
A DNA Methylation Perspective on Infertility

 , , , and
, , , and 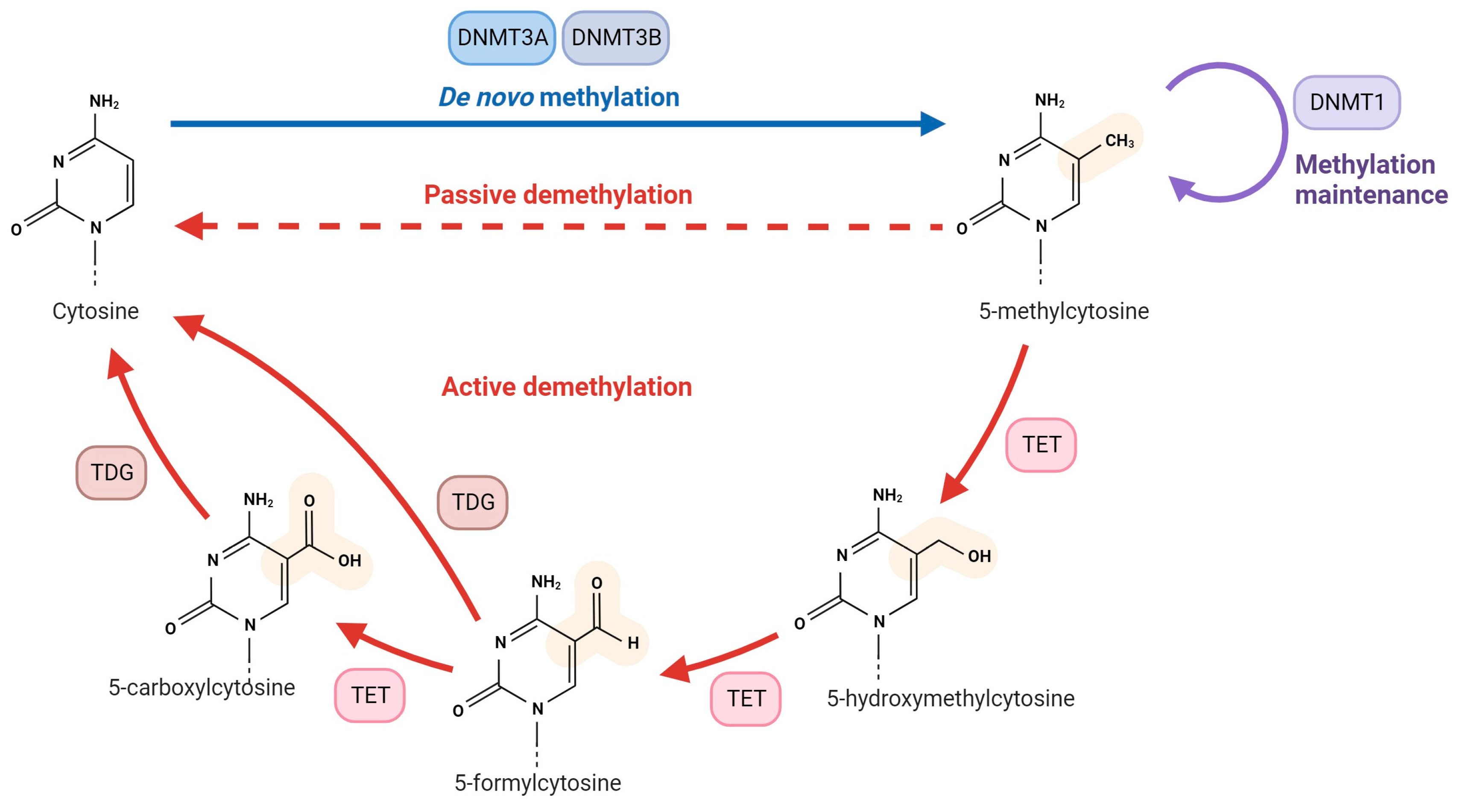{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Overview of DNA Methylation
3. DNA Methylation during Embryogenesis
4. DNA Methylation during Gametogenesis
5. DNA Methylation and Male Infertility
6. DNA Methylation and Female Infertility
6.1. Oogenesis
6.2. Oocyte Activation Deficiency
6.3. DNA Methylation and Fertility-Related Diseases
7. DNA Methylation and Age-Related Infertility
7.1. Male Infertility
7.2. Female Infertility
8. DNA Methylation Alterations in Assisted Reproductive Technology
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Agarwal, A.; Mulgund, A.; Hamada, A.; Chyatte, M.R. A unique view on male infertility around the globe. Reprod. Biol. Endocrinol. 2015, 13, 37. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Singh, A.K. Trends of male factor infertility, an important cause of infertility: A review of literature. J. Hum. Reprod. Sci. 2015, 8, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Cooper, T.G.; Jockenhövel, F.; Nieschlag, E. Variations in semen parameters from fathers. Hum. Reprod. 1991, 6, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Levine, H.; Jørgensen, N.; Martino-Andrade, A.; Mendiola, J.; Weksler-Derri, D.; Mindlis, I.; Pinotti, R.; Swan, S.H. Temporal trends in sperm count: A systematic review and meta-regression analysis. Hum. Reprod. Update 2017, 23, 646–659. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Gong, T.T.; Jiang, Y.T.; Zhang, S.; Zhao, Y.H.; Wu, Q.J. Global, regional, and national prevalence and disability-adjusted life-years for infertility in 195 countries and territories, 1990-2017: Results from a global burden of disease study, 2017. Aging 2019, 11, 10952–10991. [Google Scholar] [CrossRef]
- Walker, M.H.; Tobler, K.J. Female Infertility. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Rotondo, J.C.; Lanzillotti, C.; Mazziotta, C.; Tognon, M.; Martini, F. Epigenetics of Male Infertility: The Role of DNA Methylation. Front. Cell Dev. Biol. 2021, 9, 689624. [Google Scholar] [CrossRef]
- Vander Borght, M.; Wyns, C. Fertility and infertility: Definition and epidemiology. Clin. Biochem. 2018, 62, 2–10. [Google Scholar] [CrossRef]
- Jain, M.; Singh, M. Assisted reproductive technology (ART) techniques. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Gelbaya, T.A.; Potdar, N.; Jeve, Y.B.; Nardo, L.G. Definition and epidemiology of unexplained infertility. Obstet. Gynecol. Surv. 2014, 69, 109–115. [Google Scholar] [CrossRef]
- Allis, C.D.; Jenuwein, T. The molecular hallmarks of epigenetic control. Nat. Rev. Genet. 2016, 17, 487–500. [Google Scholar] [CrossRef]
- Ben Maamar, M.; Beck, D.; Nilsson, E.; McCarrey, J.R.; Skinner, M.K. Developmental alterations in DNA methylation during gametogenesis from primordial germ cells to sperm. iScience 2022, 25, 103786. [Google Scholar] [CrossRef]
- Stewart, K.R.; Veselovska, L.; Kelsey, G. Establishment and functions of DNA methylation in the germline. Epigenomics 2016, 8, 1399–1413. [Google Scholar] [CrossRef] [PubMed]
- Shafqat, A.; Kashir, J.; Alsalameh, S.; Alkattan, K.; Yaqinuddin, A. Fertilization, Oocyte Activation, Calcium Release and Epigenetic Remodelling: Lessons from Cancer Models. Front. Cell Dev. Biol. 2022, 10, 781953. [Google Scholar] [CrossRef] [PubMed]
- Giller, A.; Andrawus, M.; Gutman, D.; Atzmon, G. Pregnancy as a model for aging. Ageing Res. Rev. 2020, 62, 101093. [Google Scholar] [CrossRef] [PubMed]
- Salmeri, N.; Carbone, I.F.; Cavoretto, P.I.; Farina, A.; Morano, D. Epigenetics Beyond Fetal Growth Restriction: A Comprehensive Overview. Mol. Diagn. Ther. 2022, 26, 607–626. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, M.V.C.; Bourc’his, D. The diverse roles of DNA methylation in mammalian development and disease. Nat. Rev. Mol. Cell Biol. 2019, 20, 590–607. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.L.; Grant, P.A. The role of DNA methylation and histone modifications in transcriptional regulation in humans. Subcell. Biochem. 2013, 61, 289–317. [Google Scholar] [CrossRef] [PubMed]
- Moore, L.D.; Le, T.; Fan, G. DNA Methylation and Its Basic Function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef]
- Reik, W.; Dean, W.; Walter, J. Epigenetic reprogramming in mammalian development. Science 2001, 293, 1089–1093. [Google Scholar] [CrossRef]
