; Park, Y.H.; Lim, J.M.; Jung, H.; Han, J.Y. Technical note: Induction of pluripotent stem cell-like cells from chicken feather follicle cells. J. Anim. Sci. 2017, 95, 3479–3486. [Google Scholar] [CrossRef]
- Fuet, A.; Montillet, G.; Jean, C.; Aubel, P.; Kress, C.; Rival-Gervier, S.; Pain, B. NANOG Is Required for the Long-Term Establishment of Avian Somatic Reprogrammed Cells. Stem Cell Rep. 2018, 11, 1272–1286. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Intarapat, S.; Sukparangsi, W.; Gusev, O.; Sheng, G. A Bird’s-Eye View of Endangered Species Conservation: Avian Genomics and Stem Cell Approaches for Green Peafowl (Pavo muticus). Genes 2023, 14, 2040. https://doi.org/10.3390/genes14112040
Intarapat S, Sukparangsi W, Gusev O, Sheng G. A Bird’s-Eye View of Endangered Species Conservation: Avian Genomics and Stem Cell Approaches for Green Peafowl (Pavo muticus). Genes. 2023; 14(11):2040. https://doi.org/10.3390/genes14112040
Chicago/Turabian StyleIntarapat, Sittipon, Woranop Sukparangsi, Oleg Gusev, and Guojun Sheng. 2023. "A Bird’s-Eye View of Endangered Species Conservation: Avian Genomics and Stem Cell Approaches for Green Peafowl (Pavo muticus)" Genes 14, no. 11: 2040. https://doi.org/10.3390/genes14112040
APA StyleIntarapat, S., Sukparangsi, W., Gusev, O., & Sheng, G. (2023). A Bird’s-Eye View of Endangered Species Conservation: Avian Genomics and Stem Cell Approaches for Green Peafowl (Pavo muticus). Genes, 14(11), 2040. https://doi.org/10.3390/genes14112040