- Marcho, C.; Oluwayiose, O.A.; Pilsner, J.R. The preconception environment and sperm epigenetics. Andrology 2020, 8, 924–942. [Google Scholar] [CrossRef]
- Santos, F.; Dean, W. Epigenetic reprogramming during early development in mammals. Reproduction 2004, 127, 643–651. [Google Scholar] [CrossRef]
- Gkountela, S.; Zhang, K.X.; Shafiq, T.A.; Liao, W.W.; Hargan-Calvopiña, J.; Chen, P.Y.; Clark, A.T. DNA Demethylation Dynamics in the Human Prenatal Germline. Cell 2015, 161, 1425–1436. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Yan, L.; Guo, H.; Li, L.; Hu, B.; Zhao, Y.; Yong, J.; Hu, Y.; Wang, X.; Wei, Y.; et al. The Transcriptome and DNA Methylome Landscapes of Human Primordial Germ Cells. Cell 2015, 161, 1437–1452. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Chen, T. DNA Methylation Reprogramming during Mammalian Development. Genes 2019, 10, 257. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.C.; Feng, S.Q. Epigenetic Modifications as Therapeutic Targets. Curr. Drug Targets 2020, 21, 1046. [Google Scholar] [CrossRef] [PubMed]
- Lucifero, D.; Mann, M.R.; Bartolomei, M.S.; Trasler, J.M. Gene-specific timing and epigenetic memory in oocyte imprinting. Hum. Mol. Genet. 2004, 13, 839–849. [Google Scholar] [CrossRef] [PubMed]
- Dupont, C.; Armant, D.R.; Brenner, C.A. Epigenetics: Definition, mechanisms and clinical perspective. Semin. Reprod. Med. 2009, 27, 351–357. [Google Scholar] [CrossRef]
- Henckel, A.; Chebli, K.; Kota, S.K.; Arnaud, P.; Feil, R. Transcription and histone methylation changes correlate with imprint acquisition in male germ cells. EMBO J. 2012, 31, 606–615. [Google Scholar] [CrossRef]
- Ravitsky, V.; Kimmins, S. The forgotten men: Rising rates of male infertility urgently require new approaches for its prevention, diagnosis and treatment. Biol. Reprod. 2019, 101, 872–874. [Google Scholar] [CrossRef]
- Rotondo, J.C.; Bosi, S.; Bazzan, E.; Di Domenico, M.; De Mattei, M.; Selvatici, R.; Patella, A.; Marci, R.; Tognon, M.; Martini, F. Methylenetetrahydrofolate reductase gene promoter hypermethylation in semen samples of infertile couples correlates with recurrent spontaneous abortion. Hum. Reprod. 2012, 27, 3632–3638. [Google Scholar] [CrossRef]
- Agarwal, A.; Allamaneni, S.S. Sperm DNA damage assessment: A test whose time has come. Fertil. Steril. 2005, 84, 850–853. [Google Scholar] [CrossRef]
- Bungum, M.; Bungum, L.; Giwercman, A. Sperm chromatin structure assay (SCSA): A tool in diagnosis and treatment of infertility. Asian J. Androl. 2011, 13, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Li, T.; Xie, Y.; Guo, Y.; Yang, Q.-Y.; Liang, X.; Deng, C.-H.; Liu, G.-H. Sperm DNA fragmentation index influences assisted reproductive technology outcome: A systematic review and meta-analysis combined with a retrospective cohort study. Andrologia 2019, 51, e13263. [Google Scholar] [CrossRef] [PubMed]
- Khezri, A.; Narud, B.; Stenseth, E.-B.; Johannisson, A.; Myromslien, F.D.; Gaustad, A.H.; Wilson, R.C.; Lyle, R.; Morrell, J.M.; Kommisrud, E.; et al. DNA methylation patterns vary in boar sperm cells with different levels of DNA fragmentation. BMC Genom. 2019, 20, 897. [Google Scholar] [CrossRef]
- Song, B.; Wang, C.; Chen, Y.; Li, G.; Gao, Y.; Zhu, F.; Wu, H.; Lv, M.; Zhou, P.; Wei, Z.; et al. Sperm DNA integrity status is associated with DNA methylation signatures of imprinted genes and non-imprinted genes. J. Assist. Reprod. Genet. 2021, 38, 2041–2048. [Google Scholar] [CrossRef] [PubMed]
- Haggarty, P. Genetic and metabolic determinants of human epigenetic variation. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Saraswathy, K.N.; Kaur, L.; Talwar, S.; Mishra, J.; Huidrom, S.; Sachdeva, M.P.; Puri, M. Methylenetetrahydrofolate Reductase Gene-specific Methylation and Recurrent Miscarriages: A Case-Control Study from North India. J. Hum. Reprod. Sci. 2018, 11, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Kulac, T.; Hekim, N.; Kocamanoglu, F.; Beyaz, C.; Gunes, S.; Asci, R. Methylation patterns of methylenetetrahydrofolate reductase gene promoter in infertile males. Andrologia 2021, 53, e13942. [Google Scholar] [CrossRef]
- Födinger, M.; Hörl, W.H.; Sunder-Plassmann, G. Molecular biology of 5,10-methylenetetrahydrofolate reductase. J. Nephrol. 2000, 13, 20–33. [Google Scholar]
- Chern, C.L.; Huang, R.F.; Chen, Y.H.; Cheng, J.T.; Liu, T.Z. Folate deficiency-induced oxidative stress and apoptosis are mediated via homocysteine-dependent overproduction of hydrogen peroxide and enhanced activation of NF-kappaB in human Hep G2 cells. Biomed. Pharmacother. 2001, 55, 434–442. [Google Scholar] [CrossRef]
- Choi, Y.; Kim, J.O.; Shim, S.H.; Lee, Y.; Kim, J.H.; Jeon, Y.J.; Ko, J.J.; Lee, W.S.; Kim, N.K. Genetic Variation of Methylenetetrahydrofolate Reductase (MTHFR) and Thymidylate Synthase (TS) Genes Is Associated with Idiopathic Recurrent Implantation Failure. PLoS ONE 2016, 11, e0160884. [Google Scholar] [CrossRef]
- Poorang, S.; Abdollahi, S.; Anvar, Z.; Tabei, S.M.B.; Jahromi, B.N.; Moein-Vaziri, N.; Gharesi-Fard, B.; Banaei, M.; Dastgheib, S.A. The Impact of Methylenetetrahydrofolate Reductase (MTHFR) Sperm Methylation and Variants on Semen Parameters and the Chance of Recurrent Pregnancy Loss in the Couple. Clin. Lab. 2018, 64, 1121–1128. [Google Scholar] [CrossRef] [PubMed]
- Ullah, N.; Mansoor, A.; Micheal, S.; Mirza, B.; Qamar, R.; Mazhar, K.; Siddiqi, S. MTHFR polymorphisms as risk for male infertility in Pakistan and its comparison with socioeconomic status in the world. Pers. Med. 2019, 16, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Shen, O.; Qin, Y.; Niu, X.; Lu, C.; Xia, Y.; Song, L.; Wang, S.; Wang, X. Idiopathic male infertility is strongly associated with aberrant promoter methylation of methylenetetrahydrofolate reductase (MTHFR). PLoS ONE 2010, 5, e13884. [Google Scholar] [CrossRef] [PubMed]
- Botezatu, A.; Socolov, R.; Socolov, D.; Iancu, I.V.; Anton, G. Methylation pattern of methylene tetrahydrofolate reductase and small nuclear ribonucleoprotein polypeptide N promoters in oligoasthenospermia: A case-control study. Reprod. Biomed. Online 2014, 28, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Karaca, M.Z.; Konac, E.; Yurteri, B.; Bozdag, G.; Sogutdelen, E.; Bilen, C.Y. Association between methylenetetrahydrofolate reductase (MTHFR) gene promoter hypermethylation and the risk of idiopathic male infertility. Andrologia 2017, 49, e12698. [Google Scholar] [CrossRef] [PubMed]
- Khazamipour, N.; Noruzinia, M.; Fatehmanesh, P.; Keyhanee, M.; Pujol, P. MTHFR promoter hypermethylation in testicular biopsies of patients with non-obstructive azoospermia: The role of epigenetics in male infertility. Hum. Reprod. 2009, 24, 2361–2364. [Google Scholar] [CrossRef] [PubMed]
- Uysal, F.; Akkoyunlu, G.; Ozturk, S. Decreased expression of DNA methyltransferases in the testes of patients with non-obstructive azoospermia leads to changes in global DNA methylation levels. Reprod. Fertil. Dev. 2019, 31, 1386–1394. [Google Scholar] [CrossRef]
- Tang, Q.; Chen, Y.; Wu, W.; Ding, H.; Xia, Y.; Chen, D.; Wang, X. Idiopathic male infertility and polymorphisms in the DNA methyltransferase genes involved in epigenetic marking. Sci. Rep. 2017, 7, 11219. [Google Scholar] [CrossRef]
- Zhu, W.; Du, J.; Chen, Q.; Zhang, Z.; Wu, B.; Xu, J.; Li, T.; Bi, Y.; Shi, H.; Li, R. Association of UHRF1 gene polymorphisms with oligospermia in Chinese males. J. Assist. Reprod. Genet. 2019, 36, 2563–2573. [Google Scholar] [CrossRef]
- Dong, J.; Wang, X.; Cao, C.; Wen, Y.; Sakashita, A.; Chen, S.; Zhang, J.; Zhang, Y.; Zhou, L.; Luo, M.; et al. UHRF1 suppresses retrotransposons and cooperates with PRMT5 and PIWI proteins in male germ cells. Nat. Commun. 2019, 10, 4705. [Google Scholar] [CrossRef]
- Ni, K.; Dansranjavin, T.; Rogenhofer, N.; Oeztuerk, N.; Deuker, J.; Bergmann, M.; Schuppe, H.C.; Wagenlehner, F.; Weidner, W.; Steger, K.; et al. TET enzymes are successively expressed during human spermatogenesis and their expression level is pivotal for male fertility. Hum. Reprod. 2016, 31, 1411–1424. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Liu, L.; Wang, H.; Gou, M.; Gong, P.; Tian, C.; Deng, W.; Yang, J.; Zhou, T.T.; Xu, G.L.; et al. Tet1 Deficiency Leads to Premature Reproductive Aging by Reducing Spermatogonia Stem Cells and Germ Cell Differentiation. iScience 2020, 23, 100908. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, S.; Bergmann, M.; Bohle, R.M.; Weidner, W.; Steger, K. Genetic imprinting during impaired spermatogenesis. Mol. Hum. Reprod. 2006, 12, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Li, X.P.; Hao, C.L.; Wang, Q.; Yi, X.M.; Jiang, Z.S. H19 gene methylation status is associated with male infertility. Exp. Ther. Med. 2016, 12, 451–456. [Google Scholar] [CrossRef]
- Cannarella, R.; Crafa, A.; Barbagallo, F.; Lundy, S.D.; La Vignera, S.; Condorelli, R.A.; Calogero, A.E. H19 Sperm Methylation in Male Infertility: A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2023, 24, 7224. [Google Scholar] [CrossRef]
- Leitão, E.; Di Persio, S.; Laurentino, S.; Wöste, M.; Dugas, M.; Kliesch, S.; Neuhaus, N.; Horsthemke, B. The sperm epigenome does not display recurrent epimutations in patients with severely impaired spermatogenesis. Clin. Epigenetics 2020, 12, 61. [Google Scholar] [CrossRef]
- Kläver, R.; Tüttelmann, F.; Bleiziffer, A.; Haaf, T.; Kliesch, S.; Gromoll, J. DNA methylation in spermatozoa as a prospective marker in andrology. Andrology 2013, 1, 731–740. [Google Scholar] [CrossRef]
- Montjean, D.; Ravel, C.; Benkhalifa, M.; Cohen-Bacrie, P.; Berthaut, I.; Bashamboo, A.; McElreavey, K. Methylation changes in mature sperm deoxyribonucleic acid from oligozoospermic men: Assessment of genetic variants and assisted reproductive technology outcome. Fertil. Steril. 2013, 100, 1241–1247. [Google Scholar] [CrossRef]
- El Hajj, N.; Zechner, U.; Schneider, E.; Tresch, A.; Gromoll, J.; Hahn, T.; Schorsch, M.; Haaf, T. Methylation status of imprinted genes and repetitive elements in sperm DNA from infertile males. Sex. Dev. 2011, 5, 60–69. [Google Scholar] [CrossRef]
- Dong, H.; Wang, Y.; Zou, Z.; Chen, L.; Shen, C.; Xu, S.; Zhang, J.; Zhao, F.; Ge, S.; Gao, Q.; et al. Abnormal Methylation of Imprinted Genes and Cigarette Smoking: Assessment of Their Association With the Risk of Male Infertility. Reprod. Sci. 2017, 24, 114–123. [Google Scholar] [CrossRef]
- Peng, Y.; Guo, R.; Shi, B.; Li, D. The role of long non-coding RNA H19 in infertility. Cell Death Discov. 2023, 9, 268. [Google Scholar] [CrossRef] [PubMed]
- Boissonnas, C.C.; Abdalaoui, H.E.; Haelewyn, V.; Fauque, P.; Dupont, J.M.; Gut, I.; Vaiman, D.; Jouannet, P.; Tost, J.; Jammes, H. Specific epigenetic alterations of IGF2-H19 locus in spermatozoa from infertile men. Eur. J. Hum. Genet. 2010, 18, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Santi, D.; De Vincentis, S.; Magnani, E.; Spaggiari, G. Impairment of sperm DNA methylation in male infertility: A meta-analytic study. Andrology 2017, 5, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Hattori, H.; Hiura, H.; Kitamura, A.; Miyauchi, N.; Kobayashi, N.; Takahashi, S.; Okae, H.; Kyono, K.; Kagami, M.; Ogata, T.; et al. Association of four imprinting disorders and ART. Clin. Epigenetics 2019, 11, 21. [Google Scholar] [CrossRef] [PubMed]
- Lou, H.; Le, F.; Hu, M.; Yang, X.; Li, L.; Wang, L.; Wang, N.; Gao, H.; Jin, F. Aberrant DNA Methylation of IGF2-H19 Locus in Human Fetus and in Spermatozoa from Assisted Reproductive Technologies. Reprod. Sci. 2019, 26, 997–1004. [Google Scholar] [CrossRef]
- Sendžikaitė, G.; Kelsey, G. The role and mechanisms of DNA methylation in the oocyte. Essays Biochem. 2019, 63, 691–705. [Google Scholar] [CrossRef]
- Gahurova, L.; Tomizawa, S.-i.; Smallwood, S.A.; Stewart-Morgan, K.R.; Saadeh, H.; Kim, J.; Andrews, S.R.; Chen, T.; Kelsey, G. Transcription and chromatin determinants of de novo DNA methylation timing in oocytes. Epigenetics Chromatin 2017, 10, 25. [Google Scholar] [CrossRef]
- Gu, C.; Liu, S.; Wu, Q.; Zhang, L.; Guo, F. Integrative single-cell analysis of transcriptome, DNA methylome and chromatin accessibility in mouse oocytes. Cell Res. 2019, 29, 110–123. [Google Scholar] [CrossRef]
- Veselovska, L.; Smallwood, S.A.; Saadeh, H.; Stewart, K.R.; Krueger, F.; Maupetit-Méhouas, S.; Arnaud, P.; Tomizawa, S.; Andrews, S.; Kelsey, G. Deep sequencing and de novo assembly of the mouse oocyte transcriptome define the contribution of transcription to the DNA methylation landscape. Genome Biol. 2015, 16, 209. [Google Scholar] [CrossRef]
- Hackett, J.A.; Sengupta, R.; Zylicz, J.J.; Murakami, K.; Lee, C.; Down, T.A.; Surani, M.A. Germline DNA demethylation dynamics and imprint erasure through 5-hydroxymethylcytosine. Science 2013, 339, 448–452. [Google Scholar] [CrossRef]
- Quenneville, S.; Verde, G.; Corsinotti, A.; Kapopoulou, A.; Jakobsson, J.; Offner, S.; Baglivo, I.; Pedone, P.V.; Grimaldi, G.; Riccio, A.; et al. In embryonic stem cells, ZFP57/KAP1 recognize a methylated hexanucleotide to affect chromatin and DNA methylation of imprinting control regions. Mol. Cell 2011, 44, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Kashir, J.; Ganesh, D.; Jones, C.; Coward, K. Oocyte activation deficiency and assisted oocyte activation: Mechanisms, obstacles and prospects for clinical application. Hum. Reprod. Open 2022, 2022, hoac003. [Google Scholar] [CrossRef] [PubMed]
- Kashir, J.; Heindryckx, B.; Jones, C.; De Sutter, P.; Parrington, J.; Coward, K. Oocyte activation, phospholipase C zeta and human infertility. Hum. Reprod. Update 2010, 16, 690–703. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, W.M.; Kashir, J.; Jones, C.; Coward, K. Oocyte activation and phospholipase C zeta (PLCζ): Diagnostic and therapeutic implications for assisted reproductive technology. Cell Commun. Signal. 2012, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Kashir, J.; Jones, C.; Coward, K. Calcium oscillations, oocyte activation, and phospholipase C zeta. In Calcium Signal; Springer: Dordrecht, The Netherlands, 2012; pp. 1095–1121. [Google Scholar]
- Kashir, J.; Nomikos, M.; Lai, F.A.; Swann, K. Sperm-induced Ca2+ release during egg activation in mammals. Biochem. Biophys. Res. Commun. 2014, 450, 1204–1211. [Google Scholar] [CrossRef] [PubMed]
- Sanusi, R. Calcium Signals and the Activation of Embryo Development in Mice and Humans. Ph.D. Thesis, Cardiff University, Cardiff, UK, 2016. [Google Scholar]
- Bonte, D. The Importance of Calcium: The Quest for Optimal Diagnosis and Treatment for Fertilization Failure and Embryo Arrest in Human. Ph.D. Thesis, Ghent University, Ghent, Belgium, 2019. [Google Scholar]
- Ozil, J.-P.; Banrezes, B.; Tóth, S.; Pan, H.; Schultz, R.M. Ca2+ oscillatory pattern in fertilized mouse eggs affects gene expression and development to term. Dev. Biol. 2006, 300, 534–544. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Wang, J.; Qu, R.; Zhang, W.; Tan, Y.; Sha, Y.; Li, L.; Yin, T. Genetic mechanisms of fertilization failure and early embryonic arrest: A comprehensive review. Hum. Reprod. Update 2023, dmad026. [Google Scholar] [CrossRef]
- Rawat, K.; Sandhu, A.; Gautam, V.; Saha, P.K.; Saha, L. Role of genomic DNA methylation in PCOS pathogenesis: A systematic review and meta-analysis involving case-controlled clinical studies. Mol. Hum. Reprod. 2022, 28, gaac024. [Google Scholar] [CrossRef]
- Xue, Q.; Lin, Z.; Cheng, Y.H.; Huang, C.C.; Marsh, E.; Yin, P.; Milad, M.P.; Confino, E.; Reierstad, S.; Innes, J.; et al. Promoter methylation regulates estrogen receptor 2 in human endometrium and endometriosis. Biol. Reprod. 2007, 77, 681–687. [Google Scholar] [CrossRef]
- Izawa, M.; Harada, T.; Taniguchi, F.; Ohama, Y.; Takenaka, Y.; Terakawa, N. An epigenetic disorder may cause aberrant expression of aromatase gene in endometriotic stromal cells. Fertil. Steril. 2008, 89, 1390–1396. [Google Scholar] [CrossRef]
- Xue, Q.; Lin, Z.; Yin, P.; Milad, M.P.; Cheng, Y.H.; Confino, E.; Reierstad, S.; Bulun, S.E. Transcriptional activation of steroidogenic factor-1 by hypomethylation of the 5’ CpG island in endometriosis. J. Clin. Endocrinol. Metab. 2007, 92, 3261–3267. [Google Scholar] [CrossRef] [PubMed]
- Zidan, H.E.; Rezk, N.A.; Alnemr, A.A.; Abd El Ghany, A.M. COX-2 gene promoter DNA methylation status in eutopic and ectopic endometrium of Egyptian women with endometriosis. J. Reprod. Immunol. 2015, 112, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Strawn, E.; Basir, Z.; Halverson, G.; Guo, S.W. Aberrant expression of deoxyribonucleic acid methyltransferases DNMT1, DNMT3A, and DNMT3B in women with endometriosis. Fertil. Steril. 2007, 87, 24–32. [Google Scholar] [CrossRef]
- Wu, Y.; Halverson, G.; Basir, Z.; Strawn, E.; Yan, P.; Guo, S.W. Aberrant methylation at HOXA10 may be responsible for its aberrant expression in the endometrium of patients with endometriosis. Am. J. Obstet. Gynecol. 2005, 193, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Strawn, E.; Basir, Z.; Halverson, G.; Guo, S.W. Promoter hypermethylation of progesterone receptor isoform B (PR-B) in endometriosis. Epigenetics 2006, 1, 106–111. [Google Scholar] [CrossRef]
- Mortlock, S.; Houshdaran, S.; Kosti, I.; Rahmioglu, N.; Nezhat, C.; Vitonis, A.F.; Andrews, S.V.; Grosjean, P.; Paranjpe, M.; Horne, A.W.; et al. Global endometrial DNA methylation analysis reveals insights into mQTL regulation and associated endometriosis disease risk and endometrial function. Commun. Biol. 2023, 6, 780. [Google Scholar] [CrossRef]
- Si, J.; Meir, A.Y.; Hong, X.; Wang, G.; Huang, W.; Pearson, C.; Adams, W.G.; Wang, X.; Liang, L. Maternal pre-pregnancy BMI, offspring epigenome-wide DNA methylation, and childhood obesity: Findings from the Boston Birth Cohort. BMC Med. 2023, 21, 317. [Google Scholar] [CrossRef]
- Sukur, G.; Uysal, F.; Cinar, O. High-fat diet induced obesity alters Dnmt1 and Dnmt3a levels and global DNA methylation in mouse ovary and testis. Histochem. Cell Biol. 2023, 159, 339–352. [Google Scholar] [CrossRef]
- Khandwala, Y.S.; Zhang, C.A.; Lu, Y.; Eisenberg, M.L. The age of fathers in the USA is rising: An analysis of 168 867 480 births from 1972 to 2015. Hum. Reprod. 2017, 32, 2110–2116. [Google Scholar] [CrossRef]
- Mathews, T.J.; Hamilton, B.E. Mean Age of Mothers is on the Rise: United States, 2000–2014. NCHS Data Brief. 2016, 232, 1–8. [Google Scholar]
- Gourinat, A.; Mazeaud, C.; Hubert, J.; Eschwege, P.; Koscinski, I. Impact of paternal age on assisted reproductive technology outcomes and offspring health: A systematic review. Andrology 2023, 11, 973–986. [Google Scholar] [CrossRef] [PubMed]
- Vitagliano, A.; Paffoni, A.; Viganò, P. Does maternal age affect assisted reproduction technology success rates after euploid embryo transfer? A systematic review and meta-analysis. Fertil. Steril. 2023, 120, 251–265. [Google Scholar] [CrossRef] [PubMed]
- Murugesu, S.; Kasaven, L.S.; Petrie, A.; Vaseekaran, A.; Jones, B.P.; Bracewell-Milnes, T.; Barcroft, J.F.; Grewal, K.J.; Getreu, N.; Galazis, N.; et al. Does advanced paternal age affect outcomes following assisted reproductive technology? A systematic review and meta-analysis. Reprod. BioMed. Online 2022, 45, 283–331. [Google Scholar] [CrossRef] [PubMed]
- McPherson, N.O.; Zander-Fox, D.; Vincent, A.D.; Lane, M. Combined advanced parental age has an additive negative effect on live birth rates-data from 4057 first IVF/ICSI cycles. J. Assist. Reprod. Genet. 2018, 35, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Agarwal, A.; Rohra, V.K.; Assidi, M.; Abu-Elmagd, M.; Turki, R.F. Effects of increased paternal age on sperm quality, reproductive outcome and associated epigenetic risks to offspring. Reprod. Biol. Endocrinol. 2015, 13, 35. [Google Scholar] [CrossRef]
- Sartorius, G.A.; Nieschlag, E. Paternal age and reproduction. Hum. Reprod. Update 2009, 16, 65–79. [Google Scholar] [CrossRef]
- Horvath, S.; Raj, K. DNA methylation-based biomarkers and the epigenetic clock theory of ageing. Nat. Rev. Genet. 2018, 19, 371–384. [Google Scholar] [CrossRef]
- Horvath, S. DNA methylation age of human tissues and cell types. Genome Biol. 2013, 14, R115. [Google Scholar] [CrossRef]
- Hannum, G.; Guinney, J.; Zhao, L.; Zhang, L.; Hughes, G.; Sadda, S.; Klotzle, B.; Bibikova, M.; Fan, J.-B.; Gao, Y.; et al. Genome-wide Methylation Profiles Reveal Quantitative Views of Human Aging Rates. Mol. Cell 2013, 49, 359–367. [Google Scholar] [CrossRef]
- Lu, A.T.; Quach, A.; Wilson, J.G.; Reiner, A.P.; Aviv, A.; Raj, K.; Hou, L.; Baccarelli, A.A.; Li, Y.; Stewart, J.D.; et al. DNA methylation GrimAge strongly predicts lifespan and healthspan. Aging 2019, 11, 303–327. [Google Scholar] [CrossRef]
- Levine, M.E.; Lu, A.T.; Quach, A.; Chen, B.H.; Assimes, T.L.; Bandinelli, S.; Hou, L.; Baccarelli, A.A.; Stewart, J.D.; Li, Y.; et al. An epigenetic biomarker of aging for lifespan and healthspan. Aging 2018, 10, 573–591. [Google Scholar] [CrossRef] [PubMed]
- Belsky, D.W.; Caspi, A.; Arseneault, L.; Baccarelli, A.; Corcoran, D.L.; Gao, X.; Hannon, E.; Harrington, H.L.; Rasmussen, L.J.H.; Houts, R.; et al. Quantification of the pace of biological aging in humans through a blood test, the DunedinPoAm DNA methylation algorithm. eLife 2020, 9, e54870. [Google Scholar] [CrossRef] [PubMed]
- Belsky, D.W.; Caspi, A.; Corcoran, D.L.; Sugden, K.; Poulton, R.; Arseneault, L.; Baccarelli, A.; Chamarti, K.; Gao, X.; Hannon, E.; et al. DunedinPACE, a DNA methylation biomarker of the pace of aging. eLife 2022, 11, e73420. [Google Scholar] [CrossRef] [PubMed]
- Shireby, G.L.; Davies, J.P.; Francis, P.T.; Burrage, J.; Walker, E.M.; Neilson, G.W.A.; Dahir, A.; Thomas, A.J.; Love, S.; Smith, R.G.; et al. Recalibrating the epigenetic clock: Implications for assessing biological age in the human cortex. Brain 2020, 143, 3763–3775. [Google Scholar] [CrossRef] [PubMed]
- Mongelli, A.; Panunzi, S.; Nesta, M.; Gottardi Zamperla, M.; Atlante, S.; Barbi, V.; Mongiardini, V.; Ferraro, F.; De Martino, S.; Cis, L.; et al. Distinguishable DNA methylation defines a cardiac-specific epigenetic clock. Clin. Epigenetics 2023, 15, 53. [Google Scholar] [CrossRef]
- Oluwayiose, O.A.; Wu, H.; Saddiki, H.; Whitcomb, B.W.; Balzer, L.B.; Brandon, N.; Suvorov, A.; Tayyab, R.; Sites, C.K.; Hill, L.; et al. Sperm DNA methylation mediates the association of male age on reproductive outcomes among couples undergoing infertility treatment. Sci. Rep. 2021, 11, 3216. [Google Scholar] [CrossRef]
- Pilsner, J.R.; Saddiki, H.; Whitcomb, B.W.; Suvorov, A.; Buck Louis, G.M.; Mumford, S.L.; Schisterman, E.F.; Oluwayiose, O.A.; Balzer, L.B. Sperm epigenetic clock associates with pregnancy outcomes in the general population. Hum. Reprod. 2022, 37, 1581–1593. [Google Scholar] [CrossRef]
- Yu, B.; Russanova, V.R.; Gravina, S.; Hartley, S.; Mullikin, J.C.; Ignezweski, A.; Graham, J.; Segars, J.H.; DeCherney, A.H.; Howard, B.H. DNA methylome and transcriptome sequencing in human ovarian granulosa cells links age-related changes in gene expression to gene body methylation and 3’-end GC density. Oncotarget 2015, 6, 3627–3643. [Google Scholar] [CrossRef]
- Marshall, K.L.; Wang, J.; Ji, T.; Rivera, R.M. The effects of biological aging on global DNA methylation, histone modification, and epigenetic modifiers in the mouse germinal vesicle stage oocyte. Anim. Reprod. 2018, 15, 1253–1267. [Google Scholar] [CrossRef]
- Li, Q.; Cai, M.; Wang, J.; Gao, Q.; Guo, X.; Jia, X.; Xu, S.; Zhu, H. Decreased ovarian function and autophagy gene methylation in aging rats. J. Ovarian Res. 2020, 13, 12. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: An expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, W.; Wu, Z.; Dai, L.; Koji, T. Changes in DNA Methylation of Oocytes and Granulosa Cells Assessed by HELMET during Folliculogenesis in Mouse Ovary. Acta Histochem. Cytochem. 2018, 51, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Ranke, M.B.; Saenger, P. Turner’s syndrome. Lancet 2001, 358, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Trolle, C.; Nielsen, M.M.; Skakkebæk, A.; Lamy, P.; Vang, S.; Hedegaard, J.; Nordentoft, I.; Ørntoft, T.F.; Pedersen, J.S.; Gravholt, C.H. Widespread DNA hypomethylation and differential gene expression in Turner syndrome. Sci. Rep. 2016, 6, 34220. [Google Scholar] [CrossRef] [PubMed]
- Rajpathak, S.N.; Deobagkar, D.D. Evidence for epigenetic alterations in Turner syndrome opens up feasibility of new pharmaceutical interventions. Curr. Pharm. Des. 2014, 20, 1778–1785. [Google Scholar] [CrossRef]
- Simpson, J.L. Genetic and phenotypic heterogeneity in ovarian failure: Overview of selected candidate genes. Ann. N. Y. Acad. Sci. 2008, 1135, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Noma, T.; Kanai, Y.; Kanai-Azuma, M.; Ishii, M.; Fujisawa, M.; Kurohmaru, M.; Kawakami, H.; Wood, S.A.; Hayashi, Y. Stage- and sex-dependent expressions of Usp9x, an X-linked mouse ortholog of Drosophila Fat facets, during gonadal development and oogenesis in mice. Gene Expr. Patterns 2002, 2, 87–91. [Google Scholar] [CrossRef]
- Luoh, S.W.; Bain, P.A.; Polakiewicz, R.D.; Goodheart, M.L.; Gardner, H.; Jaenisch, R.; Page, D.C. Zfx mutation results in small animal size and reduced germ cell number in male and female mice. Development 1997, 124, 2275–2284. [Google Scholar] [CrossRef]
- Rajpathak, S.N.; Vellarikkal, S.K.; Patowary, A.; Scaria, V.; Sivasubbu, S.; Deobagkar, D.D. Human 45, X fibroblast transcriptome reveals distinct differentially expressed genes including long noncoding RNAs potentially associated with the pathophysiology of Turner syndrome. PLoS ONE 2014, 9, e100076. [Google Scholar] [CrossRef]
- Moore, R.K.; Shimasaki, S. Molecular biology and physiological role of the oocyte factor, BMP-15. Mol. Cell. Endocrinol. 2005, 234, 67–73. [Google Scholar] [CrossRef]
- Paulini, F.; Melo, E.O. The role of oocyte-secreted factors GDF9 and BMP15 in follicular development and oogenesis. Reprod. Domest. Anim. 2011, 46, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, F.; McTavish, K.J.; Shimasaki, S. Integral role of GDF-9 and BMP-15 in ovarian function. Mol. Reprod. Dev. 2011, 78, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Afkhami, F.; Shahbazi, S.; Farzadi, L.; Danaei, S. Novel bone morphogenetic protein 15 (BMP15) gene variants implicated in premature ovarian insufficiency. Reprod. Biol. Endocrinol. 2022, 20, 42. [Google Scholar] [CrossRef] [PubMed]
- Di Pasquale, E.; Rossetti, R.; Marozzi, A.; Bodega, B.; Borgato, S.; Cavallo, L.; Einaudi, S.; Radetti, G.; Russo, G.; Sacco, M. Identification of new variants of human BMP15 gene in a large cohort of women with premature ovarian failure. J. Clin. Endocrinol. Metab. 2006, 91, 1976–1979. [Google Scholar] [CrossRef] [PubMed]
- Tiotiu, D.; Alvaro Mercadal, B.; Imbert, R.; Verbist, J.; Demeestere, I.; De Leener, A.; Englert, Y.; Vassart, G.; Costagliola, S.; Delbaere, A. Variants of the BMP15 gene in a cohort of patients with premature ovarian failure. Hum. Reprod. 2010, 25, 1581–1587. [Google Scholar] [CrossRef] [PubMed]
- Hanson, B.M.; Tao, X.; Zhan, Y.; Jenkins, T.G.; Morin, S.J.; Scott, R.T.; Seli, E.U. Young women with poor ovarian response exhibit epigenetic age acceleration based on evaluation of white blood cells using a DNA methylation-derived age prediction model. Hum. Reprod. 2020, 35, 2579–2588. [Google Scholar] [CrossRef]
- Lee, Y.; Bohlin, J.; Page, C.M.; Nustad, H.E.; Harris, J.R.; Magnus, P.; Jugessur, A.; Magnus, M.C.; Håberg, S.E.; Hanevik, H.I. Associations between epigenetic age acceleration and infertility. Hum. Reprod. 2022, 37, 2063–2074. [Google Scholar] [CrossRef]
- Suvakov, S.; Ghamrawi, R.; Cubro, H.; Tu, H.; White, W.M.; Tobah, Y.S.B.; Milic, N.M.; Grande, J.P.; Cunningham, J.M.; Chebib, F.T.; et al. Epigenetic and senescence markers indicate an accelerated ageing-like state in women with preeclamptic pregnancies. EBioMedicine 2021, 70, 103536. [Google Scholar] [CrossRef]
- Owen, C.M.; Segars, J.H., Jr. Imprinting disorders and assisted reproductive technology. Semin. Reprod. Med. 2009, 27, 417–428. [Google Scholar] [CrossRef]
- Gomes, M.V.; Huber, J.; Ferriani, R.A.; Amaral Neto, A.M.; Ramos, E.S. Abnormal methylation at the KvDMR1 imprinting control region in clinically normal children conceived by assisted reproductive technologies. Mol. Hum. Reprod. 2009, 15, 471–477. [Google Scholar] [CrossRef]
- Barberet, J.; Barry, F.; Choux, C.; Guilleman, M.; Karoui, S.; Simonot, R.; Bruno, C.; Fauque, P. What impact does oocyte vitrification have on epigenetics and gene expression? Clin. Epigenetics 2020, 12, 121. [Google Scholar] [CrossRef] [PubMed]
- Maher, E.R.; Afnan, M.; Barratt, C.L. Epigenetic risks related to assisted reproductive technologies: Epigenetics, imprinting, ART and icebergs? Hum. Reprod. 2003, 18, 2508–2511. [Google Scholar] [CrossRef]
- Novakovic, B.; Lewis, S.; Halliday, J.; Kennedy, J.; Burgner, D.P.; Czajko, A.; Kim, B.; Sexton-Oates, A.; Juonala, M.; Hammarberg, K.; et al. Assisted reproductive technologies are associated with limited epigenetic variation at birth that largely resolves by adulthood. Nat. Commun. 2019, 10, 3922. [Google Scholar] [CrossRef] [PubMed]
- Lidegaard, Ø.; Pinborg, A.; Andersen, A.N. Imprinting diseases and IVF: Danish National IVF cohort study. Hum. Reprod. 2005, 20, 950–954. [Google Scholar] [CrossRef] [PubMed]
- Doornbos, M.E.; Maas, S.M.; McDonnell, J.; Vermeiden, J.P.W.; Hennekam, R.C.M. Infertility, assisted reproduction technologies and imprinting disturbances: A Dutch study. Hum. Reprod. 2007, 22, 2476–2480. [Google Scholar] [CrossRef]
- Sutcliffe, A.G.; Peters, C.J.; Bowdin, S.; Temple, K.; Reardon, W.; Wilson, L.; Clayton-Smith, J.; Brueton, L.A.; Bannister, W.; Maher, E.R. Assisted reproductive therapies and imprinting disorders--a preliminary British survey. Hum. Reprod. 2006, 21, 1009–1011. [Google Scholar] [CrossRef]
- El Hajj, N.; Haertle, L.; Dittrich, M.; Denk, S.; Lehnen, H.; Hahn, T.; Schorsch, M.; Haaf, T. DNA methylation signatures in cord blood of ICSI children. Hum. Reprod. 2017, 32, 1761–1769. [Google Scholar] [CrossRef]
- Smallwood, S.A.; Kelsey, G. De novo DNA methylation: A germ cell perspective. Trends Genet. 2012, 28, 33–42. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shacfe, G.; Turko, R.; Syed, H.H.; Masoud, I.; Tahmaz, Y.; Samhan, L.M.; Alkattan, K.; Shafqat, A.; Yaqinuddin, A. A DNA Methylation Perspective on Infertility. Genes 2023, 14, 2132. https://doi.org/10.3390/genes14122132
Shacfe G, Turko R, Syed HH, Masoud I, Tahmaz Y, Samhan LM, Alkattan K, Shafqat A, Yaqinuddin A. A DNA Methylation Perspective on Infertility. Genes. 2023; 14(12):2132. https://doi.org/10.3390/genes14122132
Chicago/Turabian StyleShacfe, Ghaleb, Rasoul Turko, Haadi Hammad Syed, Ibraheem Masoud, Yahya Tahmaz, Lara M. Samhan, Khaled Alkattan, Areez Shafqat, and Ahmed Yaqinuddin. 2023. "A DNA Methylation Perspective on Infertility" Genes 14, no. 12: 2132. https://doi.org/10.3390/genes14122132
APA StyleShacfe, G., Turko, R., Syed, H. H., Masoud, I., Tahmaz, Y., Samhan, L. M., Alkattan, K., Shafqat, A., & Yaqinuddin, A. (2023). A DNA Methylation Perspective on Infertility. Genes, 14(12), 2132. https://doi.org/10.3390/genes14122132